
Negligible Effect of Quercetin in the Pharmacokinetics of Sulfasalazine in Rats and Beagles: Metabolic Inactivation of the Interaction Potential of Quercetin with BCRP

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Animal
2.3. RNA Extraction and Quantitative PCR
2.4. Cell Culture
2.5. Changes in mRNA Levels of BCRP Expressed in LS174T Cells and Rat BCRP Expressed in the Liver, Kidney, and Small Intestine of Rats following Exposure to Quercetin
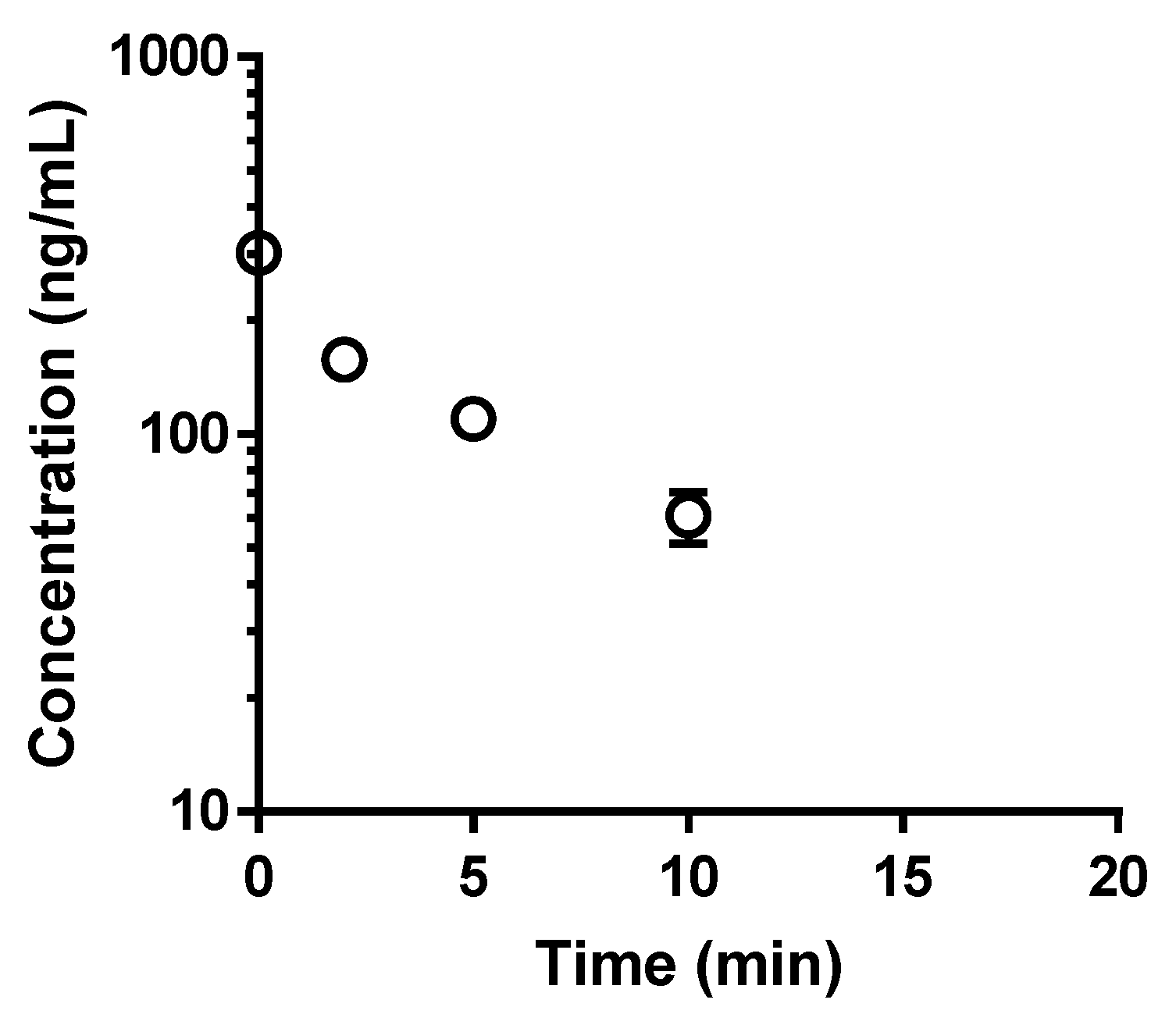
2.6. Metabolic Degradation of Quercetin in Isolated Rat Hepatocytes
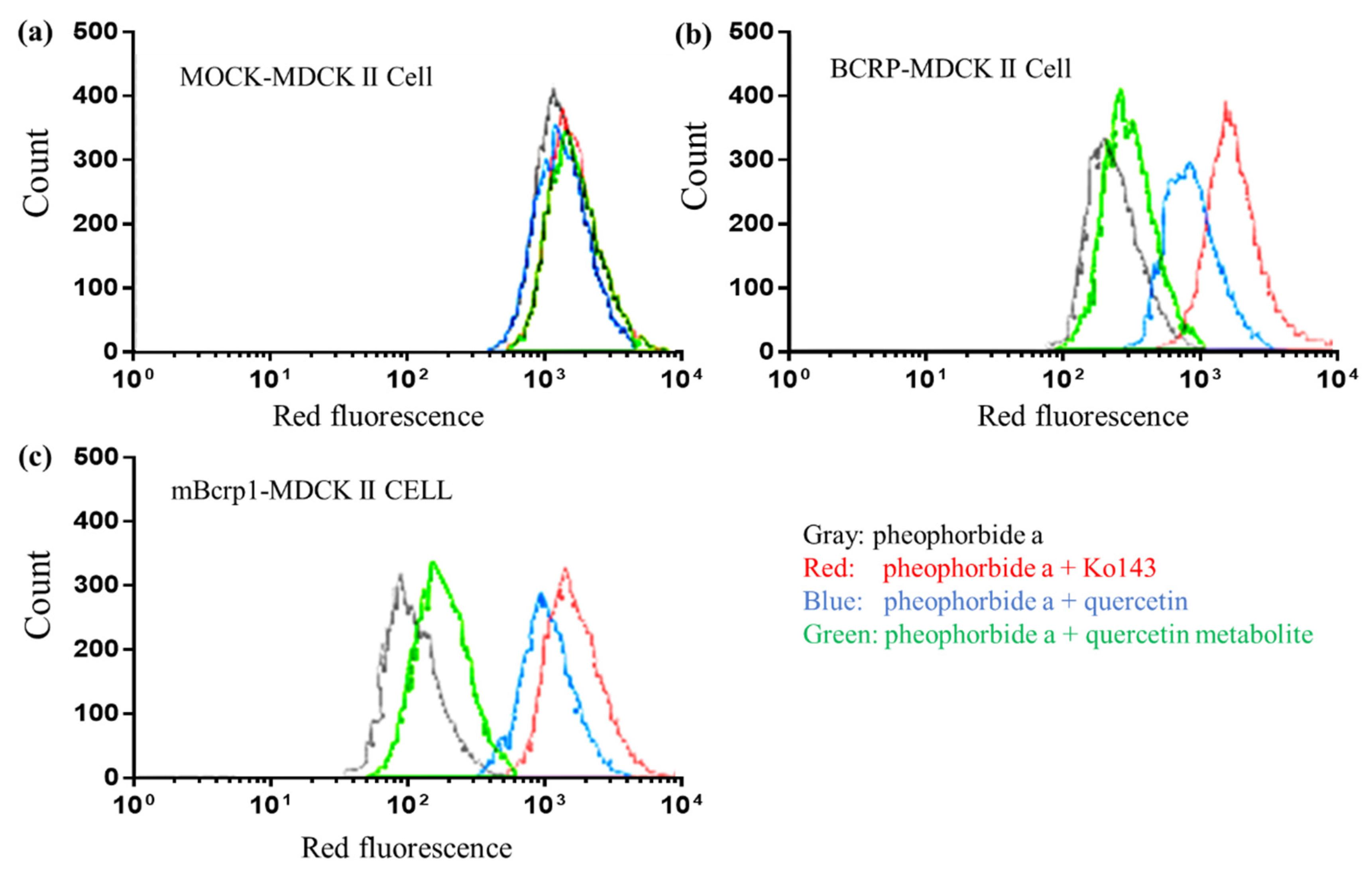
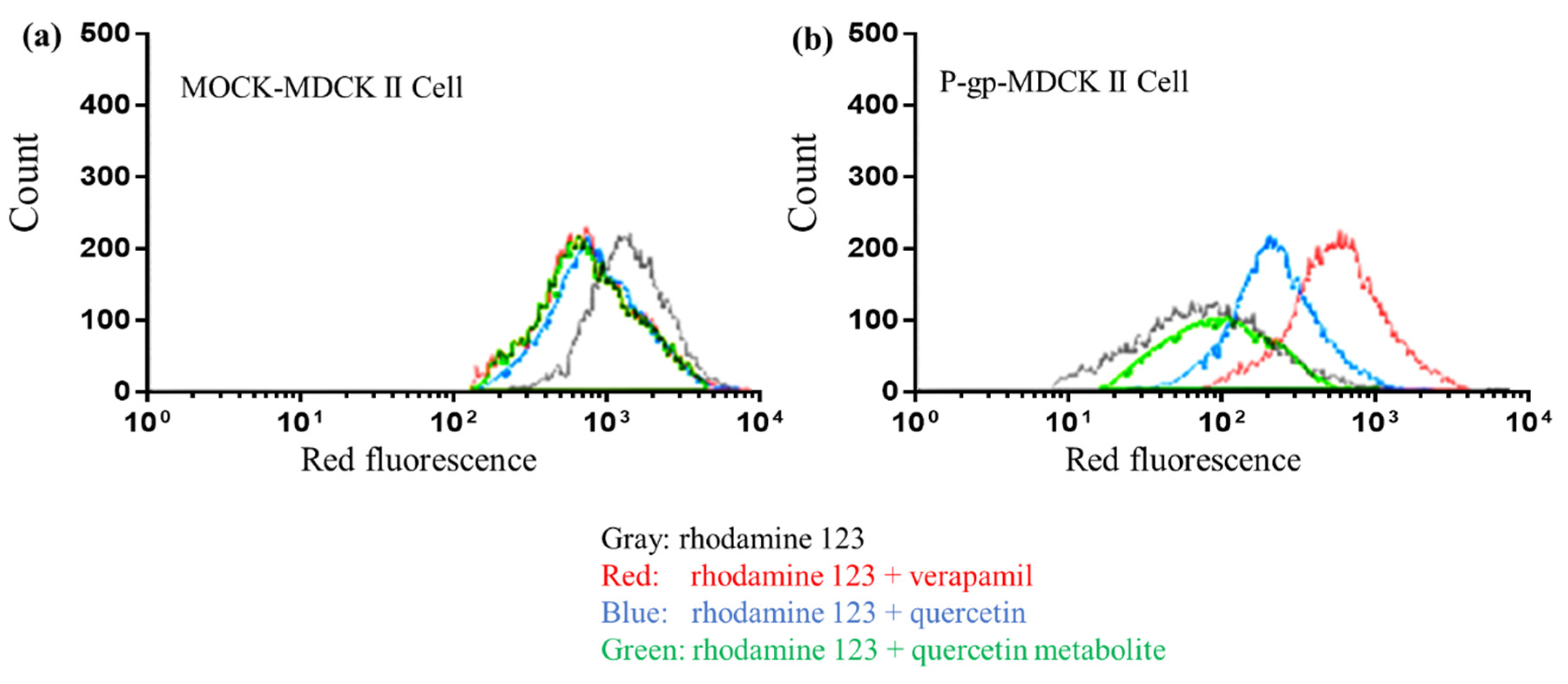
2.7. Inhibitory Effect of Quercetin and Its Metabolites on BCRP, mBcrp1, and P-gp Using Flow Cytometry
2.8. Pharmacokinetic Study of Sulfasalazine in Rats and Beagles
2.9. LC-MS/MS Analysis
2.10. Statistical Analysis
3. Results
3.1. Modulation of the Expression Level of BCRP by Administration of Quercetin
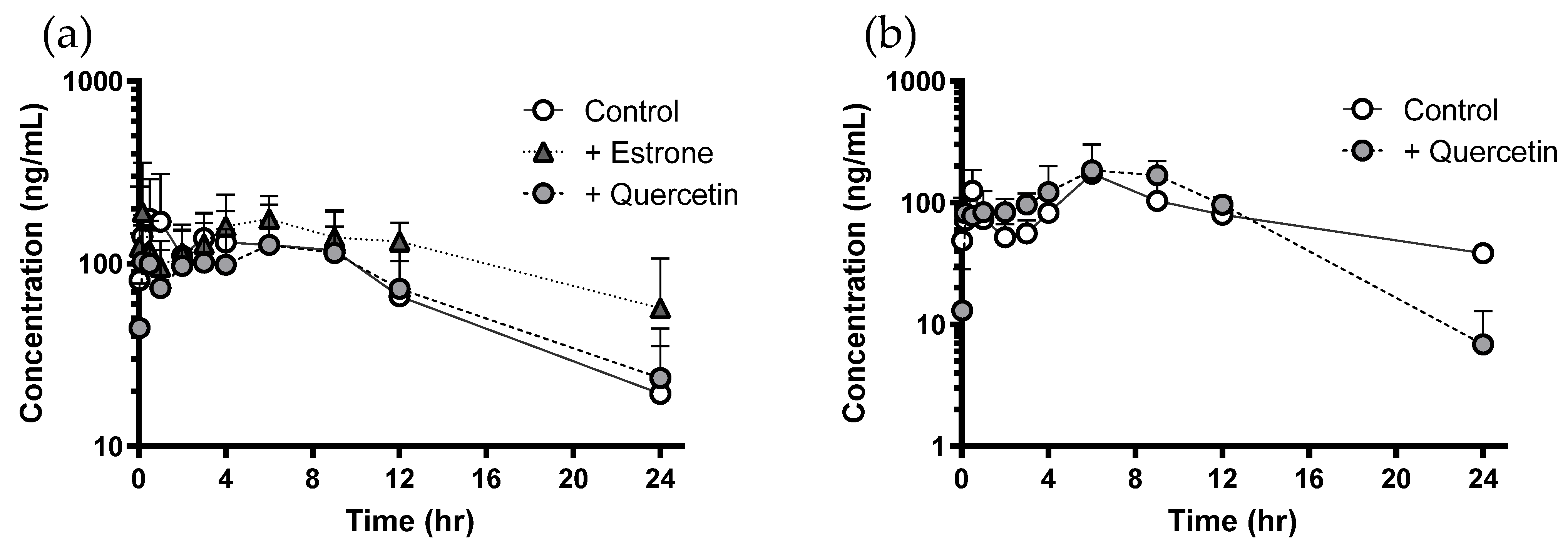
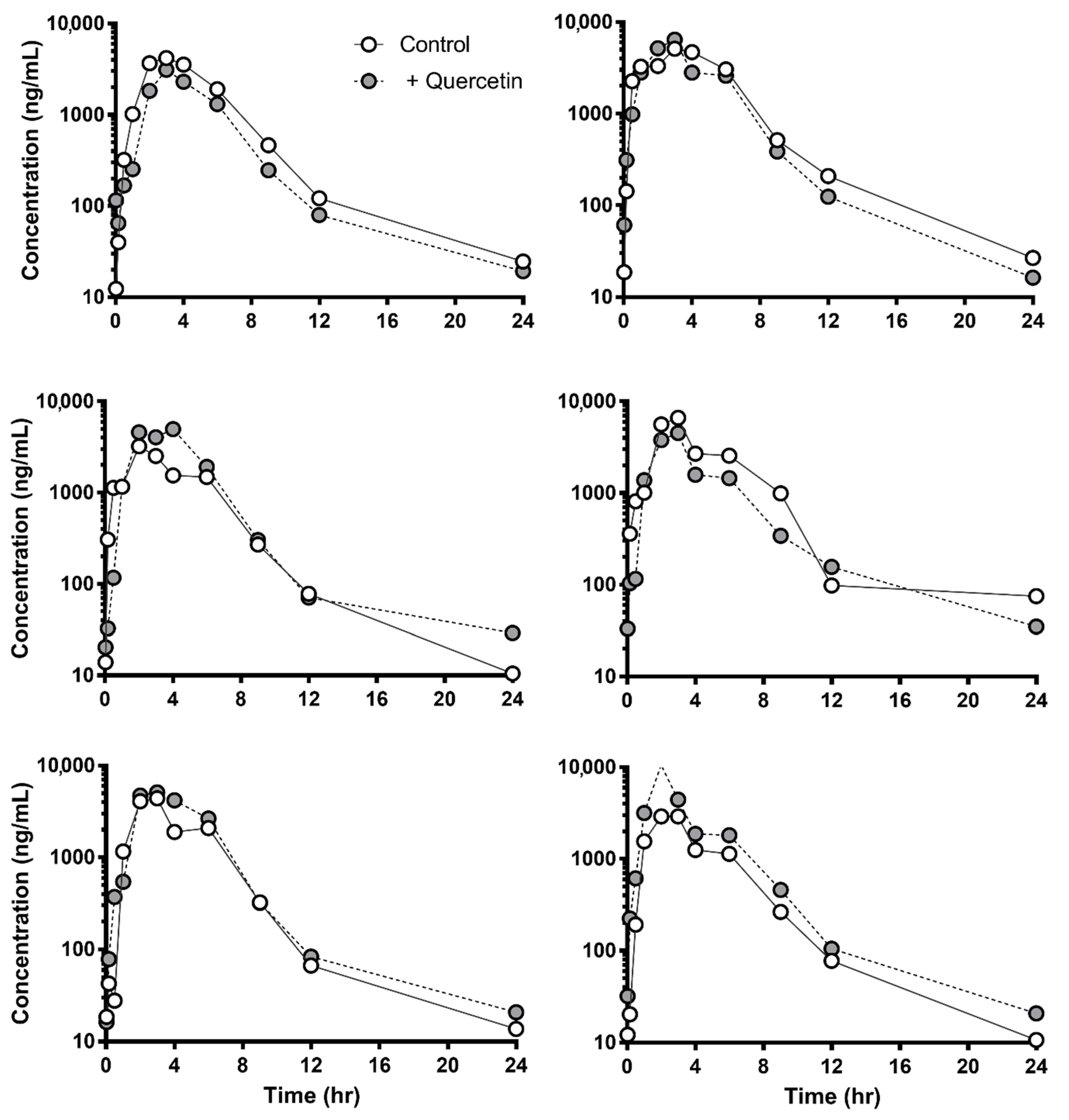
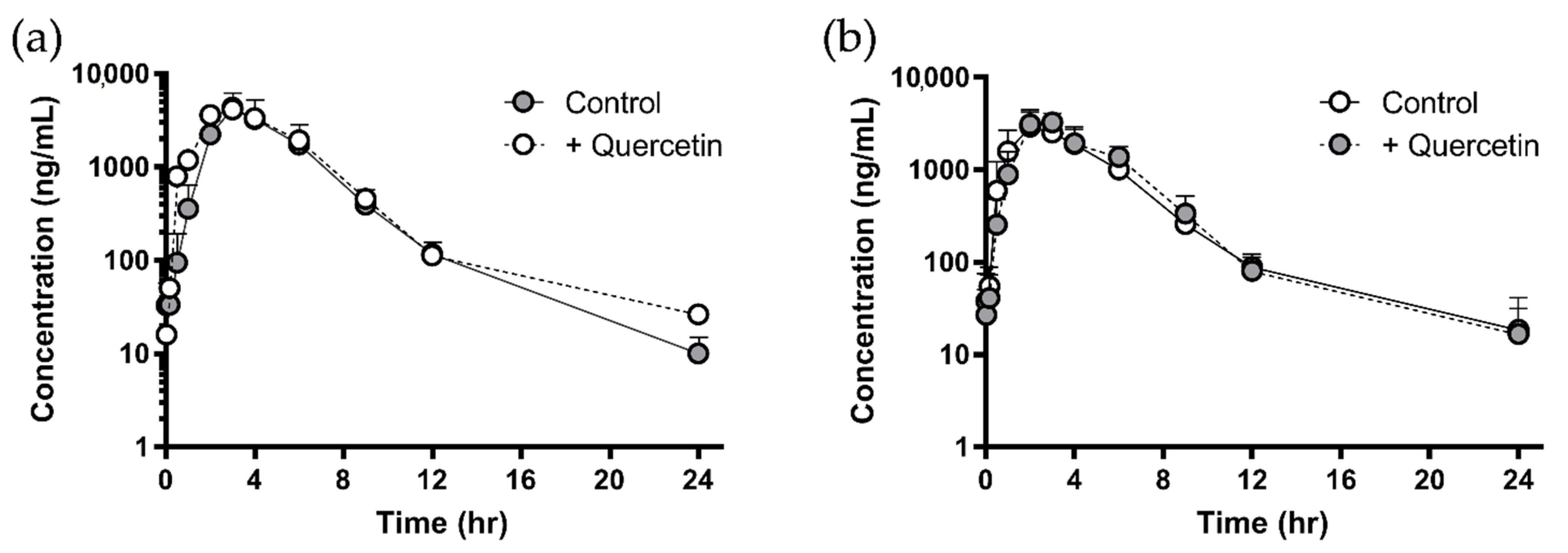
3.2. Negligible In Vivo Pharmacokinetic Change of Sulfasalazine Followed by Quercetin Administration
3.3. Loss of BCRP Inhibitory Activity by Metabolic Degradation of Quercetin
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Tarirai, C.; Viljoen, A.M.; Hamman, J.H. Herb–drug pharmacokinetic interactions reviewed. Expert Opin. Drug Metab. Toxicol. 2010, 6, 1515–1538. [Google Scholar] [CrossRef] [PubMed]
- De Smet, P.A.G.M. Clinical risk management of herb-drug interactions. Br. J. Clin. Pharmacol. 2007, 63, 258–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pangeni, R.; Kang, S.; Jha, S.K.; Subedi, L.; Park, J.W. Intestinal membrane transporter-mediated approaches to improve oral drug delivery. J. Pharm. Investig. 2021, 51, 137–158. [Google Scholar] [CrossRef]
- Huls, M.; Brown, C.; Windass, A.; Sayer, R.; Heuvel, J.V.D.; Heemskerk, S.; Russel, F.; Masereeuw, R. The breast cancer resistance protein transporter ABCG2 is expressed in the human kidney proximal tubule apical membrane. Kidney Int. 2008, 73, 220–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robey, R.W.; To, K.K.; Polgar, O.; Dohse, M.; Fetsch, P.; Dean, M.; Bates, S.E. ABCG2: A perspective. Adv. Drug Deliv. Rev. 2009, 61, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Kusuhara, H.; Furuie, H.; Inano, A.; Sunagawa, A.; Yamada, S.; Wu, C.; Fukizawa, S.; Morimoto, N.; Ieiri, I.; Morishita, M.; et al. Pharmacokinetic interaction study of sulfasalazine in healthy subjects and the impact of curcumin as an in vivo inhibitor of bcrp. Br. J. Pharmacol. 2012, 166, 1793–1803. [Google Scholar] [CrossRef]
- Sampson, L.; Rimm, E.; Hollman, P.C.; de Vries, J.H.; Katan, M.B. Flavonol and Flavone Intakes in US Health Professionals. J. Am. Diet. Assoc. 2002, 102, 1414–1420. [Google Scholar] [CrossRef]
- Harwood, M.; Danielewska-Nikiel, B.; Borzelleca, J.; Flamm, G.; Williams, G.; Lines, T. A critical review of the data related to the safety of quercetin and lack of evidence of in vivo toxicity, including lack of genotoxic/carcinogenic properties. Food Chem. Toxicol. 2007, 45, 2179–2205. [Google Scholar] [CrossRef]
- Bors, W.; Heller, W.; Michel, C.; Saran, M. [36] Flavonoids as antioxidants: Determination of radical-scavenging efficiencies. Methods Enzymol. 1990, 186, 343–355. [Google Scholar] [CrossRef]
- Vida, R.G.; Fittler, A.; Somogyi-Végh, A.; Poór, M. Dietary quercetin supplements: Assessment of online product informations and quantitation of quercetin in the products by high-performance liquid chromatography. Phytother. Res. 2019, 33, 1912–1920. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Chao, P.-D.L.; Hsiu, S.-L.; Wen, K.-C.; Hou, Y.-C. Lethal quercetin-digoxin interaction in pigs. Life Sci. 2004, 74, 1191–1197. [Google Scholar] [CrossRef]
- Brand, W.; Schutte, M.E.; Williamson, G.; van Zanden, J.J.; Cnubben, N.H.; Groten, J.P.; van Bladeren, P.J.; Rietjens, I. Flavonoid-mediated inhibition of intestinal ABC transporters may affect the oral bioavailability of drugs, food-borne toxic compounds and bioactive ingredients. Biomed. Pharmacother. 2006, 60, 508–519. [Google Scholar] [CrossRef] [PubMed]
- Cooray, H.C.; Janvilisri, T.; van Veen, H.W.; Hladky, S.B.; Barrand, M.A. Interaction of the breast cancer resistance protein with plant polyphenols. Biochem. Biophys. Res. Commun. 2004, 317, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yang, X.; Morris, M.E. Flavonoids Are Inhibitors of Breast Cancer Resistance Protein (ABCG2)-Mediated Transport. Mol. Pharmacol. 2004, 65, 1208–1216. [Google Scholar] [CrossRef] [Green Version]
- Sesink, A.L.; Arts, I.C.; de Boer, V.C.; Breedveld, P.; Schellens, J.H.; Hollman, P.C.; Russel, F.G. Breast cancer resistance protein (bcrp1/abcg2) limits net intestinal uptake of quercetin in rats by facilitating apical efflux of glucuronides. Mol. Pharmacol. 2005, 67, 1999–2006. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, M.; Ikegami, Y.; Sano, K.; Yoshida, H.; Mitomo, H.; Sawada, S.; Ishikawa, T. Transport of SN-38 by the wild type of human ABC transporter ABCG2 and its inhibition by quercetin, a natural flavonoid. J. Exp. Ther. Oncol. 2004, 4, 25–35. [Google Scholar]
- Song, Y.-K.; Yoon, J.-H.; Woo, J.K.; Kang, J.-H.; Lee, K.-R.; Oh, S.H.; Chung, S.-J.; Maeng, H.-J. Quercetin Is a Flavonoid Breast Cancer Resistance Protein Inhibitor with an Impact on the Oral Pharmacokinetics of Sulfasalazine in Rats. Pharmaceutics 2020, 12, 397. [Google Scholar] [CrossRef]
- Bakos, É.; Evers, R.; Szakacs, G.; Tusnády, G.E.; Welker, E.; Szabó, K.; de Haas, M.; van Deemter, L.; Borst, P.; Váradi, A.; et al. Functional Multidrug Resistance Protein (MRP1) Lacking the N-terminal Transmembrane Domain. J. Biol. Chem. 1998, 273, 32167–32175. [Google Scholar] [CrossRef] [Green Version]
- Pavek, P.; Merino, G.; Wagenaar, E.; Bolscher, E.; Novotna, M.; Jonker, J.W.; Schinkel, A.H. Human Breast Cancer Resistance Protein: Interactions with Steroid Drugs, Hormones, the Dietary Carcinogen 2-Amino-1-methyl-6-phenylimidazo(4,5-b)pyridine, and Transport of Cimetidine. J. Pharmacol. Exp. Ther. 2005, 312, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Jonker, J.; Smit, J.W.; Brinkhuis, R.F.; Maliepaard, M.; Beijnen, J.H.; Schellens, J.H.M.; Schinkel, A.H. Role of Breast Cancer Resistance Protein in the Bioavailability and Fetal Penetration of Topotecan. J. Natl. Cancer Inst. 2000, 92, 1651–1656. [Google Scholar] [CrossRef] [PubMed]
- Uno, S.; Uraki, M.; Ito, A.; Shinozaki, Y.; Yamada, A.; Kawase, A.; Iwaki, M. Changes in mRNA expression of ABC and SLC transporters in liver and intestines of the adjuvant-induced arthritis rat. Biopharm. Drug Dispos. 2009, 30, 49–54. [Google Scholar] [CrossRef]
- Declèves, X.; Bihorel, S.; Debray, M.; Yousif, S.; Camenisch, G.; Scherrmann, J.-M. ABC transporters and the accumulation of imatinib and its active metabolite CGP74588 in rat C6 glioma cells. Pharmacol. Res. 2008, 57, 214–222. [Google Scholar] [CrossRef]
- Brand, W.; Van Der Wel, P.A.; Rein, M.J.; Barron, D.; Williamson, G.; Bladeren, P.J.; Rietjens, I.M. Metabolism and Transport of the Citrus Flavonoid Hesperetin in Caco-2 Cell Monolayers. Drug Metab. Dispos. 2008, 36, 1794–1802. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.-H.; Lee, J.H.; Lee, Y.-J. Evaluation of the Mrp2-mediated flavonoid-drug interaction potential of quercetin in rats and in vitro models. Asian J. Pharm. Sci. 2019, 14, 621–630. [Google Scholar] [CrossRef]
- Berry, M.N.; Friend, D.S. High-yield preparation of isolated rat liver parenchymal cells: A biochemical and fine structural study. J. Cell Biol. 1969, 43, 506–520. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Hillebrand, A.; Wang, D.Q.-H.; Liu, M. Isolation and Primary Culture of Rat Hepatic Cells. J. Vis. Exp. 2012, 64, e3917. [Google Scholar] [CrossRef] [PubMed]
- Robey, R.; Honjo, Y.; van de Laar, A.; Miyake, K.; Regis, J.T.; Litman, T.; Bates, S.E. A functional assay for detection of the mitoxantrone resistance protein, MXR (ABCG2). Biochim. Biophys. Acta Biomembr. 2001, 1512, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Robey, R.W.; Steadman, K.; Polgar, O.; Morisaki, K.; Blayney, M.; Mistry, P.; Bates, S.E. Pheophorbide a Is a Specific Probe for ABCG2 Function and Inhibition. Cancer Res. 2004, 64, 1242–1246. [Google Scholar] [CrossRef] [Green Version]
- Tsukahara, S.; Imai, Y.; Sugimoto, Y.; Ueda, K.; Tsuruo, T. Reversal of breast cancer resistance protein-mediated drug resistance by estrogen antagonists and agonists. Mol. Cancer Ther. 2003, 2, 105–112. [Google Scholar]
- Zamek-Gliszczynski, M.J.; Bedwell, D.W.; Bao, J.Q.; Higgins, J.W. Characterization of SAGE Mdr1a (P-gp), Bcrp, and Mrp2 Knockout Rats Using Loperamide, Paclitaxel, Sulfasalazine, and Carboxydichlorofluorescein Pharmacokinetics. Drug Metab. Dispos. 2012, 40, 1825–1833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urquhart, B.L.; Ware, J.A.; Tirona, R.G.; Ho, R.H.; Leake, B.F.; Schwarz, U.I.; Zaher, H.; Palandra, J.; Gregor, J.C.; Dresser, G.K.; et al. Breast cancer resistance protein (ABCG2) and drug disposition: Intestinal expression, polymorphisms and sulfasalazine as an in vivo probe. Pharm. Genom. 2008, 18, 439–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaher, H.; Khan, A.A.; Palandra, J.; Brayman, T.G.; Yu, L.; Ware, J.A. Breast Cancer Resistance Protein (Bcrp/abcg2) Is a Major Determinant of Sulfasalazine Absorption and Elimination in the Mouse. Mol. Pharm. 2006, 3, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Oh, J.H.; Lee, J.; Lee, Y.J. Minor effects of the Citrus flavonoids naringin, naringenin and quercetin, on the pharmacokinetics of doxorubicin in rats. Pharmazie 2011, 66, 424–429. [Google Scholar] [PubMed]
- Jani, M.; Szabó, P.; Kis, E.; Molnár, É.; Glavinas, H.; Krajcsi, P. Kinetic Characterization of Sulfasalazine Transport by Human ATP-Binding Cassette G2. Biol. Pharm. Bull. 2009, 32, 497–499. [Google Scholar] [CrossRef] [Green Version]
- MacLean, C.; Moenning, U.; Reichel, A.; Fricker, G. Closing the Gaps: A Full Scan of the Intestinal Expression of P-Glycoprotein, Breast Cancer Resistance Protein, and Multidrug Resistance-Associated Protein 2 in Male and Female Rats. Drug Metab. Dispos. 2008, 36, 1249–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azadkhan, A.K.; Truelove, S.C.; Aronson, J.K. The disposition and metabolism of sulphasalazine (salicylazosulphapyridine) in man. Br. J. Clin. Pharmacol. 1982, 13, 523–528. [Google Scholar] [CrossRef]
- Chungi, V.S.; Dittert, L.W.; Shargel, L. Pharmacokinetics of Sulfasalazine Metabolites in Rats Following Concomitant Oral Administration of Riboflavin. Pharm. Res. 1989, 6, 1067–1072. [Google Scholar] [CrossRef]
- Crespy, V.; Morand, C.; Manach, C.; Besson, C.; Demigne, C.; Remesy, C. Part of quercetin absorbed in the small intestine is conjugated and further secreted in the intestinal lumen. Am. J. Physiol. Content 1999, 277, G120–G126. [Google Scholar] [CrossRef] [PubMed]
- Mullen, W.; Edwards, C.A.; Crozier, A. Absorption, excretion and metabolite profiling of methyl-, glucuronyl-, glucosyl- and sulpho-conjugates of quercetin in human plasma and urine after ingestion of onions. Br. J. Nutr. 2006, 96, 107–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, S.-L.; Lin, Y.-C.; Lin, Y.-L.; Li, C.-C.; Chuang, C.-H. Comparing the metabolism of quercetin in rats, mice and gerbils. Eur. J. Nutr. 2015, 55, 413–422. [Google Scholar] [CrossRef]
- Mohos, V.; Fliszár-Nyúl, E.; Ungvári, O.; Kuffa, K.; Needs, P.W.; Kroon, P.A.; Telbisz, Á.; Özvegy-Laczka, C.; Poór, M. Inhibitory Effects of Quercetin and Its Main Methyl, Sulfate, and Glucuronic Acid Conjugates on Cytochrome P450 Enzymes, and on OATP, BCRP and MRP2 Transporters. Nutrients 2020, 12, 2306. [Google Scholar] [CrossRef]
- Crespy, V.; Morand, C.; Besson, C.; Manach, C.; Demigne, C.; Remesy, C. Quercetin, but not Its Glycosides, Is Absorbed from the Rat Stomach. J. Agric. Food Chem. 2002, 50, 618–621. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Liu, Y.; Huang, X.; Zhang, R.; Yang, C.; Zhou, J.; Zhang, Y.; Wan, J.; Shi, S. Quercetin3obetadglucoside decreases the bioavailability of cyclosporin a through regulation of drug metabolizing enzymes, transporters and nuclear receptors in rats. Mol. Med. Rep. 2018, 18, 2599–2612. [Google Scholar] [PubMed] [Green Version]
- Östlund, J.; Žlábek, V.; Zamaratskaia, G. In vitro inhibition of human CYP2E1 and CYP3A by quercetin and myricetin in hepatic microsomes is not gender dependent. Toxicology 2017, 381, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Pal, D.; Mitra, A.K. MDR- and CYP3A4-Mediated Drug–Drug Interactions. J. Neuroimmune Pharmacol. 2006, 1, 323–339. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession No. | Sequence (5′-3′) | Condition |
|---|---|---|---|
| r18s RNA [21] | M11188 | FW- CGCCGCTAGAGGTGAAATTC RW- CCAGTCGGCATCGTTTATGG | 95 °C—10 s, 95 °C—5 s, 61 °C—7 s, 72 °C—10 s |
| Rat BCRP [22] | NM_181381 | FW- CAGCAGGTTACCACTGTGAG RW- TTCCCCTCTGTTTAACATTACA | 95 °C—10 s, 95 °C—5 s, 56 °C—6 s, 72 °C—6s |
| Human BCRP [23] | NM_004827.2 | FW- CAGGTCTGTTGGTCAATCTCACA RW- TCCATATCGTGGAATGCTGAAG | 95 °C—10 s, 95 °C—5 s, 58 °C—15 s, 72 °C—15 s |
| Single Administration (n = 6) | Multiple Administration (n = 3) | ||||
|---|---|---|---|---|---|
| Control | +Quercetin | +Estrone | Control | +Quercetin | |
| AUClast (μg∙h/mL) | 1.84 ± 0.557 | 1.65 ± 0.912 | 2.73 ± 0.301 ** | 1.87 ± 0.684 | 1.87 ± 0.367 |
| Cmax (ng/mL) | 226 ± 130 | 126 ± 64 | 277 ± 118 | 141 ± 34 | 111 ± 10 |
| Tmax (h) | 1.06 ± 1.47 | 0.506 ± 0.749 | 3.39 ± 3.61 | 0.389 ± 0.192 | 1.72 ± 1.44 |
| Single Administration (n = 6) | Multiple Administration (n = 3) | |||||
|---|---|---|---|---|---|---|
| 7-Day Treatment | Recovery Phase | |||||
| Control | +Quercetin | Control | +Quercetin | Control | +Quercetin | |
| AUClast (μg∙h/mL) | 21.0 ± 6.80 | 21.4 ± 5.63 | 18.1 ± 7.85 | 21.0 ± 7.20 | 13.7 ± 6.34 | 15.1 ± 3.15 |
| Cmax (μg/mL) | 4.41 ± 1.35 | 4.83 ± 1.19 | 4.40 ± 1.79 | 4.23 ± 1.09 | 3.19 ± 1.24 | 3.64 ± 0.916 |
| Tmax (h) | 2.83 ± 0.408 | 3.20 ± 0.447 | 3.33 ± 0.577 | 3.00 ± 1.00 | 2.00 ± 1.00 | 2.67 ± 0.577 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, J.-H.; Kim, D.; Lee, H.; Kim, G.; Park, T.; Kim, M.C.; Lee, Y.-J. Negligible Effect of Quercetin in the Pharmacokinetics of Sulfasalazine in Rats and Beagles: Metabolic Inactivation of the Interaction Potential of Quercetin with BCRP. Pharmaceutics 2021, 13, 1989. https://doi.org/10.3390/pharmaceutics13121989
Oh J-H, Kim D, Lee H, Kim G, Park T, Kim MC, Lee Y-J. Negligible Effect of Quercetin in the Pharmacokinetics of Sulfasalazine in Rats and Beagles: Metabolic Inactivation of the Interaction Potential of Quercetin with BCRP. Pharmaceutics. 2021; 13(12):1989. https://doi.org/10.3390/pharmaceutics13121989
Chicago/Turabian StyleOh, Ju-Hee, Dokeun Kim, Haejun Lee, Gyeonghee Kim, Taehoon Park, Min Chang Kim, and Young-Joo Lee. 2021. "Negligible Effect of Quercetin in the Pharmacokinetics of Sulfasalazine in Rats and Beagles: Metabolic Inactivation of the Interaction Potential of Quercetin with BCRP" Pharmaceutics 13, no. 12: 1989. https://doi.org/10.3390/pharmaceutics13121989
APA StyleOh, J. -H., Kim, D., Lee, H., Kim, G., Park, T., Kim, M. C., & Lee, Y. -J. (2021). Negligible Effect of Quercetin in the Pharmacokinetics of Sulfasalazine in Rats and Beagles: Metabolic Inactivation of the Interaction Potential of Quercetin with BCRP. Pharmaceutics, 13(12), 1989. https://doi.org/10.3390/pharmaceutics13121989