
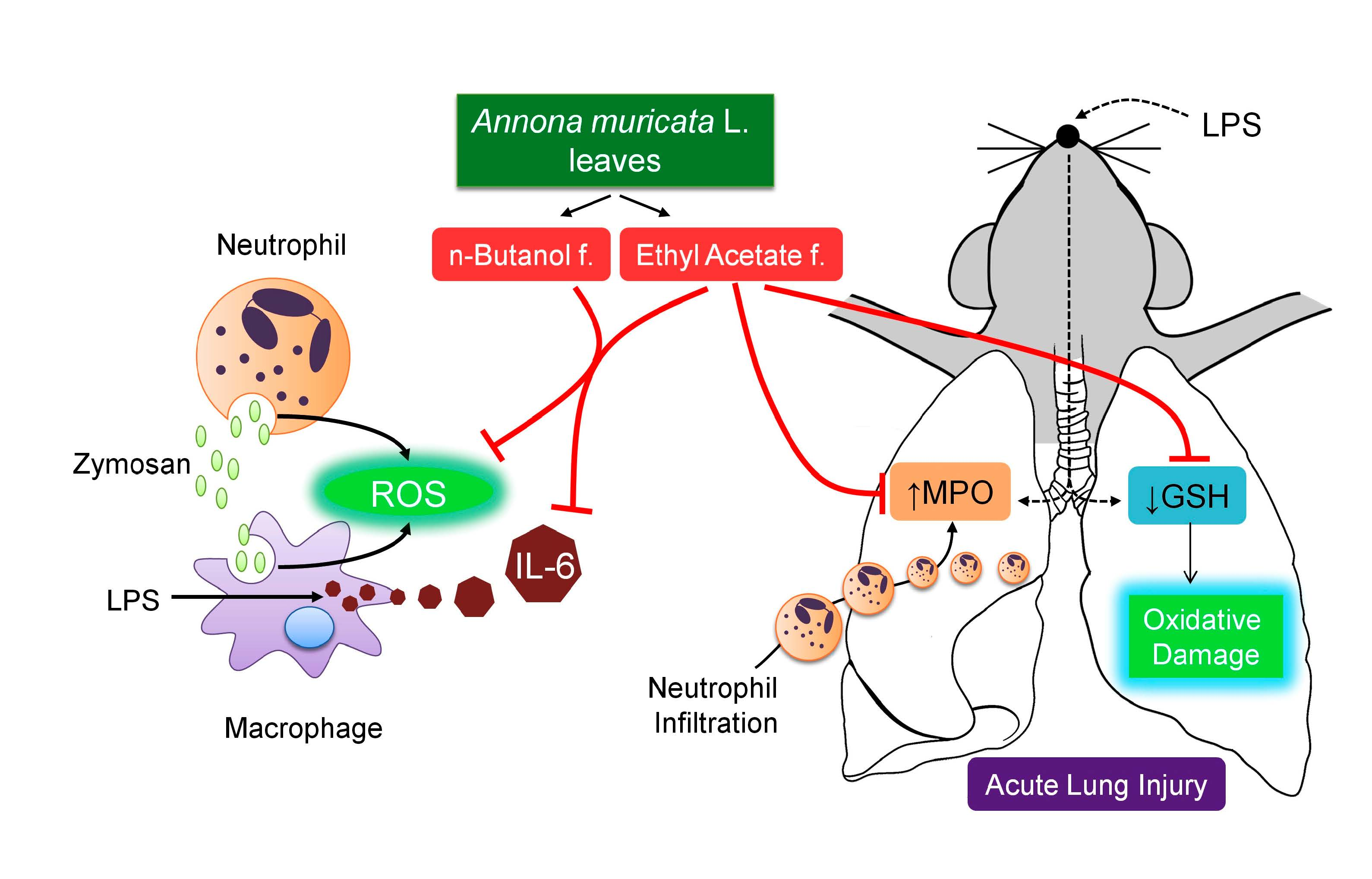
Polyphenols-Rich Fraction from Annona muricata Linn. Leaves Attenuates Oxidative and Inflammatory Responses in Neutrophils, Macrophages, and Experimental Lung Injury

 , ,
, ,  ,
, 
Abstract
:
1. Introduction
2. Material and Methods
2.1. Reagents
2.2. Animals
2.3. Plant Extracts and Fractions
2.4. HPLC-ESI/MSn Characterization of Phytoconstituents in EtOAc.f
2.5. Primary Cell Culture
2.6. ROS Production by Neutrophils and Macrophages
2.7. Cell Viability of Neutrophils
2.8. BMDMs Phagocytosis Analysis
2.9. Evaluation of Cytokine Secretion, Co-Stimulatory Molecules Expression, and Cell Viability Assay in BMDMs
2.10. Acute Lung Injury (ALI) Induction
2.11. Cytokine Levels and Cell Count in Bronchoalveolar Lavage Fluid (BALF)
2.12. Myeloperoxidase (MPO) Activity
2.13. Tissue Processing and Protein Dosage
2.14. Determination of Superoxide Dismutase (SOD) and Catalase (CAT) Activities, GSH, and Protein Carbonylation Content
2.15. Histopathological Analysis
2.16. In Vivo Toxicity Analysis of EtOAc.f
2.17. Statistical Analysis
3. Results
3.1. Inhibitory Effects of EtOAc.f and BuOH.f from A. muricata Leaves on Neutrophil-Derived ROS
3.2. EtOAc and BuOH Fractions Reduce BMDMs ROS Generation
3.3. EtOAc and BuOH Fractions Modulate Inflammatory Functions of BMDMs
3.4. EtOAc Fraction Reduces MPO Activity and Recovers Oxidative Damage in the Lung during Experimental LPS-Induced ALI
3.5. EtOAc.f from A. muricata Leaves Did Not Produce Toxic Effects In Vivo
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Pinto, A.C.Q. Origin and Distribution; Willians, J.T., Smith, R.W., Hughes, A., Haq, N., Clement, C.R., Eds.; Annona Species; International Centre for Underutilised Crops: Southampton, UK, 2005; pp. 17–20. [Google Scholar]
- Mootoosamy, A.; Mahomoodally, M.F. Ethnomedicinal application of native remedies used against diabetes and related complications in Mauritius. J. Ethnopharmacol. 2014, 151, 413–444. [Google Scholar] [CrossRef] [PubMed]
- Bussmann, R.; Garcia, G.R.M.; Glenn, A.; Sharon, D.; Chait, G.; Díaz, D.; Pourmand, K.; Jonat, B.; Somogy, S.; Guardado, G.; et al. Minimum inhibitory concentrations of medicinal plants used in Northern Peru as antibacterial remedies. J. Ethnopharmacol. 2010, 132, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandebroek, I.; Balick, M.J.; Ososki, A.; Kronenberg, F.; Yukes, J.; Wade, C.; Jiménez, F.; Peguero, B.; Castillo, D. The importance of botellas and other plant mixtures in Dominican traditional medicine. J. Ethnopharmacol. 2010, 128, 20–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cercato, L.M.; White, P.A.; Nampo, F.K.; Santos, M.R.; Camargo, E.A. A systematic review of medicinal plants used for weight loss in Brazil: Is there potential for obesity treatment? J. Ethnopharmacol. 2015, 176, 286–296. [Google Scholar] [CrossRef]
- Ritter, R.A.; Monteiro, M.V.B.; Monteiro, F.; Rodrigues, S.T.; Soares, M.L.; Silva, J.C.R.; Palha, M.D.D.C.; Biondi, G.F.; Rahal, S.; Tourinho, M.M. Ethnoveterinary knowledge and practices at Colares island, Pará state, eastern Amazon, Brazil. J. Ethnopharmacol. 2012, 144, 346–352. [Google Scholar] [CrossRef] [Green Version]
- Gyesi, J.N.; Opoku, R.; Borquaye, L.S. Chemical Composition, Total Phenolic Content, and Antioxidant Activities of the Essential Oils of the Leaves and Fruit Pulp of Annona muricata L. (Soursop) from Ghana. Biochem. Res. Int. 2019, 2019, 4164576. [Google Scholar] [CrossRef] [Green Version]
- George, V.C.; Kumar, D.R.N.; Rajkumar, V.; Suresh, P.K.; Kumar, R.A. Antiviral, Antioxidant, and Antihemolytic Effect of Annona muricata L. Leaves Extracts. Plants 2020, 9, 1650. [Google Scholar] [CrossRef]
- Chan, P.; Ah, R.; Mh, K. Anti-arthritic activities of Annona muricata L. leaves extract on complete Freund’s adjuvant (CFA)—Induced arthritis in rats. Planta Med. 2010, 76, 166. [Google Scholar] [CrossRef]
- Quilez, A.; La Paz, S.M.-D.; De La Puerta, R.; Fernandez-Arche, M.; Gimenez, G. Validation of ethnopharmacological use as anti-inflammatory of a decoction from Annona muricata leaves. Afr. J. Tradit. Complement. Altern. Med. 2015, 12, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Galeas-Pena, M.; McLaughlin, N.; Pociask, D. The role of the innate immune system on pulmonary infections. Biol. Chem. 2019, 400, 443–456. [Google Scholar] [CrossRef]
- Matthay, M.A.; Zemans, R.L.; Zimmerman, G.A.; Arabi, Y.; Beitler, J.R.; Mercat, A.; Herridge, M.; Randolph, A.G.; Calfee, C.S. Acute respiratory distress syndrome. Nat. Rev. Dis. Prim. 2019, 5, 18. [Google Scholar] [CrossRef] [PubMed]
- Thompson, B.T.; Chambers, R.; Liu, K. Acute Respiratory Distress Syndrome. N. Engl. J. Med. 2017, 377, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.J.; Roy, S.B.; Mehta, H.J.; Joo, M.; Sadikot, R.T. Alternative and Natural Therapies for Acute Lung Injury and Acute Respiratory Distress Syndrome. BioMed Res. Int. 2018, 2018, 2476824. [Google Scholar] [CrossRef] [Green Version]
- Justino, A.; Miranda, N.C.; Franco, R.R.; Martins, M.M.; da Silva, N.M.; Espindola, F.S. Annona muricata Linn. leaf as a source of antioxidant compounds with in vitro antidiabetic and inhibitory potential against α-amylase, α-glucosidase, lipase, non-enzymatic glycation and lipid peroxidation. Biomed. Pharmacother. 2018, 100, 83–92. [Google Scholar] [CrossRef]
- Allen, R.C. Phagocytic leukocyte oxygenation activities and chemiluminescence: A kinetic approach to analysis. Methods Enzymol. 1986, 133, 449–493. [Google Scholar] [CrossRef]
- Nuutila, J.; Lilius, E.-M. Flow cytometric quantitative determination of ingestion by phagocytes needs the distinguishing of overlapping populations of binding and ingesting cells. Cytom. Part A 2005, 65, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Khadangi, F.; Forgues, A.-S.; Tremblay-Pitre, S.; Dufour-Mailhot, A.; Henry, C.; Boucher, M.; Beaulieu, M.-J.; Morissette, M.; Fereydoonzad, L.; Brunet, D.; et al. Intranasal versus intratracheal exposure to lipopolysaccharides in a murine model of acute respiratory distress syndrome. Sci. Rep. 2021, 11, 7777. [Google Scholar] [CrossRef] [PubMed]
- Szarka, R.J.; Wang, N.; Gordon, L.; Nation, P.; Smith, R.H. A murine model of pulmonary damage induced by lipopolysaccharide via intranasal instillation. J. Immunol. Methods 1997, 202, 49–57. [Google Scholar] [CrossRef]
- Rubin, B.B.; Smith, A.; Liauw, S.; Isenman, D.; Romaschin, A.D.; Walker, P.M. Complement activation and white cell sequestration in postischemic skeletal muscle. Am. J. Physiol. Circ. Physiol. 1990, 259, H525–H531. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Li, X. Improved Pyrogallol Autoxidation Method: A Reliable and Cheap Superoxide-Scavenging Assay Suitable for All Antioxidants. J. Agric. Food Chem. 2012, 60, 6418–6424. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Browne, R.W.; Armstrong, D. Reduced Glutathione and Glutathione Disulfide. Methods Mol. Biol. 1998, 108, 347–352. [Google Scholar] [CrossRef]
- Reznick, A.Z.; Packer, L. Oxidative damage to proteins: Spectrophotometric method for carbonyl assay. Methods Enzymol. 1994, 233, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.; Tsoni, S.V.; Willment, J.; Dennehy, K.M.; Rosas, M.; Findon, H.; Haynes, K.; Steele, C.; Botto, M.; Gordon, S.; et al. Dectin-1 is required for β-glucan recognition and control of fungal infection. Nat. Immunol. 2007, 8, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Holmström, K.M.; Finkel, T. Cellular mechanisms and physiological consequences of redox-dependent signalling. Nat. Rev. Mol. Cell Biol. 2014, 15, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Moldogazieva, N.T.; Mokhosoev, I.M.; Feldman, N.B.; Lutsenko, S.V. ROS and RNS signalling: Adaptive redox switches through oxidative/nitrosative protein modifications. Free Radic. Res. 2018, 52, 507–543. [Google Scholar] [CrossRef] [PubMed]
- Ward, P.A. Oxidative stress: Acute and progressive lung injury. Ann. N. Y. Acad. Sci. 2010, 1203, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Ye, W.; Zheng, C.; Yu, D.; Zhang, F.; Pan, R.; Ni, X.; Shi, Z.; Zhang, Z.; Xiang, Y.; Sun, H.; et al. Lipoxin A4 Ameliorates Acute Pancreatitis-Associated Acute Lung Injury through the Antioxidative and Anti-Inflammatory Effects of the Nrf2 Pathway. Oxid. Med. Cell. Longev. 2019, 2019, 2197017. [Google Scholar] [CrossRef] [Green Version]
- Kellner, M.; Noonepalle, S.; Lu, Q.; Srivastava, A.; Zemskov, E.; Black, S.M. Black, ROS signaling in the pathogenesis of acute lung injury (ALI) and acute respiratory distress syndrome (ARDS). Adv. Exp. Med. Biol. 2017, 967, 105–137. [Google Scholar] [CrossRef]
- George, V.C.; Kumar, D.R.N.; Suresh, P.K.; Kumar, R.A. Antioxidant, DNA protective efficacy and HPLC analysis of Annona muricata (soursop) extracts. J. Food Sci. Technol. 2015, 52, 2328–2335. [Google Scholar] [CrossRef] [PubMed]
- Wahab, S.M.A.; Jantan, I.; Haque, A.; Arshad, L. Exploring the Leaves of Annona muricata L. as a Source of Potential Anti-inflammatory and Anticancer Agents. Front. Pharmacol. 2018, 9, 661. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Li, X.J.; Stull, N.D.; Ming, W.; Suh, C.-I.; Bissonnette, S.A.; Yaffe, M.B.; Grinstein, S.; Atkinson, S.J.; Dinauer, M.C. FcγR-stimulated activation of the NADPH oxidase: Phosphoinositide-binding protein p40phox regulates NADPH oxidase activity after enzyme assembly on the phagosome. Blood 2008, 112, 3867–3877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cachat, J.; Deffert, C.; Hugues, S.; Krause, K.-H. Phagocyte NADPH oxidase and specific immunity. Clin. Sci. 2015, 128, 635–648. [Google Scholar] [CrossRef]
- Carnesecchi, S.; Pache, J.-C.; Barazzone-Argiroffo, C. NOX enzymes: Potential target for the treatment of acute lung injury. Cell. Mol. Life Sci. 2012, 69, 2373–2385. [Google Scholar] [CrossRef]
- Lundqvist, H.; Foilin, P.; Khalfan, L.; Dahlgren, C. Phorbol myristate acetate-induced NADPH oxidase activity in human neutrophils: Only half the story has been told. J. Leukoc. Biol. 1996, 59, 270–279. [Google Scholar] [CrossRef]
- Suttles, J.; Stout, R.D. Macrophage CD40 signaling: A pivotal regulator of disease protection and pathogenesis. Semin. Immunol. 2009, 21, 257–264. [Google Scholar] [CrossRef]
- Jones, S.A.; Jenkins, B.J. Recent insights into targeting the IL-6 cytokine family in inflammatory diseases and cancer. Nat. Rev. Immunol. 2018, 18, 773–789. [Google Scholar] [CrossRef]
- Yasui, M.; Matsushima, M.; Omura, A.; Mori, K.; Ogasawara, N.; Kodera, Y.; Shiga, M.; Ito, K.; Kojima, S.; Kawabe, T. The Suppressive Effect of Quercetin on Toll-Like Receptor 7-Mediated Activation in Alveolar Macrophages. Pharmacology 2015, 96, 201–209. [Google Scholar] [CrossRef]
- Kim, S.-H.; Park, S.-Y.; Park, Y.-L.; Myung, D.-S.; Rew, J.-S.; Joo, Y.-E. Chlorogenic acid suppresses lipopolysaccharide-induced nitric oxide and interleukin-1β expression by inhibiting JAK2/STAT3 activation in RAW264.7 cells. Mol. Med. Rep. 2017, 16, 9224–9232. [Google Scholar] [CrossRef] [Green Version]
- Justino, A.; Pereira, M.N.; Vilela, D.D.; Peixoto, L.; Martins, M.M.; Teixeira, R.R.; Miranda, N.C.; da Silva, N.M.; Sousa, R.; de Oliveira, A.; et al. Peel of araticum fruit (Annona crassiflora Mart.) as a source of antioxidant compounds with α-amylase, α-glucosidase and glycation inhibitory activities. Bioorganic Chem. 2016, 69, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Domscheit, H.; Hegeman, M.A.; Carvalho, N.; Spieth, P.M. Molecular Dynamics of Lipopolysaccharide-Induced Lung Injury in Rodents. Front. Physiol. 2020, 11, 36. [Google Scholar] [CrossRef] [PubMed]
- Cercato, L.M.; Araújo, J.M.D.; Oliveira, A.S.; Melo, A.J.O.; Lima, B.S.; dos Santos, E.W.P.; Neto, A.G.D.S.; de Albuquerque-Júnior, R.L.C.; Duarte, M.C.; Araujo, A.A.S.; et al. Reduced cutaneous inflammation associated with antioxidant action after topical application of the aqueous extract of Annona muricata leaves. Inflammopharmacology 2021, 29, 307–315. [Google Scholar] [CrossRef]
- Schultz, J.; Kaminker, K. Myeloperoxidase of the leucocyte of normal human blood. I. Content and localization. Arch. Biochem. Biophys. 1962, 96, 465–467. [Google Scholar] [CrossRef]
- Daubeuf, F.; Frossard, N. Performing Bronchoalveolar Lavage in the Mouse. Curr. Protoc. Mouse Biol. 2012, 2, 167–175. [Google Scholar] [CrossRef]
- Aggarwal, N.R.; King, L.S.; D’Alessio, F.R. Diverse macrophage populations mediate acute lung inflammation and resolution. Am. J. Physiol. Cell. Mol. Physiol. 2014, 306, L709–L725. [Google Scholar] [CrossRef]
- Laskin, D.L.; Malaviya, R.; Laskin, J.D. Role of Macrophages in Acute Lung Injury and Chronic Fibrosis Induced by Pulmonary Toxicants. Toxicol. Sci. 2019, 168, 287–301. [Google Scholar] [CrossRef] [Green Version]
- Hui, L.; Zhang, X.; An, X.; Li, J.; Zang, K.; Shang, F.; Zhang, C.; Zhang, G. Higher serum procalcitonin and IL-6 levels predict worse diagnosis for acute respiratory distress syndrome patients with multiple organ dysfunction. Int. J. Clin. Exp. Pathol. 2017, 10, 7401–7407. [Google Scholar]
- Malaviya, R.; Laskin, J.D.; Laskin, D.L. Anti-TNFα therapy in inflammatory lung diseases. Pharmacol. Ther. 2017, 180, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Neveu, W.A.; Bernardo, E.; Allard, J.L.; Nagaleekar, V.; Wargo, M.J.; Davis, R.J.; Iwakura, Y.; Whittaker, L.A.; Rincon, M. Fungal Allergen β-Glucans Trigger p38 Mitogen-Activated Protein Kinase–Mediated IL-6 Translation in Lung Epithelial Cells. Am. J. Respir. Cell Mol. Biol. 2011, 45, 1133–1141. [Google Scholar] [CrossRef] [Green Version]
- Khair, O.; Devalia, J.; Abdelaziz, M.; Sapsford, R.; Tarraf, H.; Davies, R. Effect of Haemophilus influenzae endotoxin on the synthesis of IL-6, IL-8, TNF-α and expression of ICAM-1 in cultured human bronchial epithelial cells. Eur. Respir. J. 1994, 7, 2109–2116. [Google Scholar] [CrossRef] [Green Version]
- Rosanna, D.P. Reactive Oxygen Species, Inflammation, and Lung Diseases. Curr. Pharm. Des. 2012, 18, 3889–3900. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, M.; Viken, K.; Wang, Q.; Jagtap, P.; Bitterman, P.; Ingbar, D.H.; Wendt, C. Bronchoalveolar Lavage Fluid Protein Expression in Acute Respiratory Distress Syndrome Provides Insights into Pathways Activated in Subjects with Different Outcomes. Sci. Rep. 2017, 7, 7464. [Google Scholar] [CrossRef] [PubMed]
- Trajano, E.T.L.; Sternberg, C.; Caetano, M.; Silva, M.A.S.; Porto, L.C.; Santos, J.C.; Ribeiro, M.L.; Magalhães, C.B.; Zin, W.A.; Benjamim, C.F.; et al. Endotoxin-induced acute lung injury is dependent upon oxidative response. Inhal. Toxicol. 2011, 23, 918–926. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Dimitropoulou, C.; Lu, Q.; Black, S.M.; Sharma, S. Glutathione Supplementation Attenuates Lipopolysaccharide-Induced Mitochondrial Dysfunction and Apoptosis in a Mouse Model of Acute Lung Injury. Front. Physiol. 2012, 3, 161. [Google Scholar] [CrossRef] [Green Version]
- Adewole, S.O.; Ojewole, J.A.O. Protective effects of Annona muricata linn. (Annonaceae) leaf aqueous extract on serum lipid profiles and oxidative stress in hepatocytes of streptozotocin-treated diabetic rats. Afr. J. Tradit. Complement. Altern. Med. 2009, 6, 30–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsenosy, A.E.-W.; El-Far, A.H.; Sadek, K.M.; Ibrahim, S.A.; Atta, M.S.; Sayed-Ahmed, A.; Al Jaouni, S.K.; Mousa, S.A. Graviola (Annona muricata) attenuates behavioural alterations and testicular oxidative stress induced by streptozotocin in diabetic rats. PLoS ONE 2019, 14, e0222410. [Google Scholar] [CrossRef]
- Zhang, W.-J.; Wei, H.; Tien, Y.-T.; Frei, B. Genetic ablation of phagocytic NADPH oxidase in mice limits TNFα-induced inflammation in the lungs but not other tissues. Free Radic. Biol. Med. 2011, 50, 1517–1525. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-F.; Song, S.-D.; Li, Y.-J.; Hu, Z.Q.; Zhang, Z.-W.; Yan, C.-G.; Li, Z.-G.; Tang, H.-F. Protective Effect of Quercetin in LPS-Induced Murine Acute Lung Injury Mediated by cAMP-Epac Pathway. Inflammation 2018, 41, 1093–1103. [Google Scholar] [CrossRef]
- Xing, J.; Yu, Z.; Zhang, X.; Li, W.; Gao, D.; Wang, J.; Ma, X.; Nie, X.; Wang, W. Epicatechin alleviates inflammation in lipopolysaccharide-induced acute lung injury in mice by inhibiting the p38 MAPK signaling pathway. Int. Immunopharmacol. 2019, 66, 146–153. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction | ROS Generation (Percentage of Control) ± S.E.M. | ||
|---|---|---|---|
| Concentration | |||
| 1 µg/mL | 3 µg/mL | 10 µg/mL | |
| Ethanol (EtOHcr) | 94.09 ± 6.92 | 92.20 ± 5.07 | 50.07 ± 2.17 |
| Hexane (Hex.f) | 140.93 ± 7.43 | 60.77 ± 10.54 | 57.45 ± 10.15 |
| Dichloromethane (DCM.f) | 59.66 ± 15.87 | 96.84 ± 4.24 | 35.33 ± 6.83 |
| Ethyl Acetate (EtOAc.f) | 89.41 ± 14.10 | 39.91 ± 7.23 | 3.11 ± 0.32 |
| n-Butanol (BuOH.f) | 60.67 ± 12.61 | 33.71 ± 6.21 | 3.44 ± 1.16 |
| Water (Wa.f) | 76.62 ± 8.64 | 50.77 ± 11.07 | 82.74 ± 19.83 |
| Histopathology | Sal | Veh/LPS | EtOAc.f/LPS |
|---|---|---|---|
| pseudostratified epithelium | preserved | preserved | preserved |
| simple epithelium | preserved | partially preserved | partially preserved |
| connective tissue | preserved | partially preserved | preserved |
| pulmonary alveolus | preserved | partially preserved | preserved |
| inflammation | absent | accentuated | discreet |
| Veh | EtOAc | |
|---|---|---|
| WBC (×103/μL) | 13.92 ± 2.78 | 13.24 ± 2.11 |
| Neutrophil (%) | 14.6 ± 0.97 | 13.00 ± 0.83 |
| Eosinophils (%) | 4.00 ± 0.83 | 3.60 ± 0.67 |
| Basophils (%) | ND | ND |
| Lymphocytes (%) | 77.6 ± 1.69 | 81.00 ± 1.14 |
| Monocytes (%) | 2.00 ± 0.51 | 2.40 ± 0.81 |
| RBC (×103/μL) | 8.94 ± 0.12 | 8.73 ± 0.14 |
| HCT (%) | 40.64 ± 0.35 | 39.98 ± 0.41 |
| Hb (g/dL) | 14.02 ± 0.14 | 13.62 ± 0.17 |
| MCH (pg) | 15.72 ± 0.08 | 15.56 ± 0.11 |
| PL (×103/μL) | 661.00 ± 160.50 | 775.8 ± 195.90 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saraiva, A.L.; Justino, A.B.; Franco, R.R.; Silva, H.C.G.; Arruda, F.d.S.; Klein, S.G.; Celes, M.R.N.; Goulart, L.R.; Espindola, F.S. Polyphenols-Rich Fraction from Annona muricata Linn. Leaves Attenuates Oxidative and Inflammatory Responses in Neutrophils, Macrophages, and Experimental Lung Injury. Pharmaceutics 2022, 14, 1182. https://doi.org/10.3390/pharmaceutics14061182
Saraiva AL, Justino AB, Franco RR, Silva HCG, Arruda FdS, Klein SG, Celes MRN, Goulart LR, Espindola FS. Polyphenols-Rich Fraction from Annona muricata Linn. Leaves Attenuates Oxidative and Inflammatory Responses in Neutrophils, Macrophages, and Experimental Lung Injury. Pharmaceutics. 2022; 14(6):1182. https://doi.org/10.3390/pharmaceutics14061182
Chicago/Turabian StyleSaraiva, André Lopes, Allisson Benatti Justino, Rodrigo Rodrigues Franco, Heitor Cappato Guerra Silva, Felipe dos Santos Arruda, Sandra Gabriela Klein, Mara Rúbia Nunes Celes, Luiz Ricardo Goulart, and Foued Salmen Espindola. 2022. "Polyphenols-Rich Fraction from Annona muricata Linn. Leaves Attenuates Oxidative and Inflammatory Responses in Neutrophils, Macrophages, and Experimental Lung Injury" Pharmaceutics 14, no. 6: 1182. https://doi.org/10.3390/pharmaceutics14061182
APA StyleSaraiva, A. L., Justino, A. B., Franco, R. R., Silva, H. C. G., Arruda, F. d. S., Klein, S. G., Celes, M. R. N., Goulart, L. R., & Espindola, F. S. (2022). Polyphenols-Rich Fraction from Annona muricata Linn. Leaves Attenuates Oxidative and Inflammatory Responses in Neutrophils, Macrophages, and Experimental Lung Injury. Pharmaceutics, 14(6), 1182. https://doi.org/10.3390/pharmaceutics14061182