
Microbiota-Derived Short-Chain Fatty Acids: New Road in Colorectal Cancer Therapy



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Culture Conditions
2.2. Short-Chain Fatty Acids (SCFAs) Solutions
2.3. Sulforhodamine B Assay
2.4. Mixture Testing
2.5. Calculation of Predictive Mixture Effects
2.6. Regression Modeling and Statistical Analysis
2.7. Colony-Formation Assay
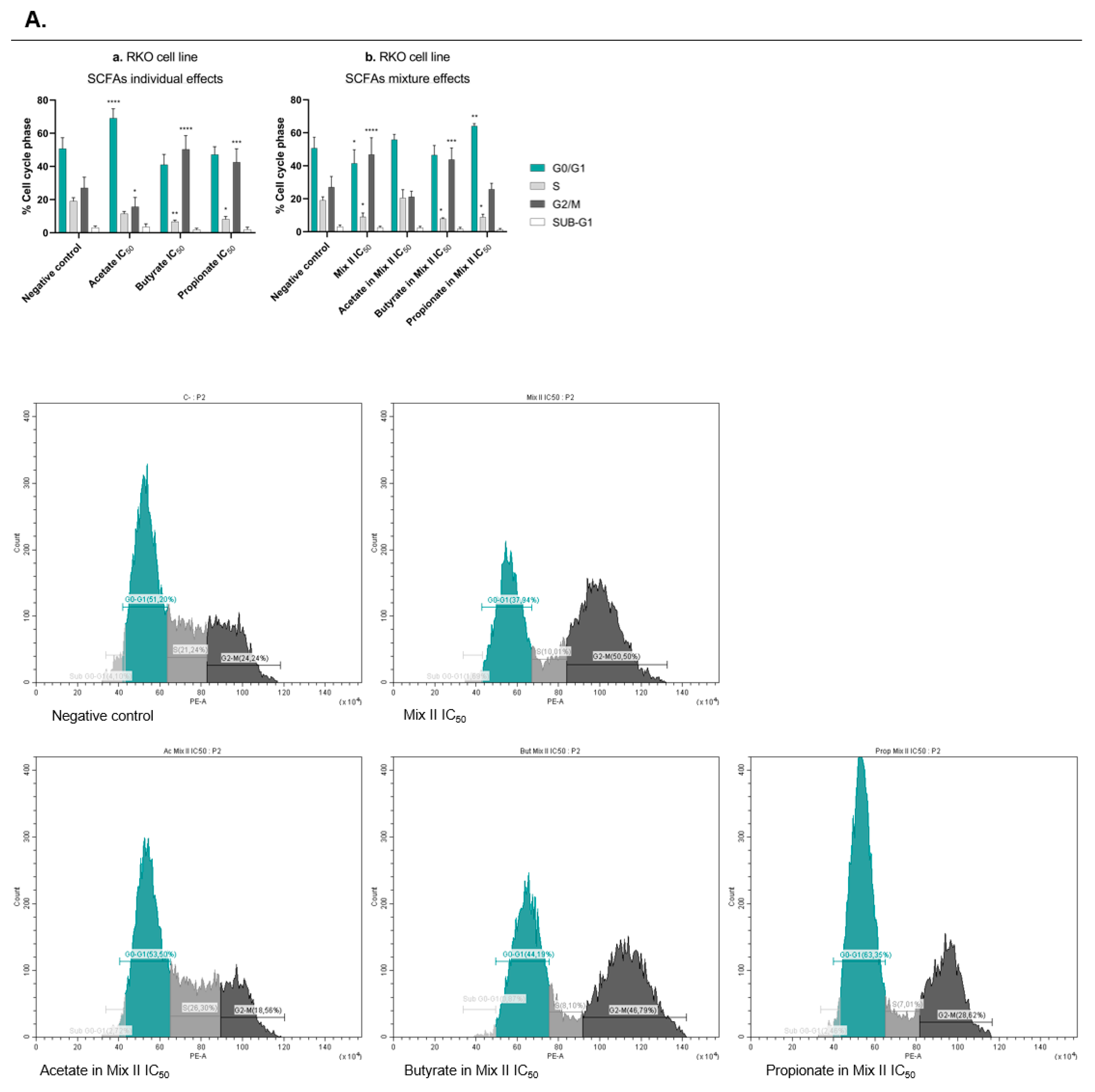
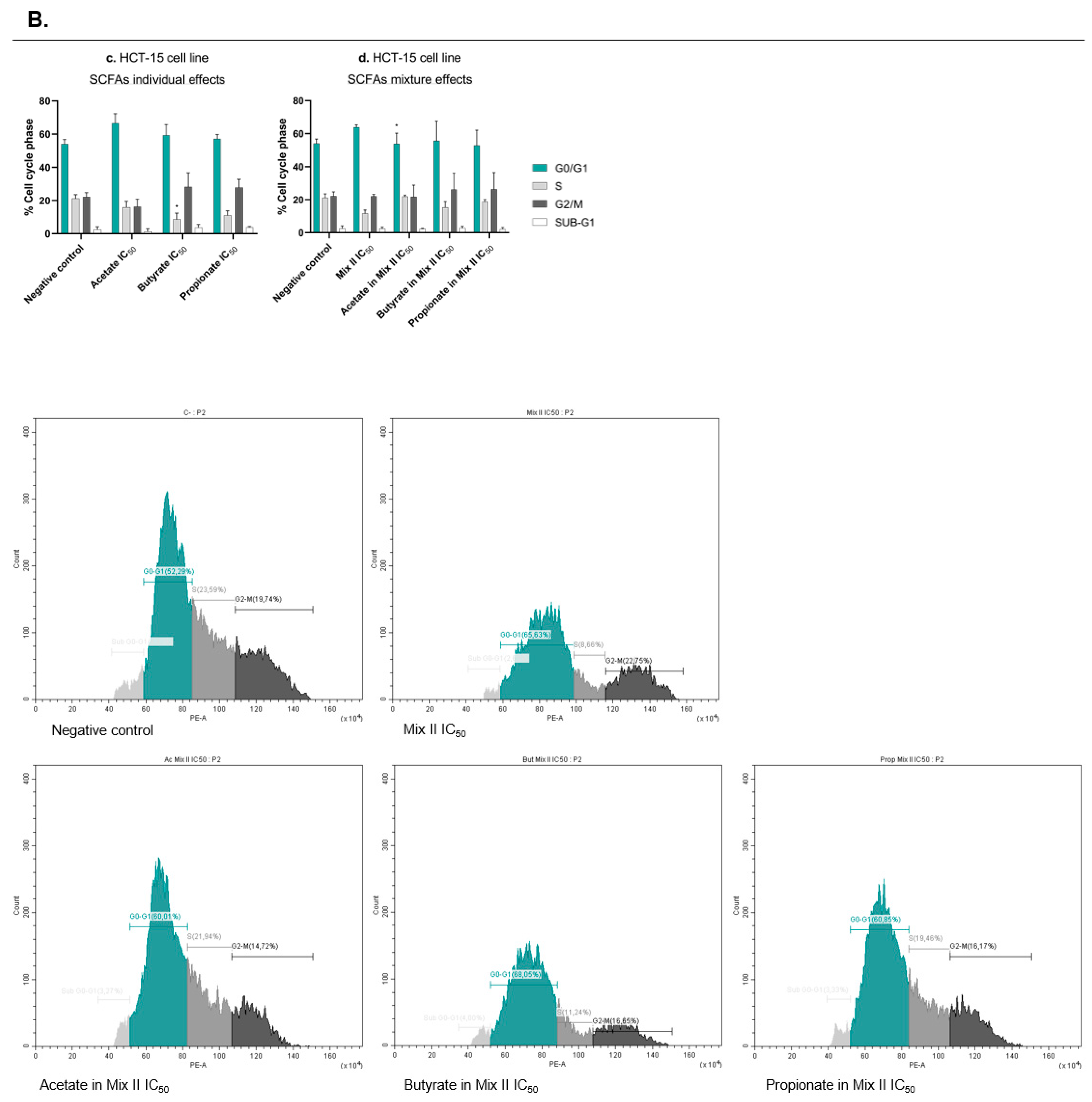
2.8. Cell-Cycle Analysis
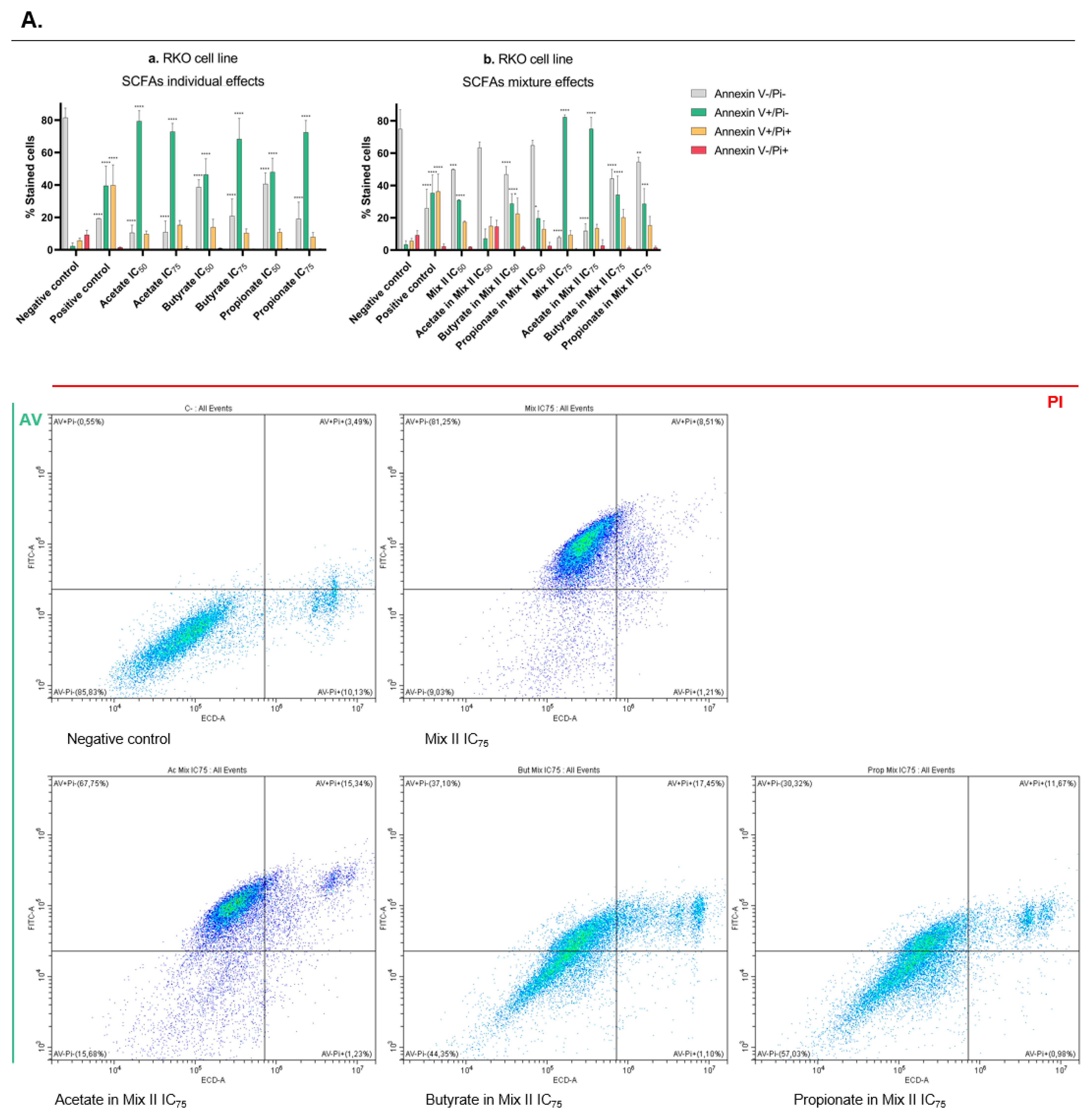
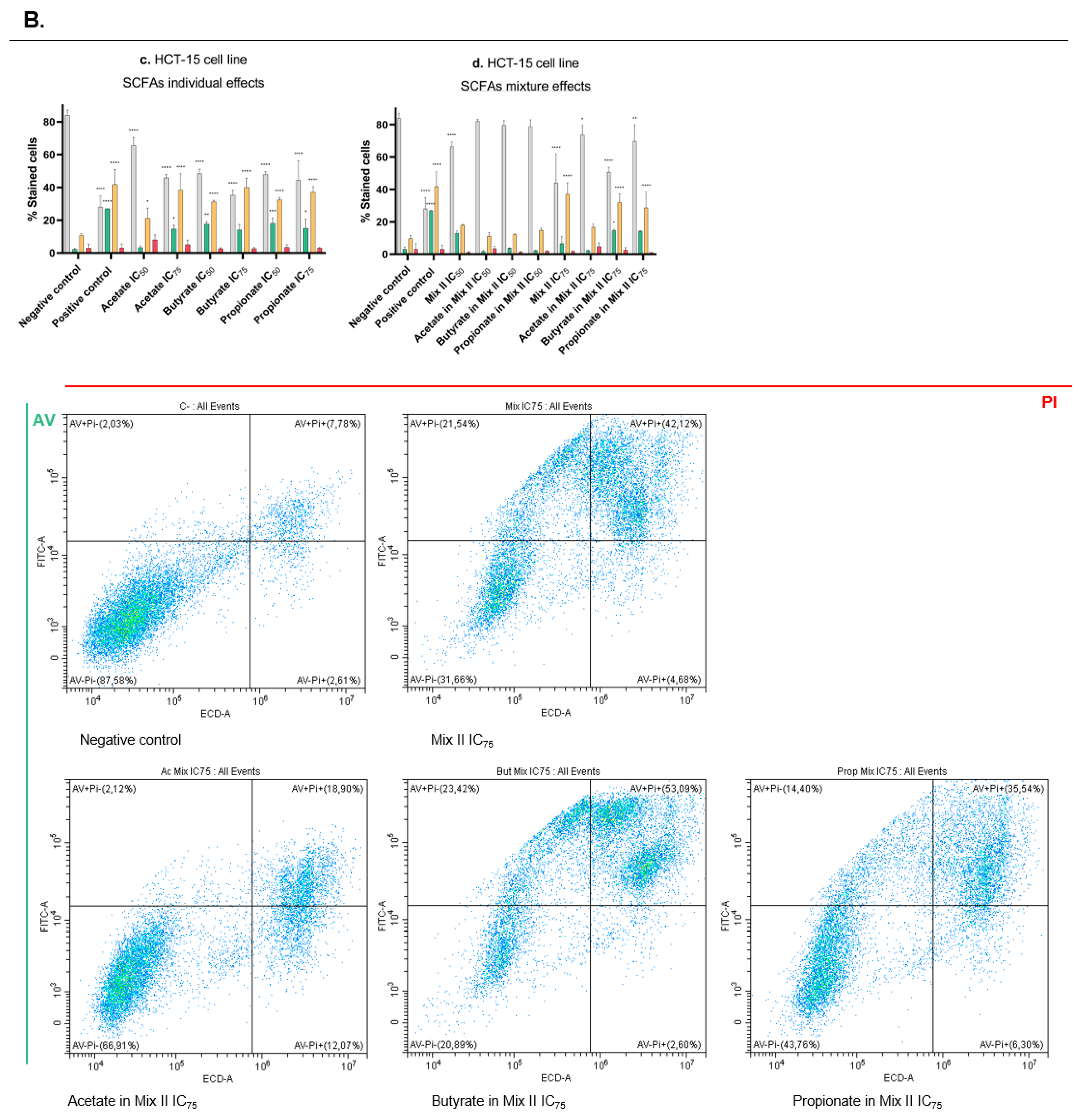
2.9. Annexin V/PI Staining Assay
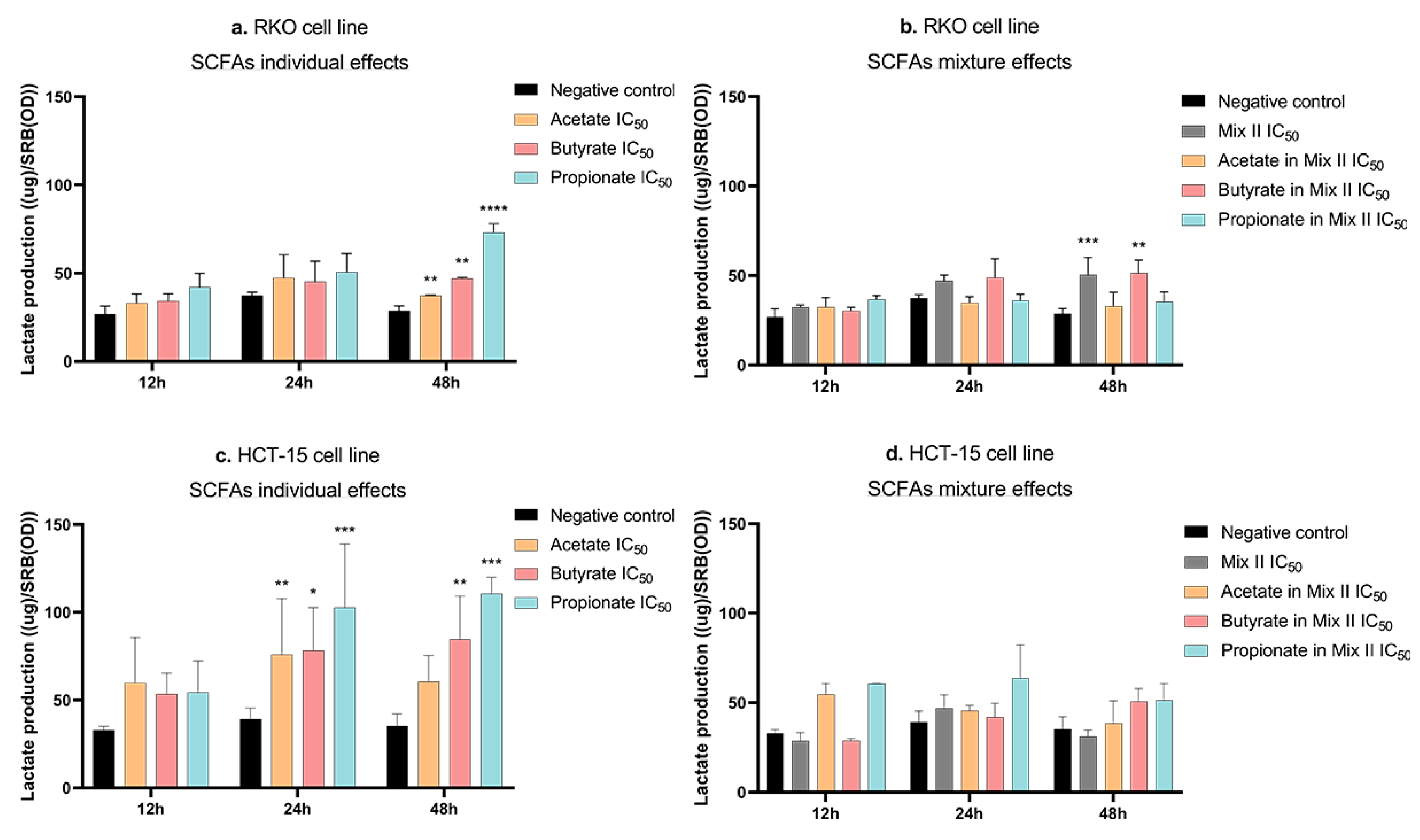
2.10. Lactate-Production Assay
2.11. Intracellular pH Evaluation
2.12. Lysosome Membrane Permeabilization Assessment
3. Results
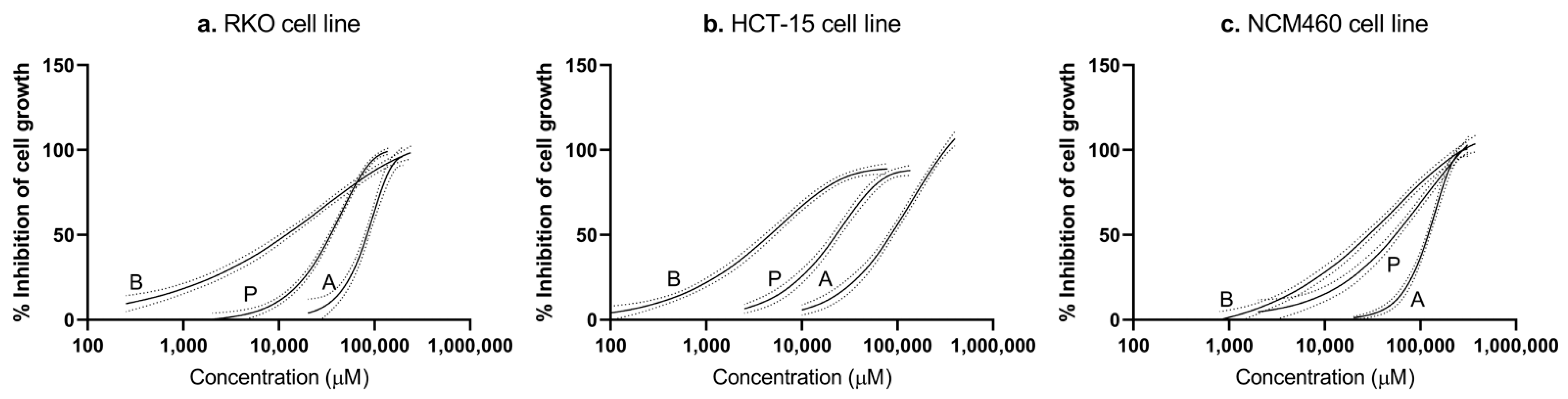
3.1. Short-Chain Fatty Acids Inhibit Cell Growth in a Dose-Dependent Manner with Higher Specificity to Colorectal Cancer Cells
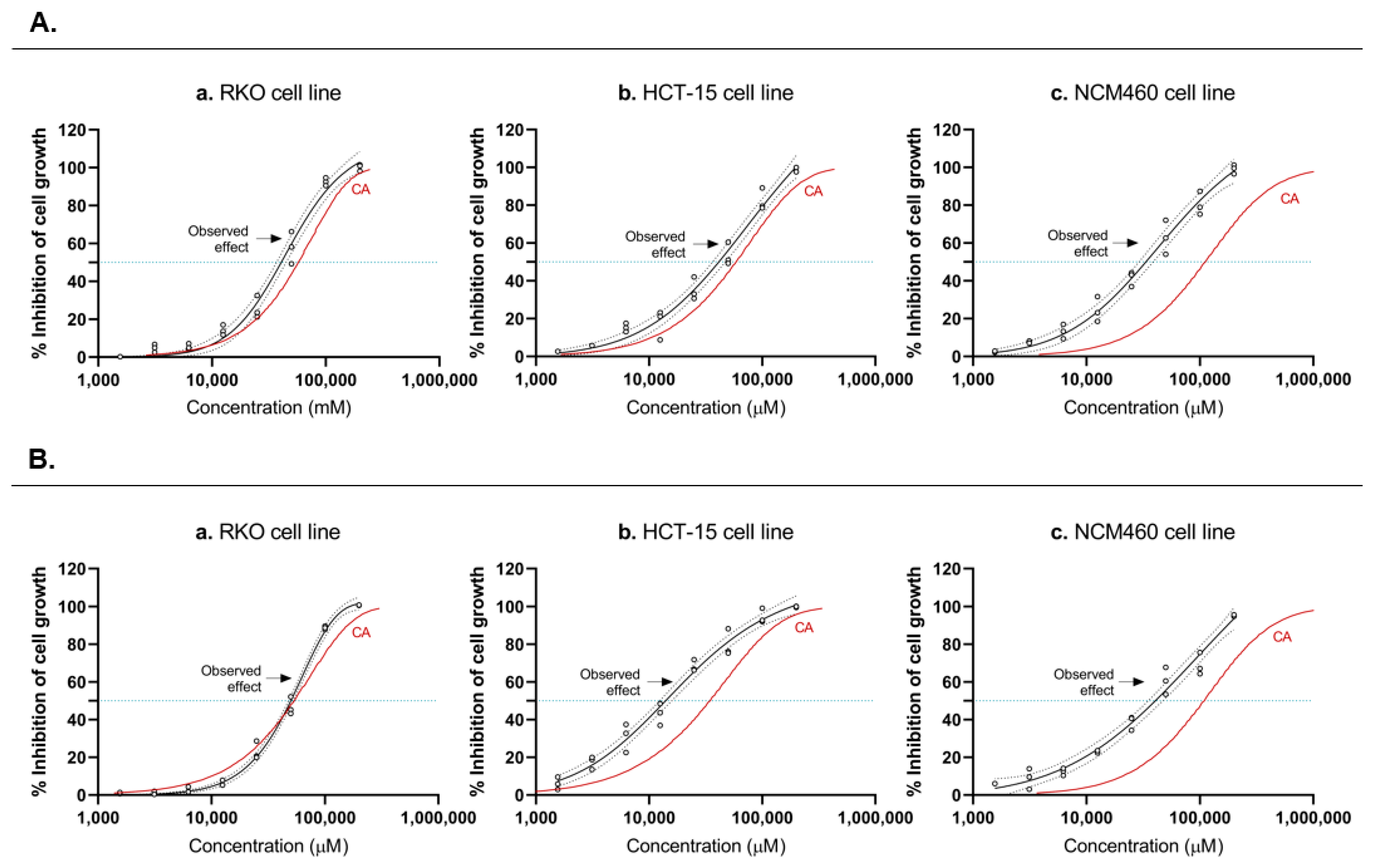
3.2. Short-Chain Fatty Acid Mixtures Exhibit an Addictive Profile in Colorectal Cancer Cells
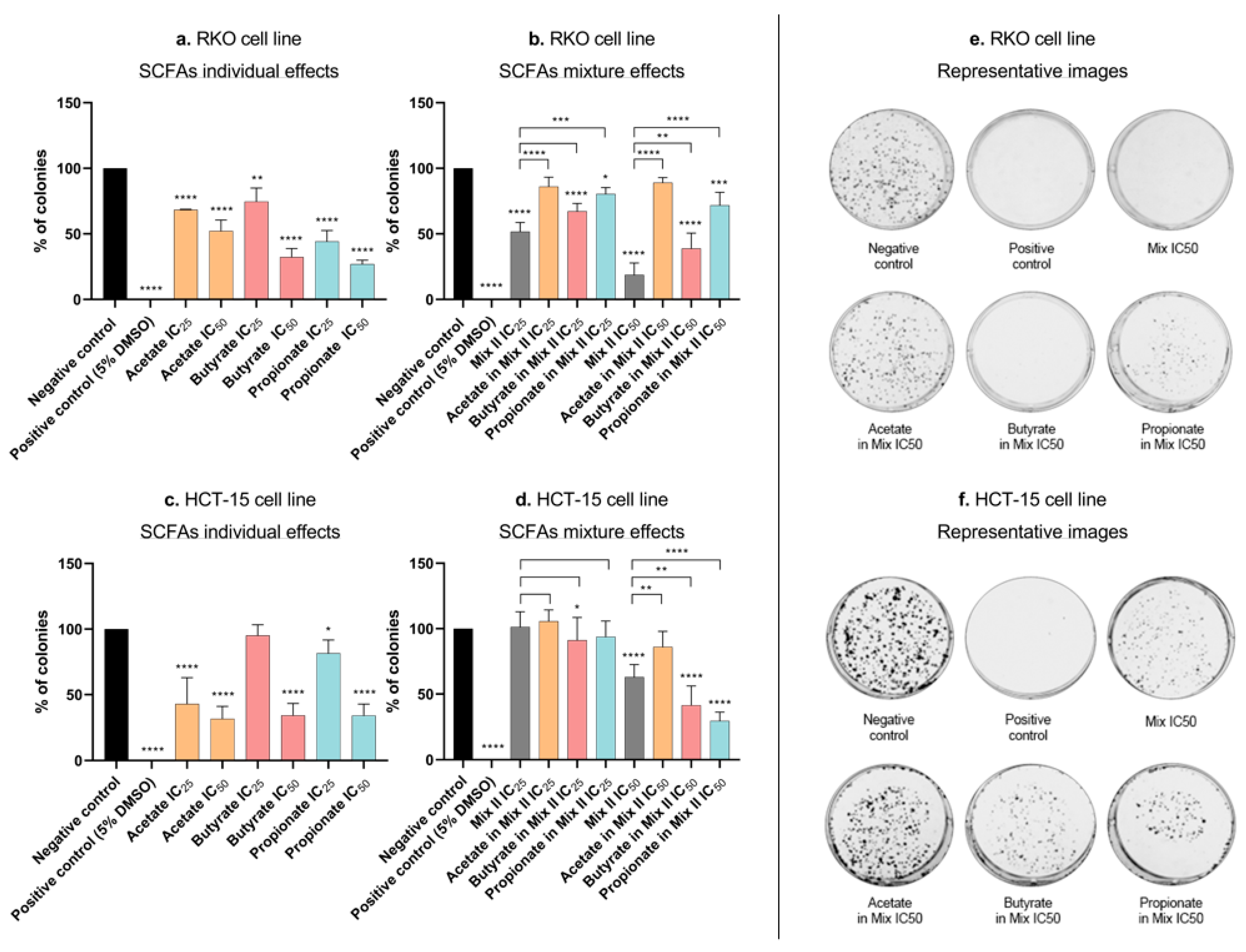
3.3. Short-Chain Fatty Acids Inhibit Colony Formation and Proliferation in Colorectal Cancer Cells
3.4. Short-Chain Fatty Acids Induce Apoptosis in CRC Cells
3.5. Short-Chain Fatty Acids Increase Lactate Production in CRC Cells
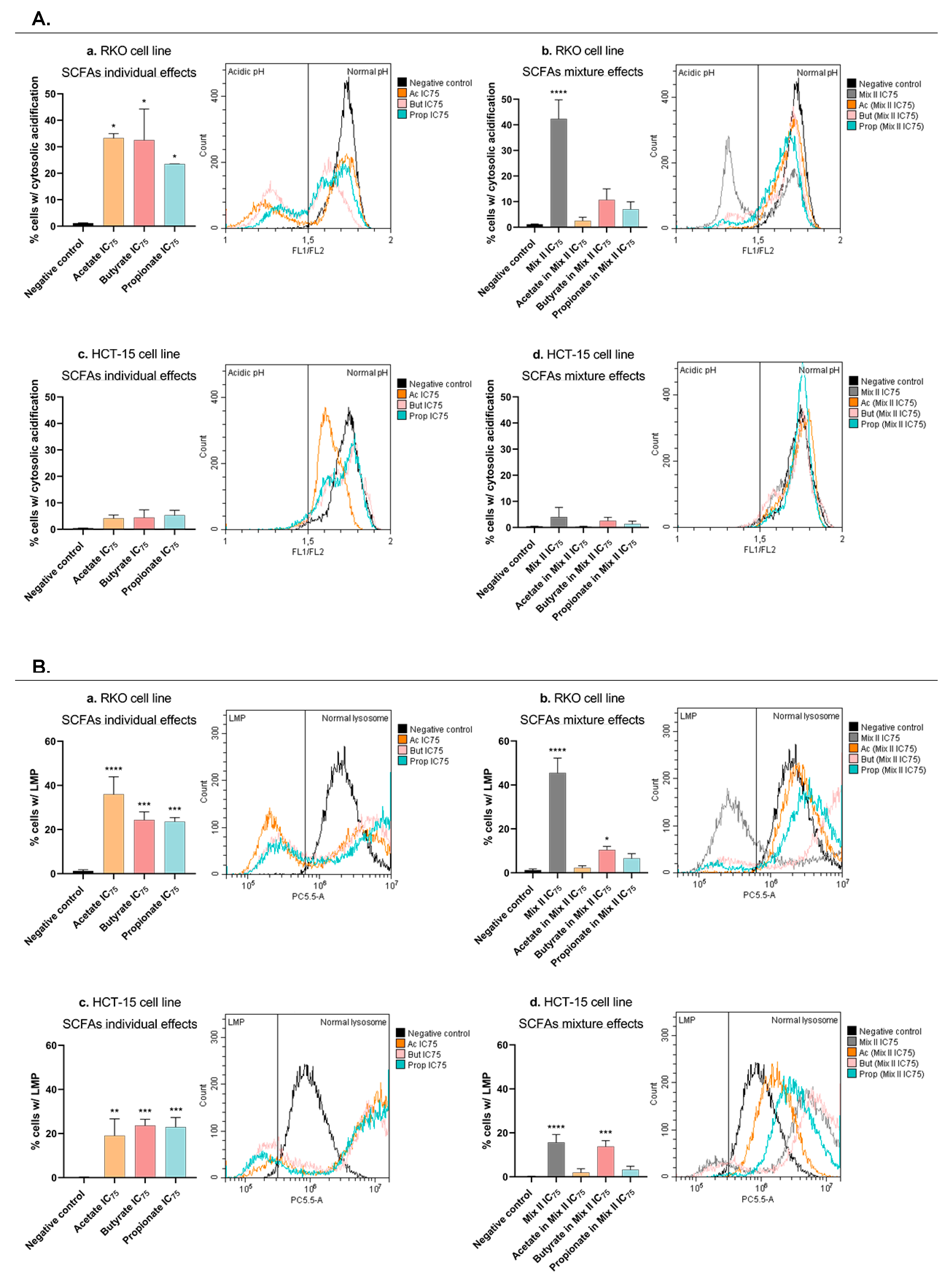
3.6. Short-Chain Fatty Acids Promote Lysosomal-Membrane Permeabilization and Induce Cytosolic Acidification in CRC Cells
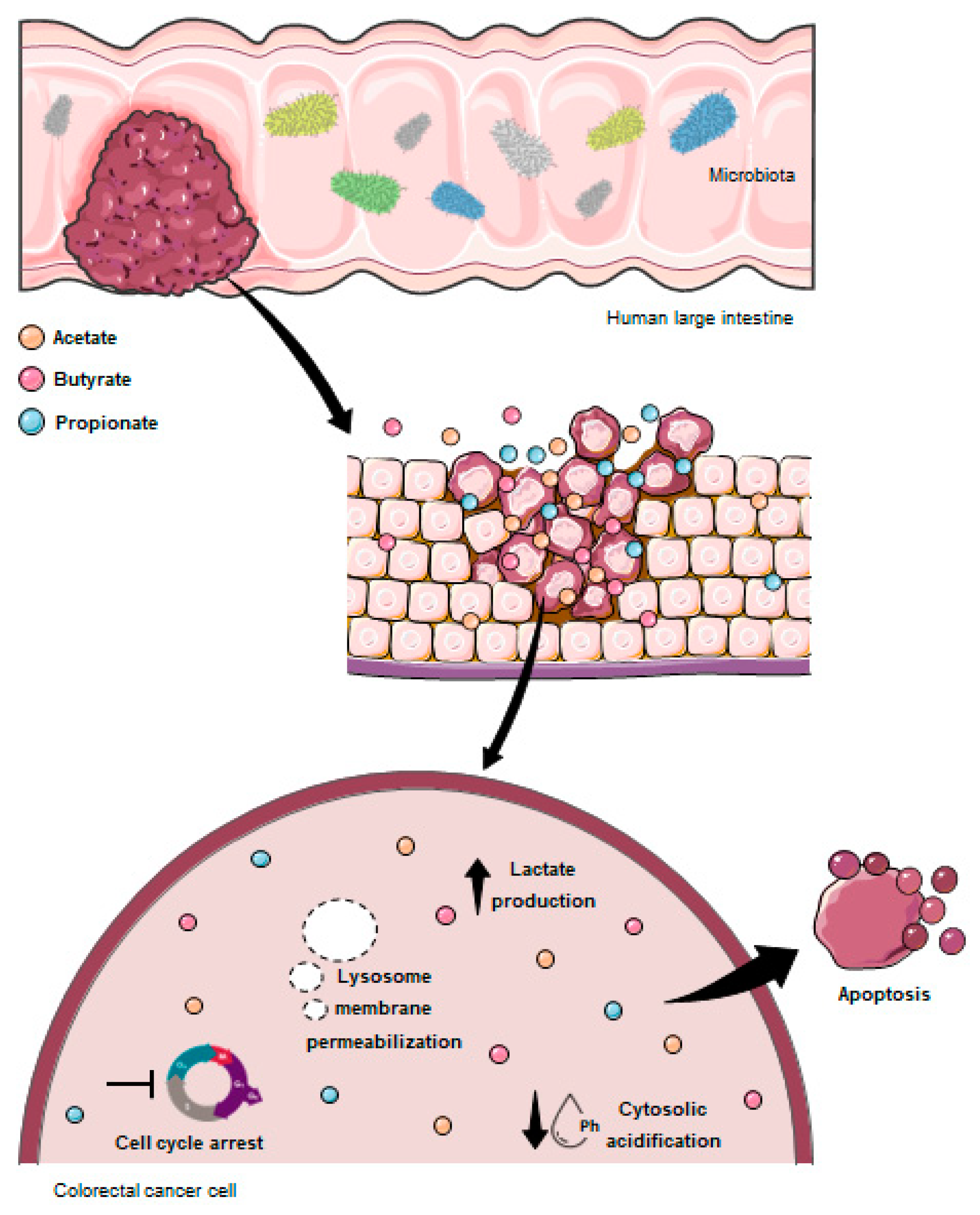
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Observatory, G.C. GLOBOCAN. Available online: https://gco.iarc.fr/ (accessed on 22 July 2022).
- Key, T.J.; Schatzkin, A.; Willett, W.C.; Allen, N.E.; Spencer, E.A.; Travis, R.C. Diet, nutrition and the prevention of cancer. Public Health Nutr. 2004, 7, 187–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pufulete, M. Intake of dairy products and risk of colorectal neoplasia. Nutr. Res. Rev. 2008, 21, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Huncharek, M.; Muscat, J.; Kupelnick, B. Colorectal cancer risk and dietary intake of calcium, vitamin D, and dairy products: A meta-analysis of 26,335 cases from 60 observational studies. Nutr. Cancer 2009, 61, 47–69. [Google Scholar] [CrossRef] [PubMed]
- Veiga, P.; Pons, N.; Agrawal, A.; Oozeer, R.; Guyonnet, D.; Brazeilles, R.; Faurie, J.-M.; van Hylckama Vlieg, J.E.T.; Houghton, L.A.; Whorwell, P.J.; et al. Changes of the human gut microbiome induced by a fermented milk product. Sci. Rep. 2014, 4, 6328. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Kasubuchi, M.; Hasegawa, S.; Hiramatsu, T.; Ichimura, A.; Kimura, I. Dietary gut microbial metabolites, short-chain fatty acids, and host metabolic regulation. Nutrients 2015, 7, 2839–2849. [Google Scholar] [CrossRef] [Green Version]
- D’Argenio, G.; Mazzacca, G. Short-chain fatty acid in the human colon. Relation to inflammatory bowel diseases and colon cancer. Adv. Exp. Med. Biol. 1999, 472, 149–158. [Google Scholar] [CrossRef]
- Yusuf, F.; Adewiah, S.; Syam, A.F.; Fatchiyah, F. Altered profile of gut microbiota and the level short chain fatty acids in colorectal cancer patients. J. Phys. Conf. Ser. 2019, 1146, 12037. [Google Scholar] [CrossRef]
- Ohigashi, S.; Sudo, K.; Kobayashi, D.; Takahashi, O.; Takahashi, T.; Asahara, T.; Nomoto, K.; Onodera, H. Changes of the intestinal microbiota, short chain fatty acids, and fecal pH in patients with colorectal cancer. Dig. Dis. Sci. 2013, 58, 1717–1726. [Google Scholar] [CrossRef]
- Sobhani, I.; Tap, J.; Roudot-Thoraval, F.; Roperch, J.P.; Letulle, S.; Langella, P.; Corthier, G.; Van Nhieu, J.T.; Furet, J.P. Microbial Dysbiosis in Colorectal Cancer (CRC) Patients. PLoS ONE 2011, 6, e16393. [Google Scholar] [CrossRef] [Green Version]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Ding, C.; Meng, T.; Lu, W.; Liu, W.; Hao, H.; Cao, L. Butyrate suppresses motility of colorectal cancer cells via deactivating Akt/ERK signaling in histone deacetylase dependent manner. J. Pharmacol. Sci. 2017, 135, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Queirós, O.; Preto, A.; Pacheco, A.; Pinheiro, C.; Azevedo-Silva, J.; Moreira, R.; Pedro, M.; Ko, Y.H.; Pedersen, P.L.; Baltazar, F.; et al. Butyrate activates the monocarboxylate transporter MCT4 expression in breast cancer cells and enhances the antitumor activity of 3-bromopyruvate. J. Bioenerg. Biomembr. 2012, 44, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Donohoe, D.R.; Collins, L.B.; Wali, A.; Bigler, R.; Sun, W.; Bultman, S.J. The Warburg effect dictates the mechanism of butyrate-mediated histone acetylation and cell proliferation. Mol. Cell 2012, 48, 612–626. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhou, L.; Bao, Y.L.; Wu, Y.; Yu, C.L.; Huang, Y.X.; Sun, Y.; Zheng, L.H.; Li, Y.X. Butyrate induces cell apoptosis through activation of JNK MAP kinase pathway in human colon cancer RKO cells. Chem. Biol. Interact. 2010, 185, 174–181. [Google Scholar] [CrossRef]
- Fung, K.Y.C.; Cosgrove, L.; Lockett, T.; Head, R.; Topping, D.L. A review of the potential mechanisms for the lowering of colorectal oncogenesis by butyrate. Br. J. Nutr. 2012, 108, 820–831. [Google Scholar] [CrossRef] [Green Version]
- Gomes, S.D.; Oliveira, C.S.; Azevedo-Silva, J.; Casanova, M.R.; Barreto, J.; Pereira, H.; Chaves, S.R.; Rodrigues, L.R.; Casal, M.; Côrte-Real, M.; et al. The Role of Diet Related Short-Chain Fatty Acids in Colorectal Cancer Metabolism and Survival: Prevention and Therapeutic Implications. Curr. Med. Chem. 2020, 27, 4087–4108. [Google Scholar] [CrossRef]
- Tedelind, S.; Westberg, F.; Kjerrulf, M.; Vidal, A. Anti-inflammatory properties of the short-chain fatty acids acetate and propionate: A study with relevance to inflammatory bowel disease. World J. Gastroenterol. 2007, 13, 2826–2832. [Google Scholar] [CrossRef]
- Hague, A.; Elder, D.J.; Hicks, D.J.; Paraskeva, C. Apoptosis in colorectal tumour cells: Induction by the short chain fatty acids butyrate, propionate and acetate and by the bile salt deoxycholate. Int. J. Cancer 1995, 60, 400–406. [Google Scholar] [CrossRef]
- Ferro, S.; Azevedo-Silva, J.; Casal, M.; Côrte-Real, M.; Baltazar, F.; Preto, A. Characterization of acetate transport in colorectal cancer cells and potential therapeutic implications. Oncotarget 2016, 7, 70639–70653. [Google Scholar] [CrossRef]
- Oliveira, C.S.F.; Pereira, H.; Alves, S.; Castro, L.; Baltazar, F.; Chaves, S.R.; Preto, A.; Côrte-Real, M. Cathepsin D protects colorectal cancer cells from acetate-induced apoptosis through autophagy-independent degradation of damaged mitochondria. Cell Death Dis. 2015, 6, e1788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, C.; Oliveira, C.S.F.; Alves, S.; Chaves, S.R.; Coutinho, O.P.; Côrte-Real, M.; Preto, A. Acetate-induced apoptosis in colorectal carcinoma cells involves lysosomal membrane permeabilization and cathepsin D release. Cell Death Dis. 2013, 4, e507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiefer, J.; Beyer-Sehlmeyer, G.; Pool-Zobel, B.L. Mixtures of SCFA, composed according to physiologically available concentrations in the gut lumen, modulate histone acetylation in human HT29 colon cancer cells. Br. J. Nutr. 2006, 96, 803–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.; Chen, Y.; Jiang, H.; Nie, D. The role of short-chain fatty acids in orchestrating two types of programmed cell death in colon cancer. Autophagy 2011, 7, 235–237. [Google Scholar] [CrossRef] [Green Version]
- Silva, E.; Rajapakse, N.; Kortenkamp, A. Something from “Nothing”—Eight Weak Estrogenic Chemicals Combined at Concentrations below NOECs Produce Significant Mixture Effects. Environ. Sci. Technol. 2002, 36, 1751–1756. [Google Scholar] [CrossRef]
- Pavlaki, M.D.; Pereira, R.; Loureiro, S.; Soares, A.M.V.M. Effects of binary mixtures on the life traits of Daphnia magna. Ecotoxicol. Environ. Saf. 2011, 74, 99–110. [Google Scholar] [CrossRef]
- Dias da Silva, D.; Carmo, H.; Silva, E. The risky cocktail: What combination effects can we expect between ecstasy and other amphetamines? Arch. Toxicol. 2013, 87, 111–122. [Google Scholar] [CrossRef]
- Silva, E.; Rajapakse, N.; Scholze, M.; Backhaus, T.; Ermler, S.; Kortenkamp, A. Joint effects of heterogeneous estrogenic chemicals in the E-screen--exploring the applicability of concentration addition. Toxicol. Sci. 2011, 122, 383–394. [Google Scholar] [CrossRef] [Green Version]
- Loewe, S.; Muischnek, H. Effect of combinations: Mathematical basis of problem. Arch. Exp. Pathol. Pharmakol. 1926, 114, 313–326. [Google Scholar] [CrossRef]
- Payne, J.; Rajapakse, N.; Wilkins, M.; Kortenkamp, A. Prediction and assessment of the effects of mixtures of four xenoestrogens. Environ. Health Perspect. 2000, 108, 983–987. [Google Scholar] [CrossRef]
- Moyer, M.P.; Manzano, L.A.; Merriman, R.L.; Stauffer, J.S.; Tanzer, L.R. NCM460, a normal human colon mucosal epithelial cell line. In Vitro Cell. Dev. Biol. Anim. 1996, 32, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Brattain, M.G.; Levine, A.E.; Chakrabarty, S.; Yeoman, L.C.; Willson, J.K.V.; Long, B. Heterogeneity of human colon carcinoma. Cancer Metastasis Rev. 1984, 3, 177–191. [Google Scholar] [CrossRef]
- Preto, A.; Figueiredo, J.; Velho, S.; Ribeiro, A.S.; Soares, P.; Oliveira, C.; Seruca, R. BRAF provides proliferation and survival signals in MSI colorectal carcinoma cells displaying BRAF(V600E) but not KRAS mutations. J. Pathol. 2008, 214, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Dexter, D.L.; Barbosa, J.A.; Calabresi, P. N,N-dimethylformamide-induced alteration of cell culture characteristics and loss of tumorigenicity in cultured human colon carcinoma cells. Cancer Res. 1979, 39, 1020–1025. [Google Scholar] [PubMed]
- Tibbetts, L.M.; Chu, M.Y.; Hager, J.C.; Dexter, D.L.; Calabresi, P. Chemotherapy of cell-line-derived human colon carcinomas in mice immunosuppressed with antithymocyte serum. Cancer 1977, 40, 2651–2659. [Google Scholar] [CrossRef]
- Vichai, V.; Kirtikara, K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat. Protoc. 2006, 1, 1112–1116. [Google Scholar] [CrossRef] [PubMed]
- Altenburger, R.; Backhaus, T.; Boedeker, W.; Faust, M.; Scholze, M.; Grimme, L.H. Predictability of the toxicity of multiple chemical mixtures to Vibrio fischeri: Mixtures composed of similarly acting chemicals. Environ. Toxicol. Chem. 2000, 19, 2341–2347. [Google Scholar] [CrossRef]
- Backhaus, T.; Altenburger, R.; Boedeker, W.; Faust, M.; Scholze, M.; Grimme, L.H. Predictability of the toxicity of a multiple mixture of dissimilarly acting chemicals to Vibrio fischeri. Environ. Toxicol. Chem. 2000, 19, 2348–2356. [Google Scholar] [CrossRef]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [Green Version]
- Scholze, M.; Boedeker, W.; Faust, M.; Backhaus, T.; Altenburger, R.; Grimme, L.H. A general best-fit method for concentration-response curves and the estimation of low-effect concentrations. Environ. Toxicol. Chem. 2001, 20, 448–457. [Google Scholar] [CrossRef]
- Ferreira, M.; Assunção, L.S.; Silva, A.H.; Filippin-Monteiro, F.B.; Creczynski-Pasa, T.B.; Sá, M.M. Allylic isothiouronium salts: The discovery of a novel class of thiourea analogues with antitumor activity. Eur. J. Med. Chem. 2017, 129, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Hammond, W.A.; Swaika, A.; Mody, K. Pharmacologic resistance in colorectal cancer: A review. Ther. Adv. Med. Oncol. 2016, 8, 57–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Yao, M.; Lv, L.; Ling, Z.; Li, L. The Human Microbiota in Health and Disease. Engineering 2017, 3, 71–82. [Google Scholar] [CrossRef]
- Teixeira-Guedes, C.; Sánchez-Moya, T.; Pereira-Wilson, C.; Ros-Berruezo, G.; López-Nicolás, R. In Vitro Modulation of Gut Microbiota and Metabolism by Cooked Cowpea and Black Bean. Foods 2020, 9, 861. [Google Scholar] [CrossRef] [PubMed]
- Casanova, M.R.; Azevedo-Silva, J.; Rodrigues, L.R.; Preto, A. Colorectal Cancer Cells Increase the Production of Short Chain Fatty Acids by Propionibacterium freudenreichii Impacting on Cancer Cells Survival. Front. Nutr. 2018, 5, 44. [Google Scholar] [CrossRef]
- Tang, T.; Yang, Z.; Wang, D.; Yang, X.; Wang, J.; Li, L.; Wen, Q.; Gao, L.; Bian, X.; Yu, S. The role of lysosomes in cancer development and progression. Cell Biosci. 2020, 10, 131. [Google Scholar] [CrossRef]
- Ballabio, A. The awesome lysosome. EMBO Mol. Med. 2016, 8, 73–76. [Google Scholar] [CrossRef]
- Liberti, M.V.; Locasale, J.W. The Warburg Effect: How Does it Benefit Cancer Cells? Trends Biochem. Sci. 2016, 41, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Kallunki, T.; Olsen, O.D.; Jäättelä, M. Cancer-associated lysosomal changes: Friends or foes? Oncogene 2013, 32, 1995–2004. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Han, X.; Xu, J. Lysosome as the Black Hole for Checkpoint Molecules. Adv. Exp. Med. Biol. 2020, 1248, 325–346. [Google Scholar] [CrossRef]
- Nogal, A.; Valdes, A.M.; Menni, C. The role of short-chain fatty acids in the interplay between gut microbiota and diet in cardio-metabolic health. Gut Microbes 2021, 13, 1–24. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Compound | Estimated Parameters for the Best-Fit Regression Model of Each Individual Agent | Relative Proportion (%) | |||||
|---|---|---|---|---|---|---|---|---|
| Regression Model | θ1 | θ2 | θmin | θmax | Mix I | Mix II | ||
| RKO | Acetate | Weibull | −6.95 × 100 | 2.32 × 101 | 0.00 | 1.00 | 65.29 | 60.00 |
| Butyrate | Weibull | −1.21 × 100 | 5.12 × 100 | 0.00 | 1.00 | 8.73 | 15.00 | |
| Propionate | Weibull | −3.05 × 100 | 1.33 × 101 | 0.00 | 1.00 | 25.98 | 25.00 | |
| HCT-15 | Acetate | Logit | −4.21 × 100 | 1.21 × 101 | 0.00 | 1.00 | 76.65 | 60.00 |
| Butyrate | Weibull | 8.70 × 102 | 4.05 × 100 | 0.00 | 1.00 | 3.91 | 15.00 | |
| Propionate | Weibull | −1.13 × 100 | 7.18 × 101 | 0.00 | 1.00 | 19.44 | 25.00 | |
| NCM460 | Acetate | Weibull | −8.05 × 100 | 2.40 × 101 | 0.00 | 1.00 | 60.31 | 60.00 |
| Butyrate | Weibull | −1.85 × 100 | 6.51 × 100 | 0.00 | 1.00 | 13.35 | 15.00 | |
| Propionate | Logit | −4.00 × 100 | 1.26 × 101 | 0.00 | 1.00 | 26.33 | 25.00 | |
| Compound | IC50 (mM) | Selectivity Index | |||
|---|---|---|---|---|---|
| RKO | HCT-15 | NCM460 | RKO | HCT-15 | |
| Acetate | 81.04 ± 2.24 | 89.52 ± 3.31 | 117.30 ± 2.99 | 1.45 | 1.31 |
| Butyrate | 10.84 ± 0.79 | 4.57 ± 0.30 | 25.96 ± 1.80 | 2.39 | 5.68 |
| Propionate | 32.25 ± 1.14 | 22.70 ± 1.59 | 51.21 ± 3.42 | 1.59 | 2.26 |
| Cell lines | Dose | Mixture II (mM) | Acetate in Mixture II (mM) | Butyrate in Mixture II (mM) | Propionate in Mixture II (mM) |
|---|---|---|---|---|---|
| RKO | IC75 | 77.32 | 46.39 | 11.60 | 19.33 |
| IC50 | 47.61 | 28.57 | 7.14 | 11.90 | |
| IC25 | 28.51 | 11.17 | 4.28 | 7.13 | |
| HCT-15 | IC75 | 39.97 | 23.98 | 6.00 | 9.99 |
| IC50 | 13.67 | 8.20 | 2.05 | 3.42 | |
| IC25 | 4.79 | 2.87 | 0.72 | 1.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, S.; Baltazar, F.; Silva, E.; Preto, A. Microbiota-Derived Short-Chain Fatty Acids: New Road in Colorectal Cancer Therapy. Pharmaceutics 2022, 14, 2359. https://doi.org/10.3390/pharmaceutics14112359
Gomes S, Baltazar F, Silva E, Preto A. Microbiota-Derived Short-Chain Fatty Acids: New Road in Colorectal Cancer Therapy. Pharmaceutics. 2022; 14(11):2359. https://doi.org/10.3390/pharmaceutics14112359
Chicago/Turabian StyleGomes, Sara, Fátima Baltazar, Elisabete Silva, and Ana Preto. 2022. "Microbiota-Derived Short-Chain Fatty Acids: New Road in Colorectal Cancer Therapy" Pharmaceutics 14, no. 11: 2359. https://doi.org/10.3390/pharmaceutics14112359
APA StyleGomes, S., Baltazar, F., Silva, E., & Preto, A. (2022). Microbiota-Derived Short-Chain Fatty Acids: New Road in Colorectal Cancer Therapy. Pharmaceutics, 14(11), 2359. https://doi.org/10.3390/pharmaceutics14112359