
Intranasal Methylprednisolone Ameliorates Neuroinflammation Induced by Chronic Toluene Exposure

 ,
,  , , , and
, , , and 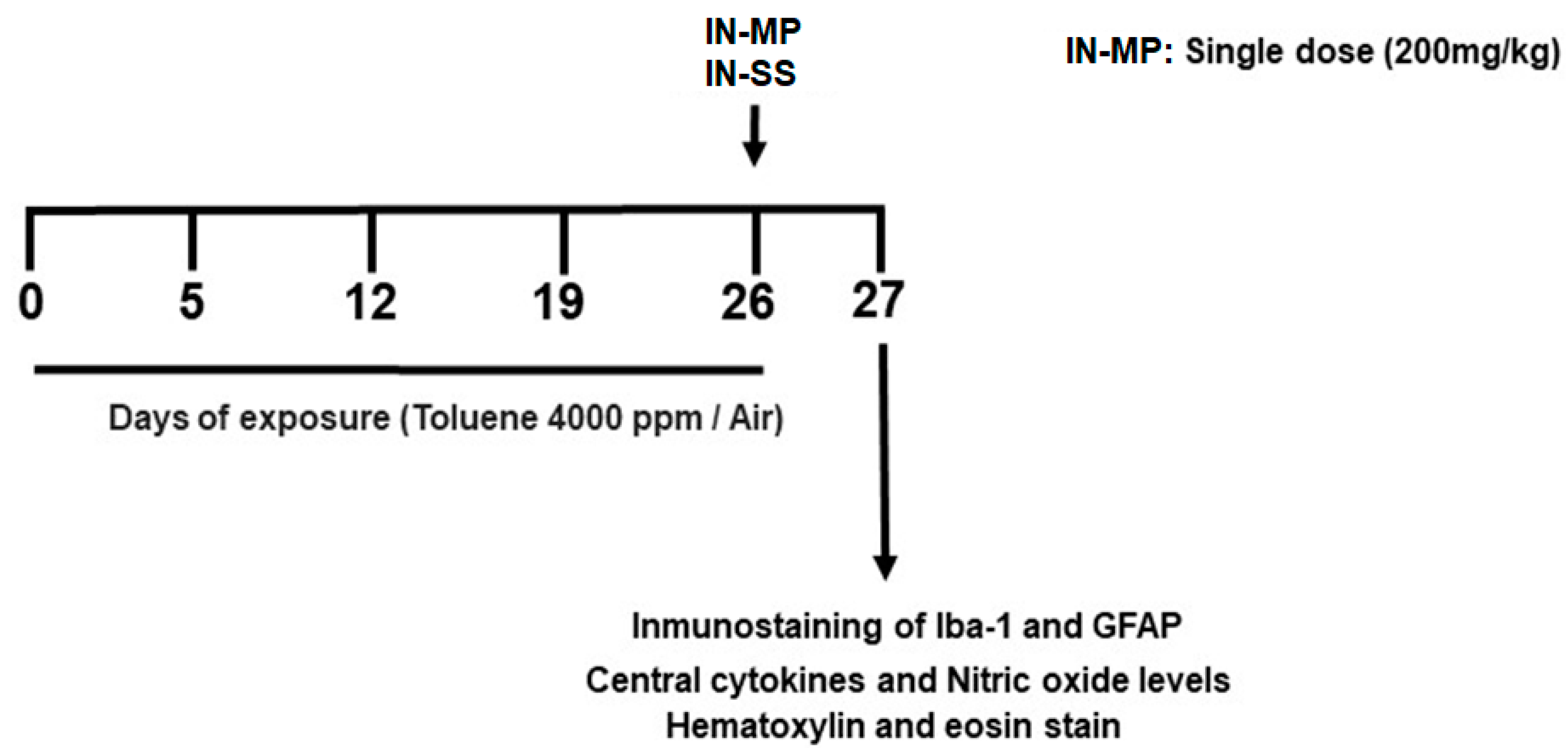{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Chronic Toluene Exposure
2.3. Effect of Intranasal Methylprednisolone Administration
2.4. Central Inflammatory Mediators
2.5. Cytokine Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Nitric Oxide Assay
2.7. Immunofluorescence (IFC) Analysis
2.8. Histological Analysis
2.9. Analysis of Addictive Behavior by Measuring the Locomotor Activity
2.10. Statistical Analysis
3. Results
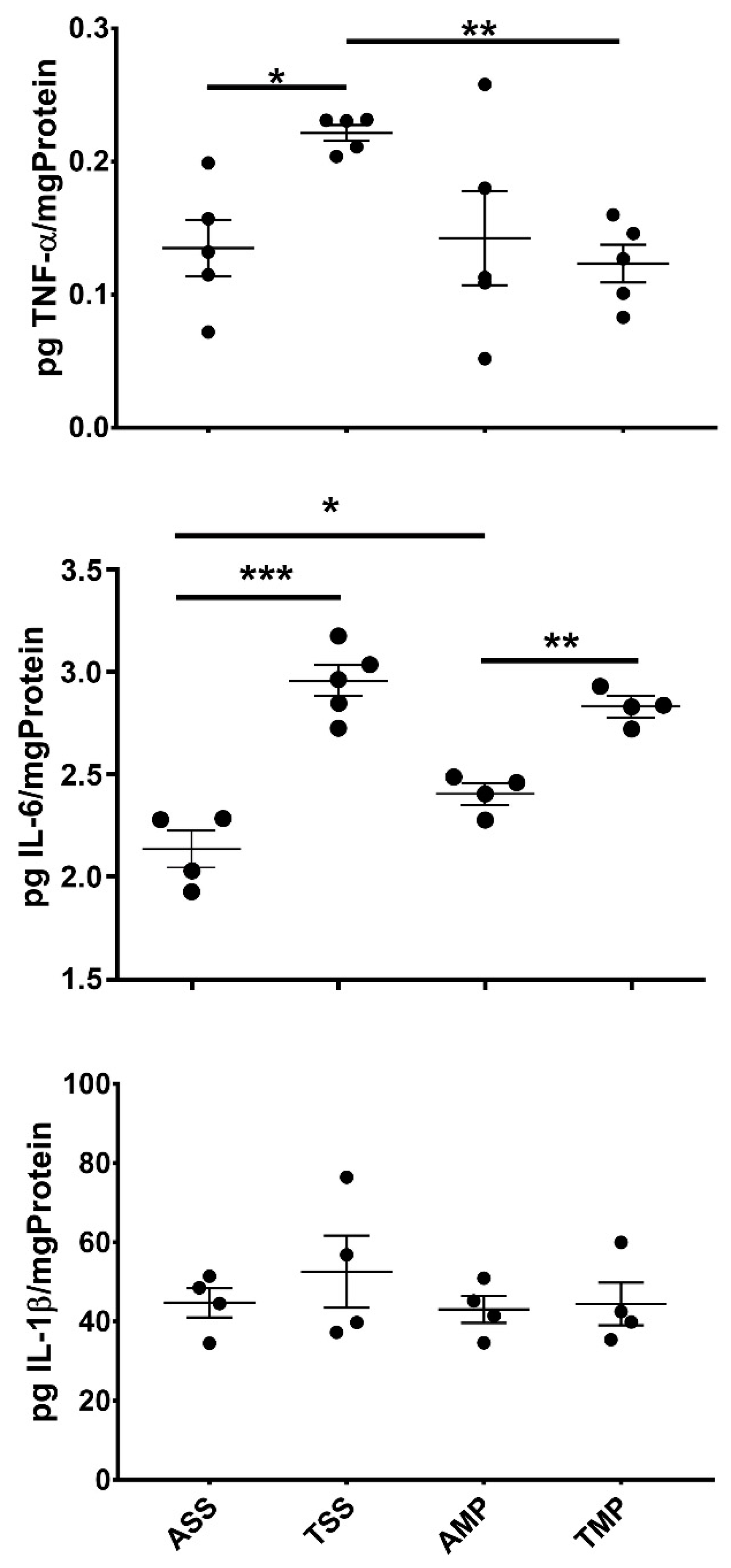
3.1. Effect of MP on the Central Levels of Cytokines
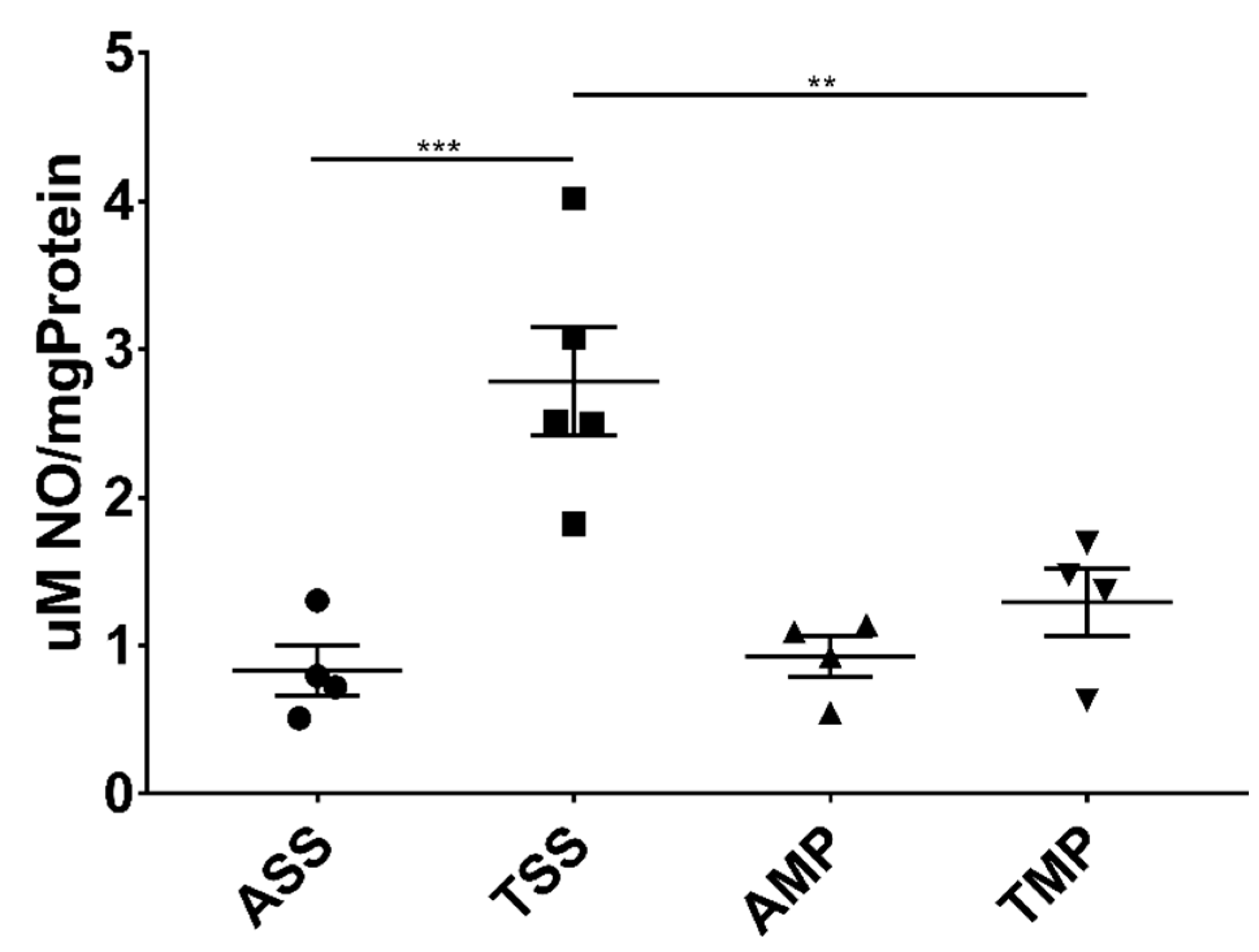
3.2. Intranasal Delivery of MP Effectively Reduced Central Levels of Nitric Oxide
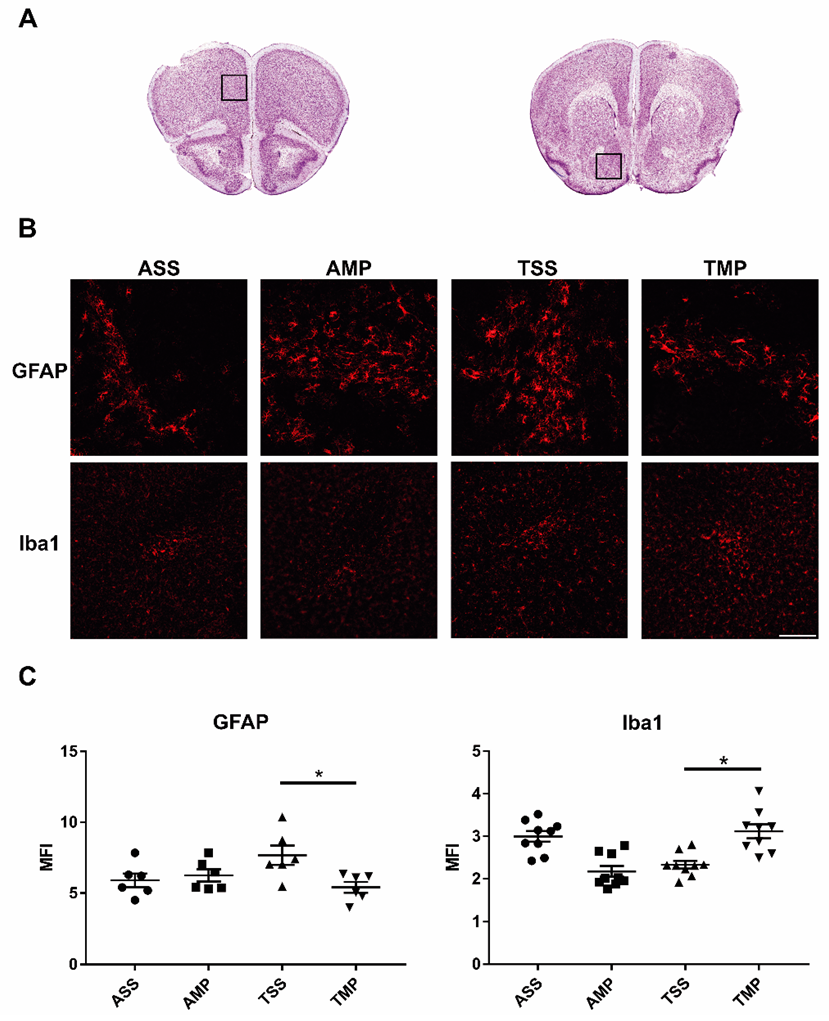
3.3. Expression of Iba-1 and GFAP in FC and NA after 24 of Treatment
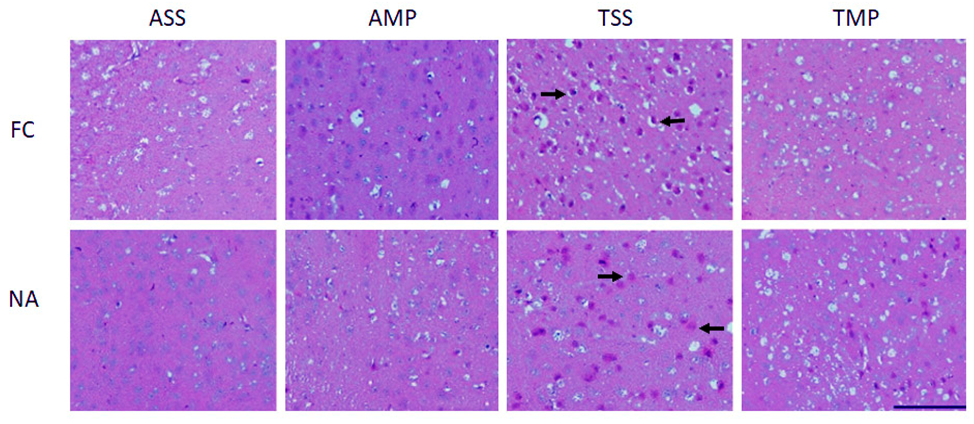
3.4. Reduced Histological Alterations by Intranasal Methylprednisolone
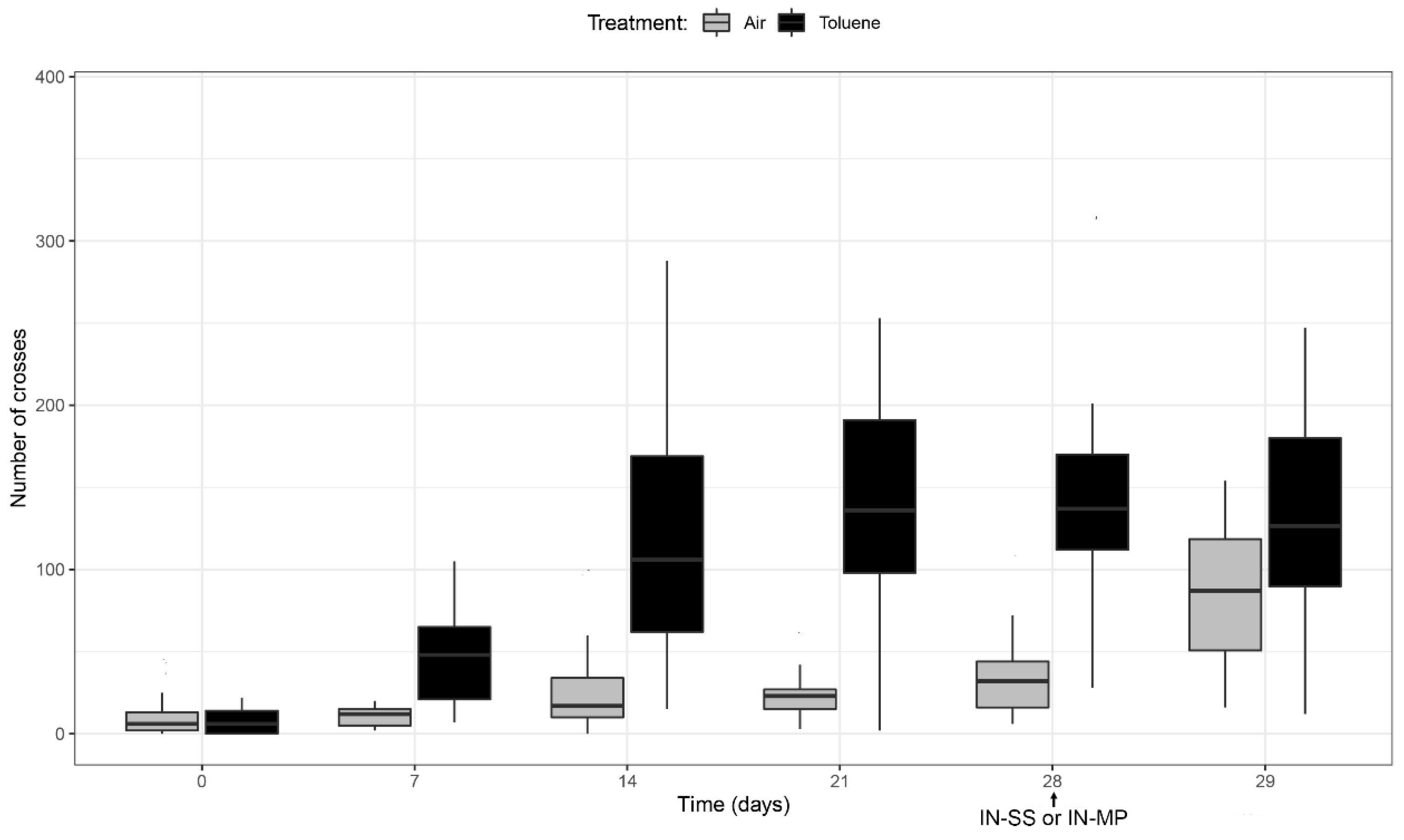
3.5. Analysis of Addictive Behavior Measuring the Locomotor Activity
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dell, C.A.; Gust, S.W.; MacLean, S. Global Issues in Volatile Substance Misuse. Subst. Use Misuse 2011, 46, 1–7. [Google Scholar] [CrossRef]
- Anthony, J.C.; Warner, L.A.; Kessler, R.C. Comparative epidemiology of dependence on tobacco, alcohol, controlled substances, and inhalants: Basic findings from the National Comorbidity Survey. Exp. Clin. Psychopharmacol. 1994, 2, 244–268. [Google Scholar] [CrossRef]
- Cruz, S.L.; Rivera-García, M.T.; Woodward, J.J. Review of Toluene Actions: Clinical Evidence, Animal Studies, and Molecular Targets. J. Drug Alcohol Res. 2014, 3, 235840. [Google Scholar] [CrossRef] [Green Version]
- Villatoro, J.A.; Cruz, S.L.; Ortiz, A.; Medina-Mora, M.E. Volatile Substance Misuse in Mexico: Correlates and Trends. Subst. Use Misuse 2011, 46, 40–45. [Google Scholar] [CrossRef]
- Cruz, S.L.; Torres-Flores, M.; Galván, E.J. Repeated toluene exposure alters the synaptic transmission of layer 5 medial prefrontal cortex. Neurotoxicol. Teratol. 2019, 73, 9–14. [Google Scholar] [CrossRef]
- Paez-Martinez, N.; Flores-Serrano, Z.; Ortiz-Lopez, L.; Ramirez-Rodriguez, G. Environmental enrichment increases doublecortin-associated new neurons and decreases neuronal death without modifying anxiety-like behavior in mice chronically exposed to toluene. Behav. Brain Res. 2013, 256, 432–440. [Google Scholar] [CrossRef]
- Ulloque, R.A. Sistema cerebral del placer y de la drogodependencia. Biomédica 1999, 19, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Callan, S.P.; Hannigan, J.H.; Bowen, S.E. Prenatal toluene exposure impairs performance in the Morris Water Maze in adolescent rats. Neuroscience 2017, 342, 180–187. [Google Scholar] [CrossRef] [Green Version]
- Montes, S.; del Carmen Solís-Guillén, R.; García-Jácome, D.; Páez-Martínez, N. Environmental enrichment reverses memory impairment induced by toluene in mice. Neurotoxicol. Teratol. 2017, 61, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Beckley, J.T.; Woodward, J.J. Volatile Solvents as Drugs of Abuse: Focus on the Cortico-Mesolimbic Circuitry. Neuropsychopharmacology 2013, 38, 2555–2567. [Google Scholar] [CrossRef] [Green Version]
- Beckstead, M.J.; Weiner, J.L.; Eger, E.I.; Gong, D.H.; Mihic, S.J. Glycine and gamma-aminobutyric acid(A) receptor function is enhanced by inhaled drugs of abuse. Mol. Pharmacol. 2000, 57, 1199–1205. [Google Scholar] [PubMed]
- Cruz, S.L.; Balster, R.L.; Woodward, J.J. Effects of volatile solvents on recombinant N-methyl-D-aspartate receptors expressed in Xenopus oocytes. Br. J. Pharmacol. 2000, 131, 1303–1308. [Google Scholar] [CrossRef] [Green Version]
- Perrine, S.A.; O’Leary-Moore, S.K.; Galloway, M.P.; Hannigan, J.H.; Bowen, S.E. Binge toluene exposure alters glutamate, glutamine and GABA in the adolescent rat brain as measured by proton magnetic resonance spectroscopy. Drug Alcohol Depend. 2011, 115, 101–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Zhou, J. Neuroinflammation in the central nervous system: Symphony of glial cells. Glia 2019, 67, 1017–1035. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.W.; Zhang, X.I.A.; Huang, W.J. Role of neuroinflammation in neurodegenerative diseases (Review). Mol. Med. Rep. 2016, 13, 3391–3396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, S.L.; Armenta-Reséndiz, M.; Carranza-Aguilar, C.J.; Galván, E.J. Minocycline prevents neuronal hyperexcitability and neuroinflammation in medial prefrontal cortex, as well as memory impairment caused by repeated toluene inhalation in adolescent rats. Toxicol. Appl. Pharmacol. 2020, 395, 114980. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Shurtleff, D.; Harris, R.A. Neuroimmune Mechanisms of Alcohol and Drug Addiction. Int. Rev. Neurobiol. 2014, 118, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Win-Shwe, T.-T.; Kunugita, N.; Yoshida, Y.; Fujimaki, H. Role of hippocampal TLR4 in neurotoxicity in mice following toluene exposure. Neurotoxicol. Teratol. 2011, 33, 598–602. [Google Scholar] [CrossRef] [PubMed]
- Win-Shwe, T.T.; Kunugita, N.; Yoshida, Y.; Nakajima, D.; Tsukahara, S.; Fujimaki, H. Differential mRNA expression of neuroimmune markers in the hippocampus of infant mice following toluene exposure during brain developmental period. J. Appl. Toxicol. 2012, 32, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Gotohda, T.; Tokunaga, I.; Kubo, S.I.; Morita, K.; Kitamura, O.; Eguchi, A. Effect of toluene inhalation on astrocytes and neurotrophic factor in rat brain. Forensic Sci. Int. 2000, 113, 233–238. [Google Scholar] [CrossRef]
- Cruz, S.L.; Bowen, S.E. The last two decades on preclinical and clinical research on inhalant effects. Neurotoxicol. Teratol. 2021, 87, 106999. [Google Scholar] [CrossRef]
- Dell, D.; Hopkins, C. Residential Volatile Substance Misuse Treatment for Indigenous Youth in Canada. Subst. Use Misuse 2011, 46, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Real, T.; Cruz, S.L.; Medina-Mora, M.E. Inhalant Addiction. In Textbook of Addiction Treatment: International Perspectives; Springer: Milan, Italy, 2015; pp. 597–619. [Google Scholar]
- Filley, C.M.; Halliday, W.; Kleinschmidt-Demasters, B.K. The Effects of Toluene on the Central Nervous System. J. Neuropathol. Exp. Neurol. 2004, 63, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Barnes, P.J. How corticosteroids control inflammation: Quintiles Prize Lecture 2005. Br. J. Pharmacol. 2006, 148, 245–254. [Google Scholar] [CrossRef]
- Whitehouse, M.W. Anti-inflammatory glucocorticoid drugs: Reflections after 60 years. Inflammopharmacology 2011, 19, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, S.; Cidlowski, J.A. Corticosteroids. Rheum. Dis. Clin. N. Am. 2016, 42, 15–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuckermann, J.P.; Kleiman, A.; McPherson, K.G.; Reichardt, H.M. Molecular Mechanisms of Glucocorticoids in the Control of Inflammation and Lymphocyte Apoptosis. Crit. Rev. Clin. Lab. Sci. 2005, 42, 71–104. [Google Scholar] [CrossRef] [PubMed]
- Erdő, F.; Bors, L.A.; Farkas, D.; Bajza, Á.; Gizurarson, S. Evaluation of intranasal delivery route of drug administration for brain targeting. Brain Res. Bull. 2018, 143, 155–170. [Google Scholar] [CrossRef]
- Rassy, D.; Bárcena, B.; Pérez-Osorio, I.N.; Espinosa, A.; Peón, A.N.; Terrazas, L.I.; Meneses, G.; Besedovsky, H.O.; Fragoso, G.; Sciutto, E. Intranasal Methylprednisolone Effectively Reduces Neuroinflammation in Mice with Experimental Autoimmune Encephalitis. J. Neuropathol. Exp. Neurol. 2020, 79, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Lungare, S.; Bowen, J.; Badhan, R. Development and Evaluation of a Novel Intranasal Spray for the Delivery of Amantadine. J. Pharm. Sci. 2016, 105, 1209–1220. [Google Scholar] [CrossRef]
- Páez-Martínez, N.; Pellicer, F.; González-Trujano, M.E.; Cruz-López, B. Environmental enrichment reduces behavioural sensitization in mice previously exposed to toluene: The role of D1 receptors. Behav. Brain Res. 2020, 390, 112624. [Google Scholar] [CrossRef] [PubMed]
- Demır, M.; Cicek, M.; Eser, N.; Yoldaş, A.; Sısman, T. Effects of Acute Toluene Toxicity on Different Regions of Rabbit Brain. Anal. Cell. Pathol. 2017, 2017, 2805370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paxinos, G.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates, 3rd ed.; Academic Press: New York, NY, USA, 2007; ISBN 978-0-12-369460-7. [Google Scholar]
- Meneses, G.; Gevorkian, G.; Florentino, A.; Bautista, M.A.; Espinosa, A.; Acero, G.; Díaz, G.; Fleury, A.; Osorio, I.N.P.; del Rey, A.; et al. Intranasal delivery of dexamethasone efficiently controls LPS-induced murine neuroinflammation. Clin. Exp. Immunol. 2017, 190, 304–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, J.M. Protein Protocols Handbook; Humana Press: Totowa, NJ, USA, 2002. [Google Scholar] [CrossRef]
- Meneses, G.; Cárdenas, G.; Espinosa, A.; Rassy, D.; Pérez-Osorio, I.N.; Bárcena, B.; Fleury, A.; Besedovsky, H.; Fragoso, G.; Sciutto, E. Sepsis: Developing new alternatives to reduce neuroinflammation and attenuate brain injury. Ann. N. Y. Acad. Sci. 2019, 1437, 43–56. [Google Scholar] [CrossRef]
- Bryan, N.S.; Grisham, M.B. Methods to detect nitric oxide and its metabolites in biological samples. Free Radic. Biol. Med. 2007, 43, 645–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aydin, K.; Sencer, S.; Demir, T.; Ogel, K.; Tunaci, A.; Minareci, O. Cranial MR findings in chronic toluene abuse by inhalation. AJNR Am. J. Neuroradiol. 2002, 23, 1173–1179. [Google Scholar]
- Montes, S.; Yee-Rios, Y.; Páez-Martínez, N. Environmental enrichment restores oxidative balance in animals chronically exposed to toluene: Comparison with melatonin. Brain Res. Bull. 2019, 144, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Celik, F.; Göçmez, C.; Kamaşak, K.; Tufek, A.; Guzel, A.; Tokgoz, O.; Fırat, U.; Evliyaoğlu, O. The comparison of neuroprotective effects of intrathecal dexmedetomidine and metilprednisolone in spinal cord injury. Int. J. Surg. 2013, 11, 414–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Liu, X.; Chen, S.; Xiao, Y.; Zhuang, W. Oral versus intravenous methylprednisolone for the treatment of multiple sclerosis relapses: A meta-analysis of randomized controlled trials. PLoS ONE 2017, 12, e0188644. [Google Scholar] [CrossRef] [Green Version]
- Hendler, J.V.; de Souza, L.; de Freitas Zernow, D.C.; Guimarães, J.; Bertotto, T.; Saldanha, C.F.; Brenol, J.C.T.; Monticielo, O.A. Survival analysis of patients with systemic lupus erythematosus in a tertiary hospital in southern Brazil. Clin. Rheumatol. 2017, 36, 2005–2010. [Google Scholar] [CrossRef]
- Tischner, D.; Reichardt, H.M. Glucocorticoids in the control of neuroinflammation. Mol. Cell. Endocrinol. 2007, 275, 62–70. [Google Scholar] [CrossRef] [Green Version]
- Djupesland, P.G.; Messina, J.C.; Mahmoud, R.A. The nasal approach to delivering treatment for brain diseases: An anatomic, physiologic, and delivery technology overview. Ther. Deliv. 2014, 5, 709–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducharme, N.; Banks, W.A.; Morley, J.E.; Robinson, S.M.; Niehoff, M.L.; Mattern, C. Brain distribution and behavioral effects of progesterone and pregnenolone after intranasal or intravenous administration. Eur. J. Pharmacol. 2010, 641, 128–134. [Google Scholar] [CrossRef] [Green Version]
- Chapman, C.D.; Frey, W.H.; Craft, S.; Danielyan, L.; Hallschmid, M.; Schiöth, H.B.; Benedict, C. Intranasal Treatment of Central Nervous System Dysfunction in Humans. Pharm. Res. 2013, 30, 2475–2484. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, A.; Meneses, G.; Chavarría, A.; Mancilla, R.; Pedraza-Chaverri, J.; Fleury, A.; Bárcena, B.; Pérez-Osorio, I.N.; Besedovsky, H.; Arauz, A.; et al. Intranasal Dexamethasone Reduces Mortality and Brain Damage in a Mouse Experimental Ischemic Stroke Model. Neurotherapeutics 2020, 17, 1907–1918. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Guo, S.-W.; Zhu, L.; Xu, X.; Liu, J. Methylprednisolone Induces Neuro-Protective Effects via the Inhibition of A1 Astrocyte Activation in Traumatic Spinal Cord Injury Mouse Models. Front. Neurosci. 2021, 15, 628917. [Google Scholar] [CrossRef] [PubMed]
- Oberleithner, H.; Riethmüller, C.; Ludwig, T.; Shahin, V.; Stock, C.; Schwab, A.; Hausberg, M.; Kusche, K.; Schillers, H. Differential action of steroid hormones on human endothelium. J. Cell Sci. 2006, 119, 1926–1932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerber, A.N.; Newton, R.; Sasse, S.K. Repression of transcription by the glucocorticoid receptor: A parsimonious model for the genomics era. J. Biol. Chem. 2021, 296, 100687. [Google Scholar] [CrossRef]
- Gao, Q.; Li, Y.; Shen, L.; Zhang, J.; Zheng, X.; Qu, R.; Liu, Z.; Chopp, M. Bone marrow stromal cells reduce ischemia-induced astrocytic activation in vitro. Neuroscience 2008, 152, 646–655. [Google Scholar] [CrossRef] [Green Version]
- Spooren, A.; Kolmus, K.; Laureys, G.; Clinckers, R.; De Keyser, J.; Haegeman, G.; Gerlo, S. Interleukin-6, a mental cytokine. Brain Res. Rev. 2011, 67, 157–183. [Google Scholar] [CrossRef]
- Sakai, T.; Honda, S.; Kuzuhara, S. Encephalomyelopathy demonstrated on MRI in a case of chronic toluene intoxication. Rinsho Shinkeigaku 2000, 40, 571–575. [Google Scholar]
- Tu, G.; Shi, Y.; Zheng, Y.; Ju, M.; He, H.; Ma, G.; Hao, G.; Luo, Z. Glucocorticoid attenuates acute lung injury through induction of type 2 macrophage. J. Transl. Med. 2017, 15, 181. [Google Scholar] [CrossRef]
- Ciriaco, M.; Ventrice, P.; Russo, G.; Scicchitano, M.; Mazzitello, G.; Scicchitano, F.; Russo, E. Corticosteroid-related central nervous system side effects. J. Pharmacol. Pharmacother. 2013, 4, 94. [Google Scholar] [CrossRef] [Green Version]
- Guerri, C.; Pascual, M. Impact of neuroimmune activation induced by alcohol or drug abuse on adolescent brain development. Int. J. Dev. Neurosci. 2019, 77, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Harricharan, R.; Abboussi, O.; Daniels, W.M.U. Addiction: A dysregulation of satiety and inflammatory processes. Prog. Brain Res. 2017, 235, 65–91. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giraldo-Velásquez, M.F.; Pérez-Osorio, I.N.; Espinosa-Cerón, A.; Bárcena, B.M.; Calderón-Gallegos, A.; Fragoso, G.; Torres-Ramos, M.; Páez-Martínez, N.; Sciutto, E. Intranasal Methylprednisolone Ameliorates Neuroinflammation Induced by Chronic Toluene Exposure. Pharmaceutics 2022, 14, 1195. https://doi.org/10.3390/pharmaceutics14061195
Giraldo-Velásquez MF, Pérez-Osorio IN, Espinosa-Cerón A, Bárcena BM, Calderón-Gallegos A, Fragoso G, Torres-Ramos M, Páez-Martínez N, Sciutto E. Intranasal Methylprednisolone Ameliorates Neuroinflammation Induced by Chronic Toluene Exposure. Pharmaceutics. 2022; 14(6):1195. https://doi.org/10.3390/pharmaceutics14061195
Chicago/Turabian StyleGiraldo-Velásquez, Manuel F., Iván N. Pérez-Osorio, Alejandro Espinosa-Cerón, Brandon M. Bárcena, Arturo Calderón-Gallegos, Gladis Fragoso, Mónica Torres-Ramos, Nayeli Páez-Martínez, and Edda Sciutto. 2022. "Intranasal Methylprednisolone Ameliorates Neuroinflammation Induced by Chronic Toluene Exposure" Pharmaceutics 14, no. 6: 1195. https://doi.org/10.3390/pharmaceutics14061195
APA StyleGiraldo-Velásquez, M. F., Pérez-Osorio, I. N., Espinosa-Cerón, A., Bárcena, B. M., Calderón-Gallegos, A., Fragoso, G., Torres-Ramos, M., Páez-Martínez, N., & Sciutto, E. (2022). Intranasal Methylprednisolone Ameliorates Neuroinflammation Induced by Chronic Toluene Exposure. Pharmaceutics, 14(6), 1195. https://doi.org/10.3390/pharmaceutics14061195