
Delivery of pDNA to the Lung by Lipopolyplexes Using N-Lauroylsarcosine and Effect on the Pulmonary Fibrosis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Construction of pDNA
2.3. Preparation of the Lipopolyplexes
2.4. Animals
2.5. Physicochemical Characteristics of Lipopolyplexes
2.6. In Vivo Gene Expression Experiments
2.7. Luciferase Assay
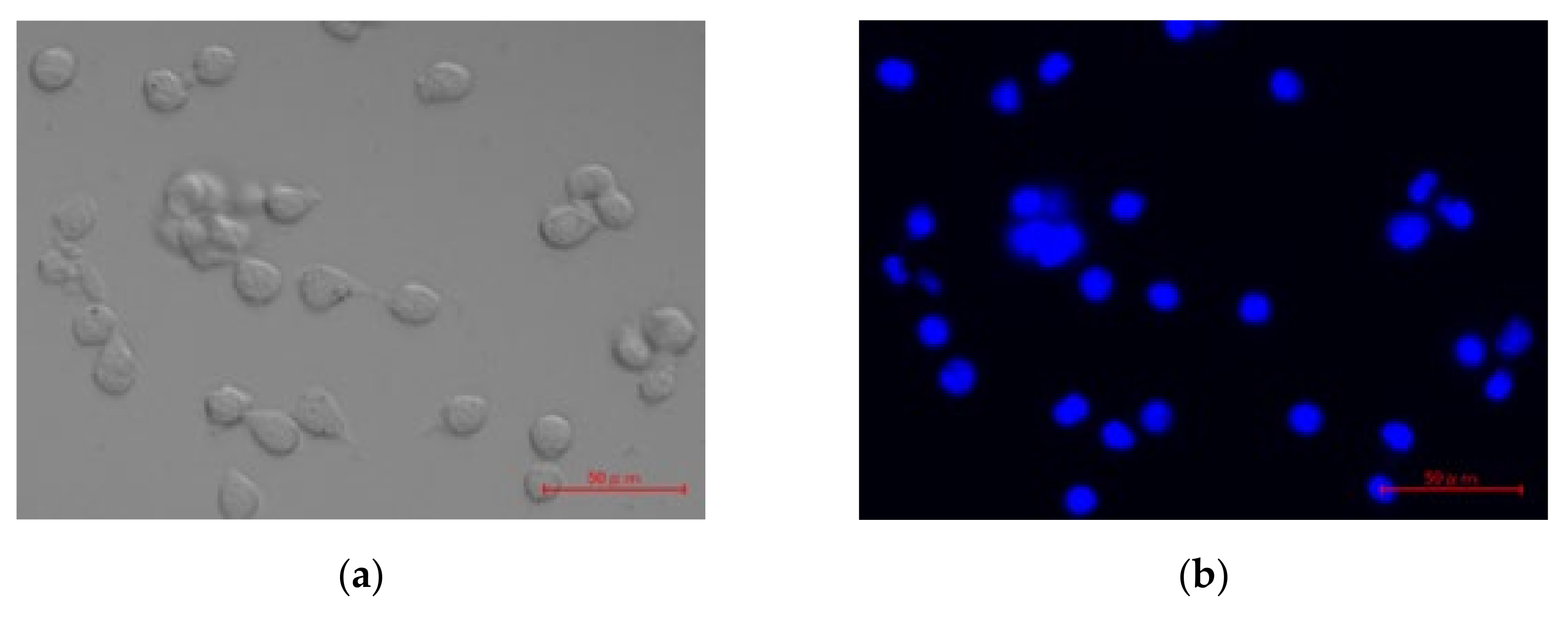
2.8. In Vitro Cellular Uptake of pDNA/DGL/DOTMA/LS
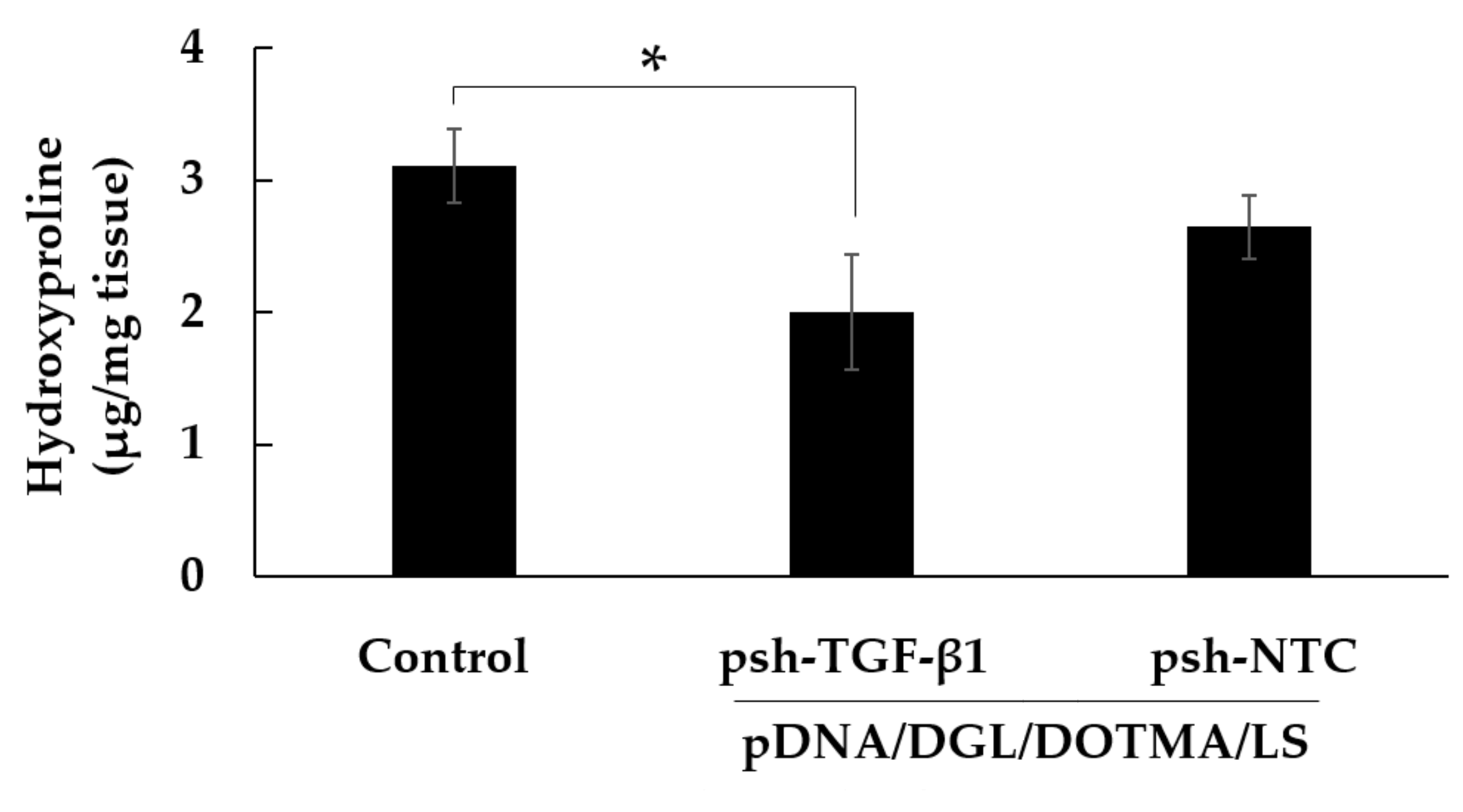
2.9. In Vivo Hydroxyproline Quantification
2.10. Statistical Analysis
3. Results
3.1. Effect of Cationic Polymers on Particle Size and ζ-Potential of Lipopolyplexes
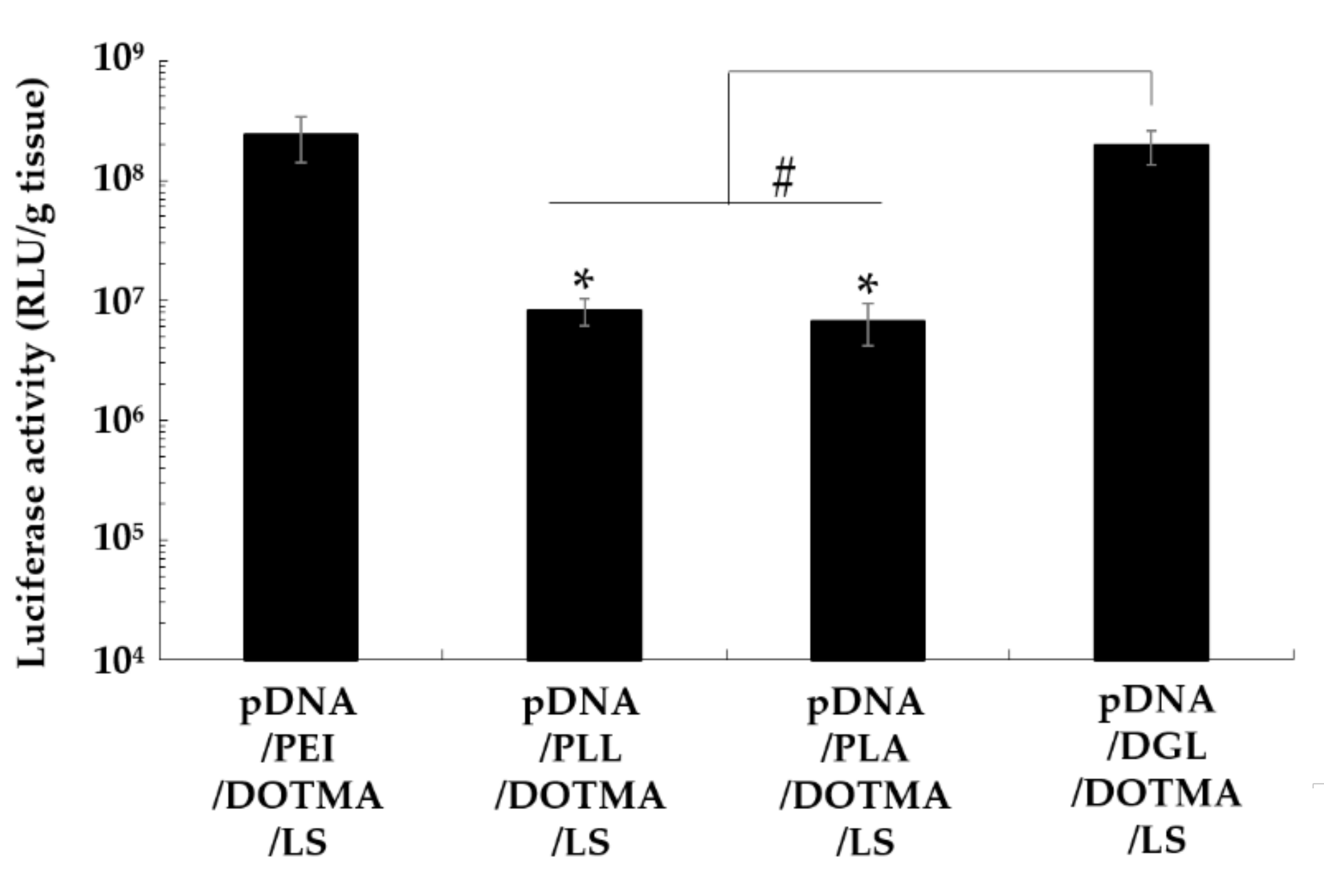
3.2. Effect of Cationic Polymers on the Lung Transfection Efficiency of Lipopolyplexes
3.3. Effect of Cationic Lipids on Particle Size and ζ-Potential of Lipopolyplexes
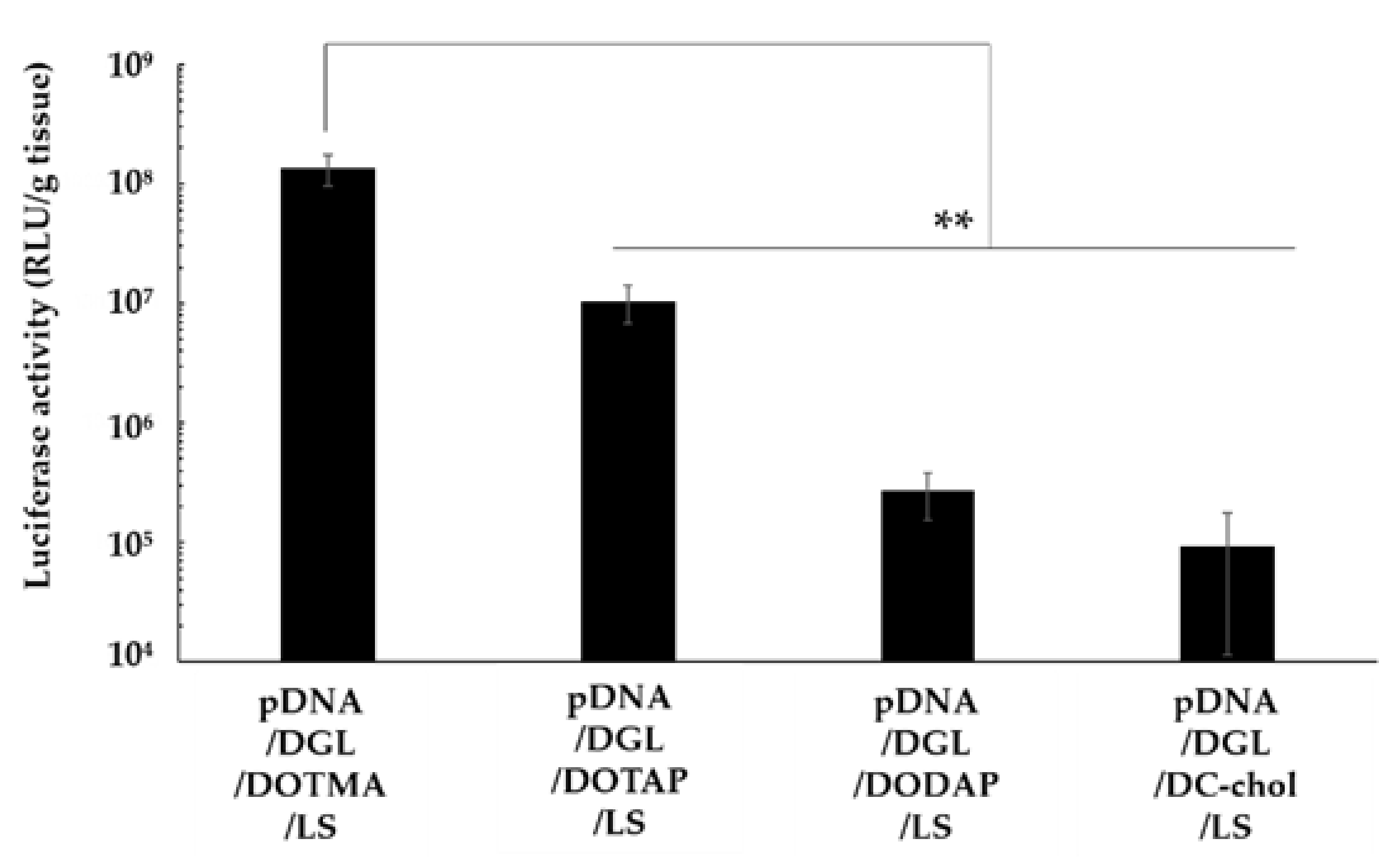
3.4. Effect of Cationic Lipids on the Lung Transfection Efficiency of Lipopolyplexes
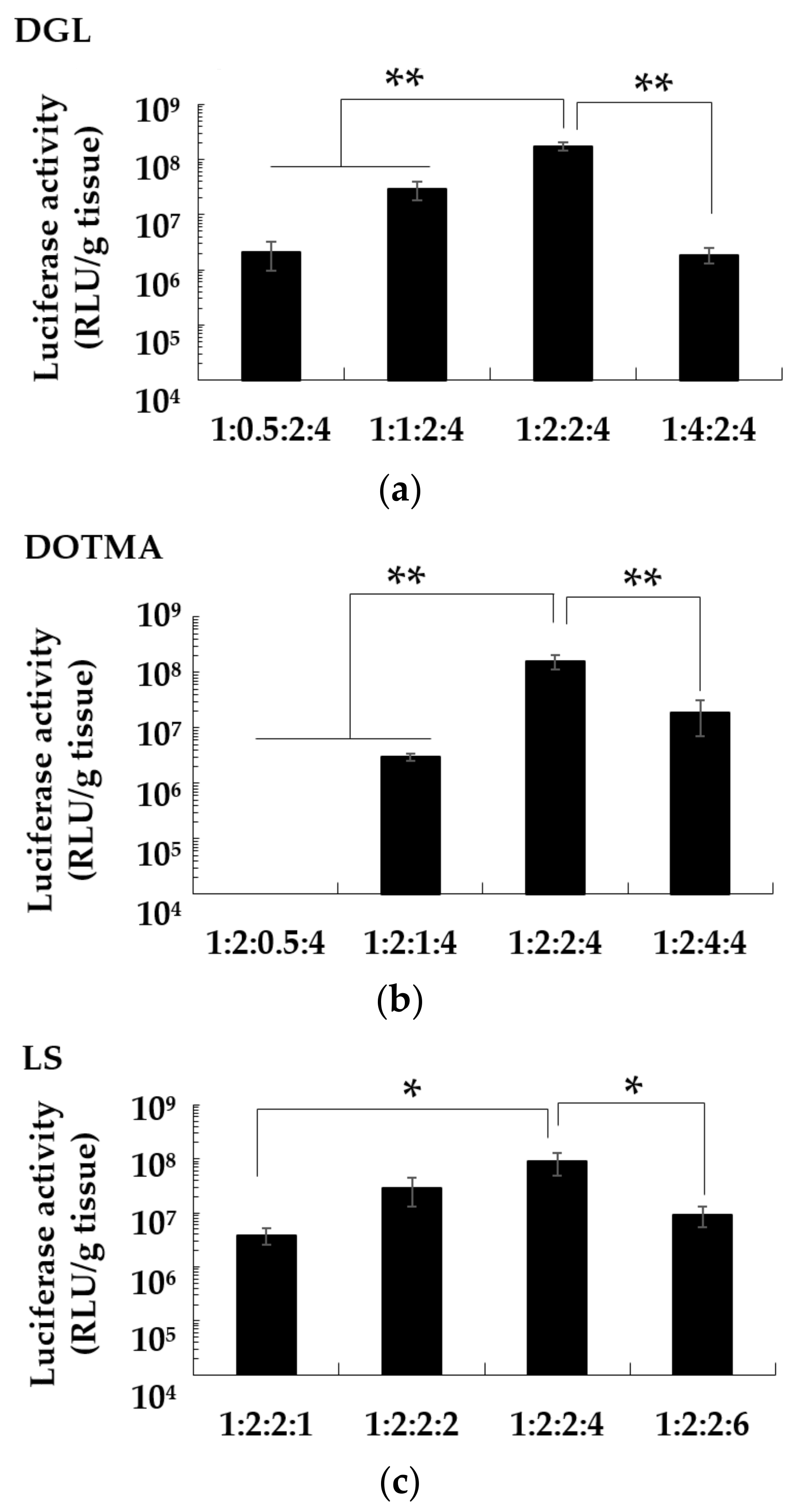
3.5. Effect of Charge Ratio on the Gene Expression of pDNA/DGL/DOTMA/LS in the Lung
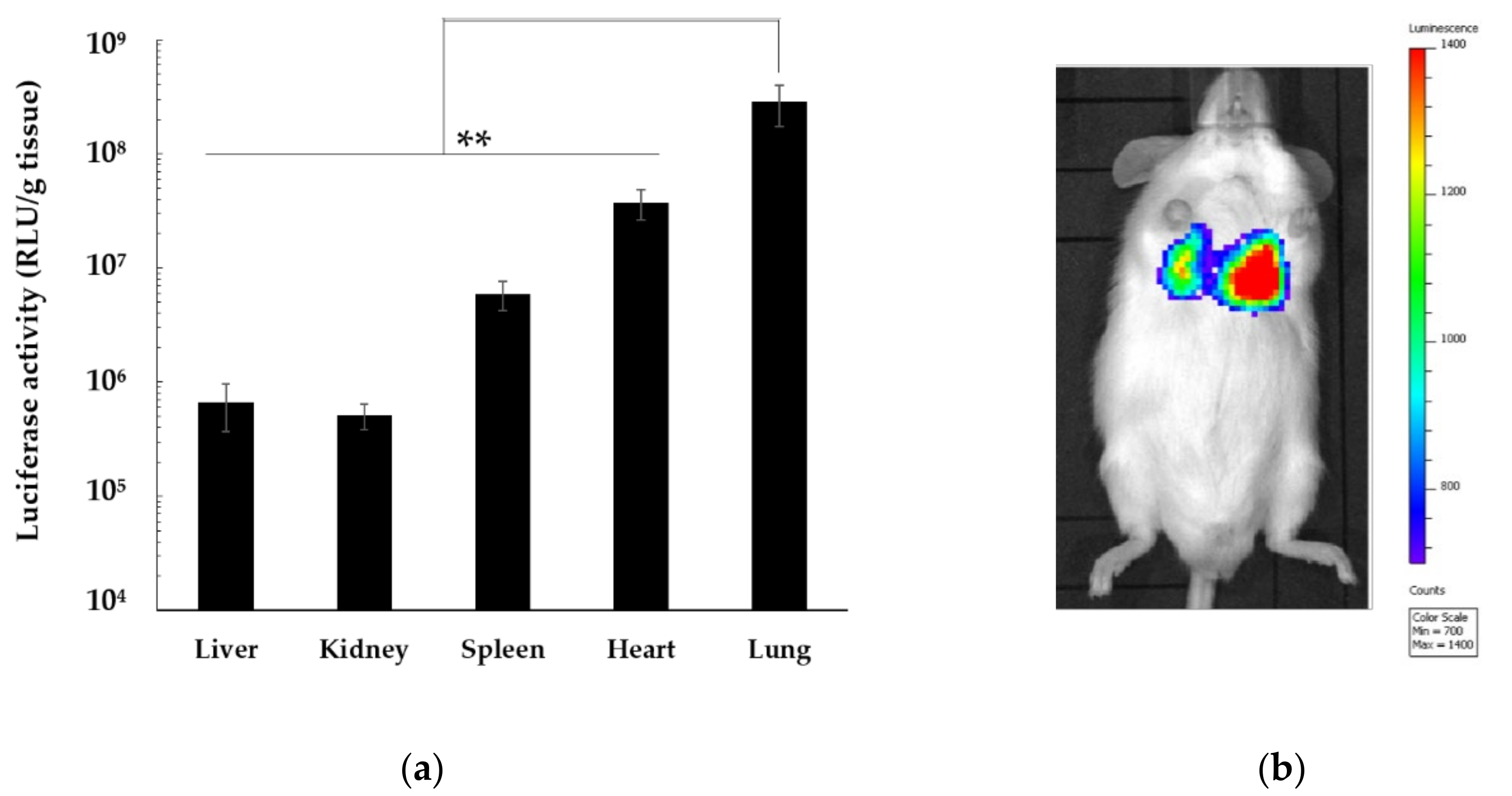
3.6. In Vivo Distribution of Gene Expression by pDNA/DGL/DOTMA/LS
3.7. Intracellular Uptake Pathway of pDNA/DGL/DOTMA/LS
3.8. Effect of pDNA/DGL/DOTMA/LS Containing psh-TGF-β1 on the Mice with IPF
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Skibba, M.; Drelich, A.; Poellmann, M.; Hong, S.; Brasier, A.R. Nanoapproaches to modifying epigenetics of epithelial mesenchymal transition for treatment of pulmonary fibrosis. Front. Pharmacol. 2020, 11, 607689. [Google Scholar] [CrossRef] [PubMed]
- Factor, P. Gene therapy for asthma. Mol. Ther. 2003, 7, 148–152. [Google Scholar] [CrossRef]
- Keedy, V.; Wang, W.; Schiller, J.; Chada, S.; Slovis, B.; Coffee, K.; Worrell, J.; Thet, L.A.; Johnson, D.H.; Carbone, D.P. Phase I study of adenovirus p53 administered by bronchoalveolar lavage in patients with bronchioloalveolar cell lung carcinoma: ECOG 6597. J. Clin. Oncol. 2008, 26, 4166–4171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itani, R.; Al Faraj, A. siRNA Conjugated nanoparticles-a next generation strategy to treat lung cancer. Int. J. Mol. Sci. 2019, 20, 6088. [Google Scholar] [CrossRef] [Green Version]
- Raghu, G.; Collard, H.R.; Egan, J.J.; Martinez, F.J.; Behr, J.; Brown, K.K.; Colby, T.V.; Cordier, J.F.; Flaherty, K.R.; Lasky, J.A.; et al. An official ATS/ERS/JRS/ALAT statement: Idiopathic pulmonary fibrosis: Evidence-based guidelines for diagnosis and management. Am. J. Respir. Crit. Care Med. 2011, 183, 788–824. [Google Scholar] [CrossRef]
- Lancaster, L.H.; de Andrade, J.A.; Zibrak, J.D.; Padilla, M.L.; Albera, C.; Nathan, S.D.; Wijsenbeek, M.S.; Stauffer, J.L.; Kirchgaessler, K.U.; Costabel, U. Pirfenidone safety and adverse event management in idiopathic pulmonary fibrosis. Eur. Respir. Rev. 2017, 26. [Google Scholar] [CrossRef] [Green Version]
- Distler, O.; Highland, K.B.; Gahlemann, M.; Azuma, A.; Fischer, A.; Mayes, M.D.; Raghu, G.; Sauter, W.; Girard, M.; Alves, M.; et al. Nintedanib for systemic sclerosis-associated interstitial lung disease. N. Engl. J. Med. 2019, 380, 2518–2528. [Google Scholar] [CrossRef]
- Ong, C.H.; Tham, C.L.; Harith, H.H.; Firdaus, N.; Israf, D.A. TGF-beta-induced fibrosis: A review on the underlying mechanism and potential therapeutic strategies. Eur. J. Pharmacol. 2021, 911, 174510. [Google Scholar] [CrossRef]
- D’Alessandro-Gabazza, C.N.; Kobayashi, T.; Boveda-Ruiz, D.; Takagi, T.; Toda, M.; Gil-Bernabe, P.; Miyake, Y.; Yasukawa, A.; Matsuda, Y.; Suzuki, N.; et al. Development and preclinical efficacy of novel transforming growth factor-beta1 short interfering RNAs for pulmonary fibrosis. Am. J. Respir. Cell Mol. Biol. 2012, 46, 397–406. [Google Scholar] [CrossRef]
- Wynn, T.A. Cellular and molecular mechanisms of fibrosis. J. Pathol. 2008, 214, 199–210. [Google Scholar] [CrossRef] [Green Version]
- Munger, J.S.; Huang, X.; Kawakatsu, H.; Griffiths, M.J.; Dalton, S.L.; Wu, J.; Pittet, J.F.; Kaminski, N.; Garat, C.; Matthay, M.A.; et al. The integrin alpha v beta 6 binds and activates latent TGF beta 1: A mechanism for regulating pulmonary inflammation and fibrosis. Cell 1999, 96, 319–328. [Google Scholar] [CrossRef] [Green Version]
- Annes, J.P.; Munger, J.S.; Rifkin, D.B. Making sense of latent TGFbeta activation. J. Cell Sci. 2003, 116, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Ruigrok, M.J.R.; Frijlink, H.W.; Melgert, B.N.; Olinga, P.; Hinrichs, W.L.J. Gene therapy strategies for idiopathic pulmonary fibrosis: Recent advances, current challenges, and future directions. Mol. Ther. Methods Clin. Dev. 2021, 20, 483–496. [Google Scholar] [CrossRef]
- Kukowska-Latallo, J.F.; Raczka, E.; Quintana, A.; Chen, C.; Rymaszewski, M.; Baker, J.R., Jr. Intravascular and endobronchial DNA delivery to murine lung tissue using a novel, nonviral vector. Hum. Gene Ther. 2000, 11, 1385–1395. [Google Scholar] [CrossRef]
- Qiu, Y.; Lam, J.K.; Leung, S.W.; Liang, W. Delivery of RNAi therapeutics to the airways-from bench to bedside. Molecules 2016, 21, 1249. [Google Scholar] [CrossRef] [Green Version]
- Kurosaki, T.; Kishikawa, R.; Matsumoto, M.; Kodama, Y.; Hamamoto, T.; To, H.; Niidome, T.; Takayama, K.; Kitahara, T.; Sasaki, H. Pulmonary gene delivery of hybrid vector, lipopolyplex containing N-lauroylsarcosine, via the systemic route. J. Control. Release 2009, 136, 213–219. [Google Scholar] [CrossRef] [Green Version]
- Abraham, J.; Bhat, S.G. Permeabilization of baker’s yeast with N-lauroyl sarcosine. J. Ind. Microbiol. Biotechnol. 2008, 35, 799–804. [Google Scholar] [CrossRef]
- Benjaminsen, R.V.; Mattebjerg, M.A.; Henriksen, J.R.; Moghimi, S.M.; Andresen, T.L. The possible “proton sponge” effect of polyethylenimine (PEI) does not include change in lysosomal pH. Mol. Ther. 2013, 21, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Vancha, A.R.; Govindaraju, S.; Parsa, K.V.; Jasti, M.; Gonzalez-Garcia, M.; Ballestero, R.P. Use of polyethyleneimine polymer in cell culture as attachment factor and lipofection enhancer. BMC Biotechnol. 2004, 4, 23. [Google Scholar] [CrossRef] [Green Version]
- Kodama, Y.; Nishigaki, W.; Nakamura, T.; Fumoto, S.; Nishida, K.; Kurosaki, T.; Nakagawa, H.; Kitahara, T.; Muro, T.; Sasaki, H. Splenic delivery system of pDNA through complexes electrostatically constructed with protamine and chondroitin sulfate. Biol. Pharm. Bull. 2018, 41, 342–349. [Google Scholar] [CrossRef] [Green Version]
- Yamagata, M.; Kawano, T.; Shiba, K.; Mori, T.; Katayama, Y.; Niidome, T. Structural advantage of dendritic poly(L-lysine) for gene delivery into cells. Bioorg. Med. Chem. 2007, 15, 526–532. [Google Scholar] [CrossRef]
- Kodama, Y.; Nakamura, T.; Kurosaki, T.; Egashira, K.; Mine, T.; Nakagawa, H.; Muro, T.; Kitahara, T.; Higuchi, N.; Sasaki, H. Biodegradable nanoparticles composed of dendrigraft poly-L-lysine for gene delivery. Eur. J. Pharm. Biopharm. 2014, 87, 472–479. [Google Scholar] [CrossRef] [Green Version]
- Stamatatos, L.; Leventis, R.; Zuckermann, M.J.; Silvius, J.R. Interactions of cationic lipid vesicles with negatively charged phospholipid vesicles and biological membranes. Biochemistry 1988, 27, 3917–3925. [Google Scholar] [CrossRef]
- Hafez, I.M.; Maurer, N.; Cullis, P.R. On the mechanism whereby cationic lipids promote intracellular delivery of polynucleic acids. Gene Ther. 2001, 8, 1188–1196. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Huang, L. A novel cationic liposome reagent for efficient transfection of mammalian cells. Biochem. Biophys. Res. Commun. 1991, 179, 280–285. [Google Scholar] [CrossRef]
- Mu, J.; Lin, J.; Huang, P.; Chen, X. Development of endogenous enzyme-responsive nanomaterials for theranostics. Chem. Soc. Rev. 2018, 47, 5554–5573. [Google Scholar] [CrossRef]
- Moore, C.B.; Guthrie, E.H.; Huang, M.T.; Taxman, D.J. Short hairpin RNA (shRNA): Design, delivery, and assessment of gene knockdown. Methods Mol. Biol. 2010, 629, 141–158. [Google Scholar] [CrossRef] [Green Version]
- Grimm, D.; Streetz, K.L.; Jopling, C.L.; Storm, T.A.; Pandey, K.; Davis, C.R.; Marion, P.; Salazar, F.; Kay, M.A. Fatality in mice due to oversaturation of cellular microRNA/short hairpin RNA pathways. Nature 2006, 441, 537–541. [Google Scholar] [CrossRef]
- Suhy, D.A.; Kao, S.C.; Mao, T.; Whiteley, L.; Denise, H.; Souberbielle, B.; Burdick, A.D.; Hayes, K.; Wright, J.F.; Lavender, H.; et al. Safe, long-term hepatic expression of anti-HCV shRNA in a nonhuman primate model. Mol. Ther. 2012, 20, 1737–1749. [Google Scholar] [CrossRef] [Green Version]
- Ruscitti, F.; Ravanetti, F.; Bertani, V.; Ragionieri, L.; Mecozzi, L.; Sverzellati, N.; Silva, M.; Ruffini, L.; Menozzi, V.; Civelli, M.; et al. Quantification of Lung Fibrosis in IPF-Like Mouse Model and Pharmacological Response to Treatment by Micro-Computed Tomography. Front. Pharmacol. 2020, 11, 1117. [Google Scholar] [CrossRef]
- Bondue, B.; Castiaux, A.; Van Simaeys, G.; Mathey, C.; Sherer, F.; Egrise, D.; Lacroix, S.; Huaux, F.; Doumont, G.; Goldman, S. Absence of early metabolic response assessed by 18F-FDG PET/CT after initiation of antifibrotic drugs in IPF patients. Respir. Res. 2019, 20, 10. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Complex | Size (nm) | PDI | ζ-Potential (mV) |
|---|---|---|---|
| pDNA/PEI/DOTMA/LS | 185.1 ± 5.87 | 0.24 ± 0.01 | 16.7 ± 0.50 |
| pDNA/PLL/DOTMA/LS | 169.7 ± 4.71 | 0.23 ± 0.01 | 14.5 ± 0.10 |
| pDNA/PLA/DOTMA/LS | 179.5 ± 2.50 | 0.21 ± 0.01 | 17.8 ± 0.46 |
| pDNA/DGL/DOTMA/LS | 211.6 ± 2.20 | 0.26 ± 0.01 | 11.3 ± 0.36 |
| Complex | Size (nm) | PDI | ζ-Potential (mV) |
|---|---|---|---|
| pDNA/DGL/DOTMA/LS | 211.6 ± 2.20 | 0.25 ± 0.02 | 11.3 ± 0.36 |
| pDNA/DGL/DOTAP/LS | 221.5 ± 10.97 | 0.25 ± 0.03 | 12.3 ± 0.70 |
| pDNA/DGL/DODAP/LS | Aggregated | 0.74 ± 0.04 | 2.93 ± 0.70 |
| pDNA/DGL/DC-chol/LS | 202.7 ± 10.17 | 0.33 ± 0.03 | 28.9 ± 0.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurosaki, T.; Kanda, H.; Hashizume, J.; Sato, K.; Harasawa, H.; Nakamura, T.; Sasaki, H.; Kodama, Y. Delivery of pDNA to the Lung by Lipopolyplexes Using N-Lauroylsarcosine and Effect on the Pulmonary Fibrosis. Pharmaceutics 2021, 13, 1983. https://doi.org/10.3390/pharmaceutics13111983
Kurosaki T, Kanda H, Hashizume J, Sato K, Harasawa H, Nakamura T, Sasaki H, Kodama Y. Delivery of pDNA to the Lung by Lipopolyplexes Using N-Lauroylsarcosine and Effect on the Pulmonary Fibrosis. Pharmaceutics. 2021; 13(11):1983. https://doi.org/10.3390/pharmaceutics13111983
Chicago/Turabian StyleKurosaki, Tomoaki, Hiroki Kanda, Junya Hashizume, Kayoko Sato, Hitomi Harasawa, Tadahiro Nakamura, Hitoshi Sasaki, and Yukinobu Kodama. 2021. "Delivery of pDNA to the Lung by Lipopolyplexes Using N-Lauroylsarcosine and Effect on the Pulmonary Fibrosis" Pharmaceutics 13, no. 11: 1983. https://doi.org/10.3390/pharmaceutics13111983
APA StyleKurosaki, T., Kanda, H., Hashizume, J., Sato, K., Harasawa, H., Nakamura, T., Sasaki, H., & Kodama, Y. (2021). Delivery of pDNA to the Lung by Lipopolyplexes Using N-Lauroylsarcosine and Effect on the Pulmonary Fibrosis. Pharmaceutics, 13(11), 1983. https://doi.org/10.3390/pharmaceutics13111983