

Direct Potential Modulation of Neurogenic Differentiation Markers by Granulocyte-Colony Stimulating Factor (G-CSF) in the Rodent Brain

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Procedure
2.3. Cutting of Brains
2.4. Isolation of RNA
2.5. DNase Digest
2.6. Isolation of Proteins
2.7. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.8. Western Blot
2.9. Statistics
3. Results
3.1. Differentiation Markers in Classic Anatomical Areas of Neurogenesis
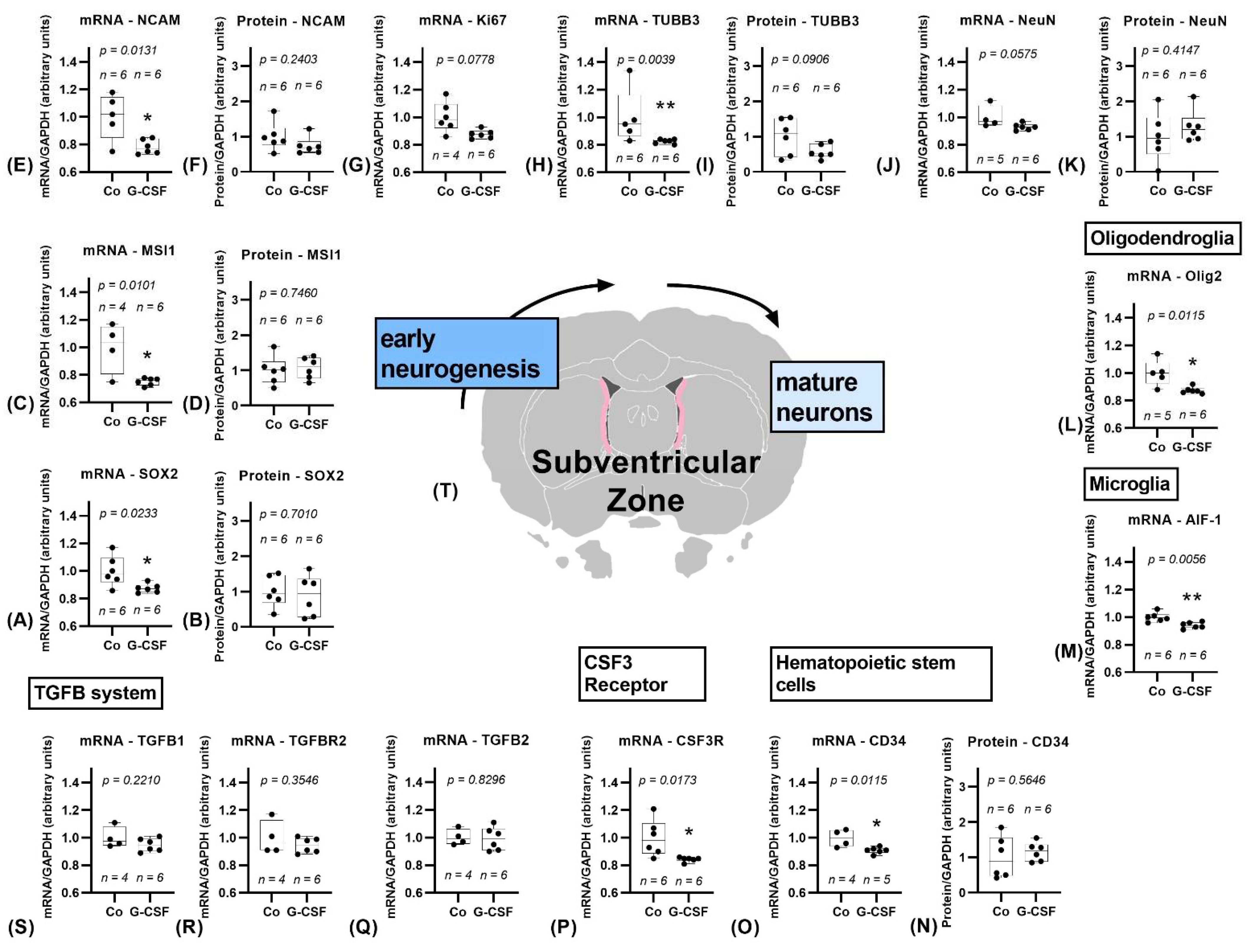
3.1.1. Subventricular Zone
3.1.2. Hippocampus
3.2. Differentiation Markers in Further Anatomic Regions
3.2.1. Corpus Callosum
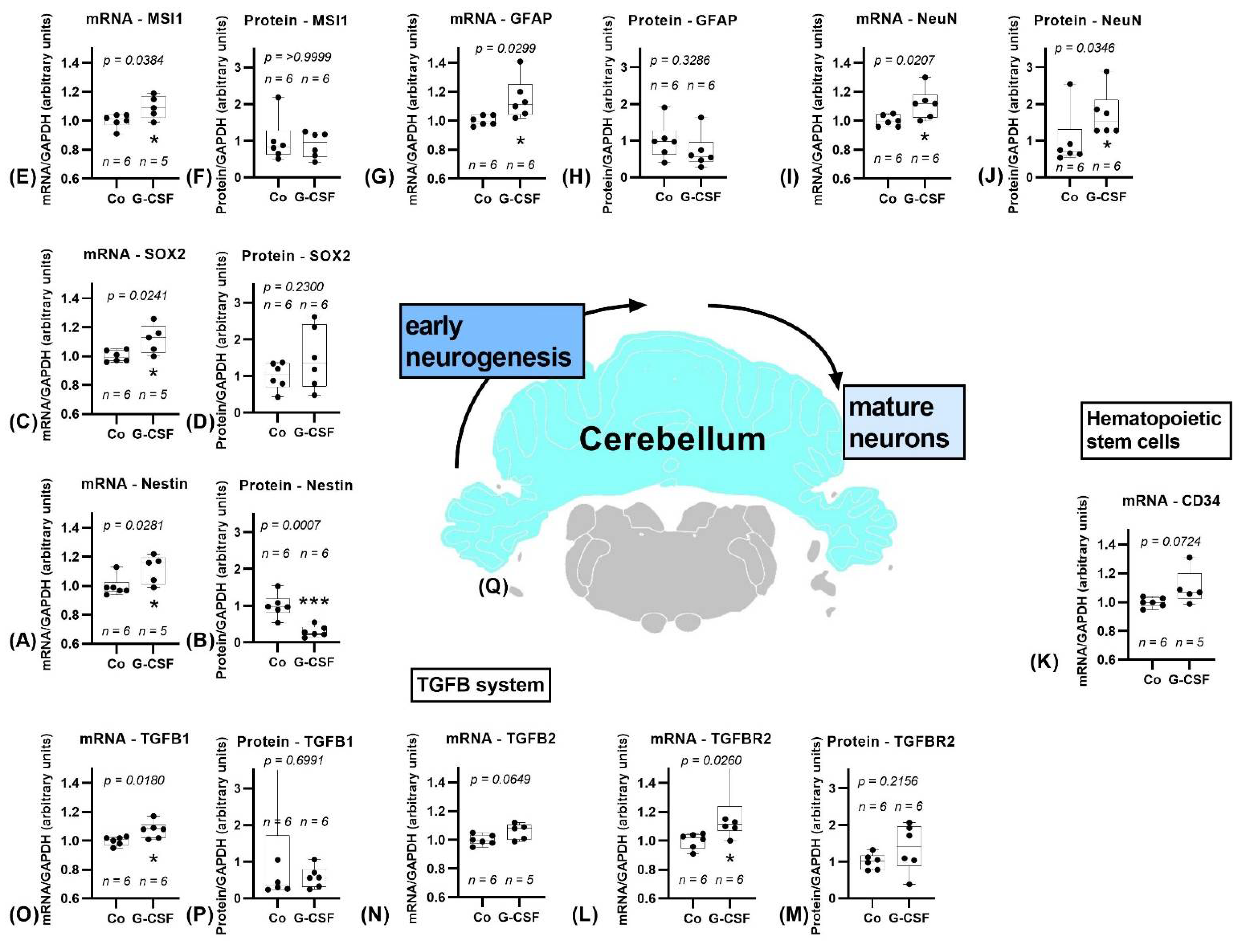
3.2.2. Cerebellum
3.2.3. Hypothalamus
3.2.4. Prefrontal Cortex
3.2.5. Motor Cortex
3.2.6. Putamen
3.2.7. Nucleus Accumbens
3.2.8. Substantia Nigra
3.2.9. Pyramidal Tracts
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Welte, K. Granulozyten-Kolonien stimulierender Faktor (G-CSF): Biochemie, Biologie und Pathophysiologie. Klin. Padiatr. 1988, 200, 157–164. [Google Scholar] [CrossRef]
- Welte, K. G-CSF: Filgrastim, lenograstim and biosimilars. Expert Opin. Biol. Ther. 2014, 14, 983–993. [Google Scholar] [CrossRef]
- Kirsch, F.; Krüger, C.; Schneider, A. The receptor for Granulocyte-colony stimulating factor (G-CSF) is expressed in radial glia during development of the nervous system. BMC Dev. Biol. 2008, 8, 32. [Google Scholar] [CrossRef]
- Wallner, S.; Peters, S.; Pitzer, C.; Resch, H.; Bogdahn, U.; Schneider, A. The Granulocyte-colony stimulating factor has a dual role in neuronal and vascular plasticity. Front. Cell Dev. Biol. 2015, 3, 48. [Google Scholar] [CrossRef]
- Schneider, A.; Krüger, C.; Steigleder, T.; Weber, D.; Pitzer, C.; Laage, R.; Aronowski, J.; Maurer, M.H.; Gassler, N.; Mier, W.; et al. The hematopoietic factor G-CSF is a neuronal ligand that counteracts programmed cell death and drives neurogenesis. J. Clin. Investig. 2005, 115, 2083–2098. [Google Scholar] [CrossRef]
- Sehara, Y.; Hayashi, T.; Deguchi, K.; Zhang, H.; Tsuchiya, A.; Yamashita, T.; Lukic, V.; Nagai, M.; Kamiya, T.; Abe, K. Potentiation of neurogenesis and angiogenesis by G-CSF after focal cerebral ischemia in rats. Brain Res. 2007, 1151, 142–149. [Google Scholar] [CrossRef]
- Schmidt, A.K.; Reich, A.; Falkenburger, B.; Schulz, J.B.; Brandenburg, L.O.; Ribes, S.; Tauber, S.C. Adjuvant granulocyte colony-stimulating factor therapy results in improved spatial learning and stimulates hippocampal neurogenesis in a mouse model of pneumococcal meningitis. J. Neuropathol. Exp. Neurol. 2015, 74, 85–94. [Google Scholar] [CrossRef]
- Chiò, A.; Mora, G.; La Bella, V.; Caponnetto, C.; Mancardi, G.; Sabatelli, M.; Siciliano, G.; Silani, V.; Corbo, M.; Moglia, C.; et al. Repeated courses of granulocyte colony-stimulating factor in amyotrophic lateral sclerosis: Clinical and biological results from a prospective multicenter study. Muscle Nerve 2011, 43, 189–195. [Google Scholar] [CrossRef]
- Johannesen, S.; Budeus, B.; Peters, S.; Iberl, S.; Meyer, A.L.; Kammermaier, T.; Wirkert, E.; Bruun, T.H.; Samara, V.C.; Schulte-Mattler, W.; et al. Biomarker Supervised G-CSF (Filgrastim) Response in ALS Patients. Front. Neurol. 2018, 9, 971. [Google Scholar] [CrossRef]
- Tarella, C.; Melazzini, M.; Petrini, M.; Leone, G.; Scime’, R.; Soligo, D.; Gualandi, F.; Moglia, C.; Mancardi, G.; Silani, V.; et al. Repeated Courses of Bone Marrow-Derived Cell Mobilization Induced by Granulocyte-Colony Stimulating Factor (G-CSF) in Amyotrophic Lateral Sclerosis: Interim Analysis of a Prospective Multicenter Trial. Blood 2007, 110, 4065. [Google Scholar] [CrossRef]
- Dietrich, J.; Baryawno, N.; Nayyar, N.; Valtis, Y.K.; Yang, B.; Ly, I.; Besnard, A.; Severe, N.; Gustafsson, K.U.; Andronesi, O.C.; et al. Bone marrow drives central nervous system regeneration after radiation injury. J. Clin. Investig. 2018, 128, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Aschauer-Wallner, S.; Leis, S.; Bogdahn, U.; Johannesen, S.; Couillard-Despres, S.; Aigner, L. Granulocyte colony-stimulating factor in traumatic spinal cord injury. Drug Discov. Today 2021, 26, 1642–1655. [Google Scholar] [CrossRef] [PubMed]
- Johannesen, S.; Huie, J.R.; Budeus, B.; Peters, S.; Wirth, A.M.; Iberl, S.; Kammermaier, T.; Kobor, I.; Wirkert, E.; Küspert, S.; et al. Modeling and Bioinformatics Identify Responders to G-CSF in Patients with Amyotrophic Lateral Sclerosis. Front. Neurol. 2021, 12, 616289. [Google Scholar] [CrossRef]
- Altman, J. Are new neurons formed in the brains of adult mammals? Science 1962, 135, 1127–1128. [Google Scholar] [CrossRef]
- Kempermann, G. Neuroscience. What the bomb said about the brain. Science 2013, 340, 1180–1181. [Google Scholar] [CrossRef]
- Spalding, K.L.; Bergmann, O.; Alkass, K.; Bernard, S.; Salehpour, M.; Huttner, H.B.; Boström, E.; Westerlund, I.; Vial, C.; Buchholz, B.A.; et al. Dynamics of hippocampal neurogenesis in adult humans. Cell 2013, 153, 1219–1227. [Google Scholar] [CrossRef]
- Sorrells, S.F.; Paredes, M.F.; Cebrian-Silla, A.; Sandoval, K.; Qi, D.; Kelley, K.W.; James, D.; Mayer, S.; Chang, J.; Auguste, K.I.; et al. Human hippocampal neurogenesis drops sharply in children to undetectable levels in adults. Nature 2018, 555, 377–381. [Google Scholar] [CrossRef]
- Eriksson, P.S.; Perfilieva, E.; Björk-Eriksson, T.; Alborn, A.M.; Nordborg, C.; Peterson, D.A.; Gage, F.H. Neurogenesis in the adult human hippocampus. Nat. Med. 1998, 4, 1313–1317. [Google Scholar] [CrossRef]
- Fares, J.; Bou Diab, Z.; Nabha, S.; Fares, Y. Neurogenesis in the adult hippocampus: History, regulation, and prospective roles. Int. J. Neurosci. 2019, 129, 598–611. [Google Scholar] [CrossRef]
- Lendahl, U.; Zimmerman, L.B.; McKay, R.D. CNS stem cells express a new class of intermediate filament protein. Cell 1990, 60, 585–595. [Google Scholar] [CrossRef]
- Mullen, R.J.; Buck, C.R.; Smith, A.M. NeuN, a neuronal specific nuclear protein in vertebrates. Development 1992, 116, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Ellis, P.; Fagan, B.M.; Magness, S.T.; Hutton, S.; Taranova, O.; Hayashi, S.; McMahon, A.; Rao, M.; Pevny, L. SOX2, a persistent marker for multipotential neural stem cells derived from embryonic stem cells, the embryo or the adult. Dev. Neurosci. 2004, 26, 148–165. [Google Scholar] [CrossRef] [PubMed]
- Okano, H.; Kawahara, H.; Toriya, M.; Nakao, K.; Shibata, S.; Imai, T. Function of RNA-binding protein Musashi-1 in stem cells. Exp. Cell Res. 2005, 306, 349–356. [Google Scholar] [CrossRef]
- Guo, Z.; Wang, X.; Xiao, J.; Wang, Y.; Lu, H.; Teng, J.; Wang, W. Early postnatal GFAP-expressing cells produce multilineage progeny in cerebrum and astrocytes in cerebellum of adult mice. Brain Res. 2013, 1532, 14–20. [Google Scholar] [CrossRef]
- Couillard-Despres, S.; Quehl, E.; Altendorfer, K.; Karl, C.; Ploetz, S.; Bogdahn, U.; Winkler, J.; Aigner, L. Human in vitro reporter model of neuronal development and early differentiation processes. BMC Neurosci. 2008, 9, 31. [Google Scholar] [CrossRef]
- Jørgensen, O.S. Neural cell adhesion molecule (NCAM) as a quantitative marker in synaptic remodeling. Neurochem. Res. 1995, 20, 533–547. [Google Scholar] [CrossRef] [PubMed]
- Katsetos, C.D.; Herman, M.M.; Mörk, S.J. Class III beta-tubulin in human development and cancer. Cell Motil. Cytoskelet. 2003, 55, 77–96. [Google Scholar] [CrossRef]
- Gerdes, J.; Schwab, U.; Lemke, H.; Stein, H. Production of a mouse monoclonal antibody reactive with a human nuclear antigen associated with cell proliferation. Int. J. Cancer 1983, 31, 13–20. [Google Scholar] [CrossRef]
- Simmons, D.L.; Satterthwaite, A.B.; Tenen, D.G.; Seed, B. Molecular cloning of a cDNA encoding CD34, a sialomucin of human hematopoietic stem cells. J. Immunol. 1992, 148, 267–271. [Google Scholar]
- Massagué, J. TGFβ signalling in context. Nat. Rev. Mol. Cell Biol. 2012, 13, 616–630. [Google Scholar] [CrossRef]
- Zhou, Q.; Wang, S.; Anderson, D.J. Identification of a Novel Family of Oligodendrocyte Lineage-Specific Basic Helix–Loop–Helix Transcription Factors. Neuron 2000, 25, 331–343. [Google Scholar] [CrossRef] [Green Version]
- Deininger, M.H.; Meyermann, R.; Schluesener, H.J. The allograft inflammatory factor-1 family of proteins. FEBS Lett. 2002, 514, 115–121. [Google Scholar] [CrossRef]
- Inazawa, J.; Fukunaga, R.; Seto, Y.; Nakagawa, H.; Misawa, S.; Abe, T.; Nagata, S. Assignment of the human granulocyte colony-stimulating factor receptor gene (CSF3R) to chromosome 1 at region p35–p34.3. Genomics 1991, 10, 1075–1078. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 4th ed.; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Peters, S.; Zitzelsperger, E.; Kuespert, S.; Iberl, S.; Heydn, R.; Johannesen, S.; Petri, S.; Aigner, L.; Thal, D.R.; Hermann, A.; et al. The TGF-β System As a Potential Pathogenic Player in Disease Modulation of Amyotrophic Lateral Sclerosis. Front. Neurol. 2017, 8, 669. [Google Scholar] [CrossRef] [PubMed]
- Bakker, R.; Tiesinga, P.; Kötter, R. The Scalable Brain Atlas: Instant Web-Based Access to Public Brain Atlases and Related Content. Neuroinformatics 2015, 13, 353–366. [Google Scholar] [CrossRef]
- Papp, E.A.; Leergaard, T.B.; Calabrese, E.; Johnson, G.A.; Bjaalie, J.G. Waxholm Space atlas of the Sprague Dawley rat brain. Neuroimage 2014, 97, 374–386. [Google Scholar] [CrossRef]
- Kjonigsen, L.J.; Lillehaug, S.; Bjaalie, J.G.; Witter, M.P.; Leergaard, T.B. Waxholm Space atlas of the rat brain hippocampal region: Three-dimensional delineations based on magnetic resonance and diffusion tensor imaging. Neuroimage 2015, 108, 441–449. [Google Scholar] [CrossRef]
- Sergejeva, M.; Papp, E.A.; Bakker, R.; Gaudnek, M.A.; Okamura-Oho, Y.; Boline, J.; Bjaalie, J.G.; Hess, A. Anatomical landmarks for registration of experimental image data to volumetric rodent brain atlasing templates. J. Neurosci. Methods 2015, 240, 161–169. [Google Scholar] [CrossRef]
- Hamby, M.E.; Coskun, V.; Sun, Y.E. Transcriptional regulation of neuronal differentiation: The epigenetic layer of complexity. Biochim. Biophys. Acta 2008, 1779, 432–437. [Google Scholar] [CrossRef]
- Keiner, S.; Niv, F.; Neumann, S.; Steinbach, T.; Schmeer, C.; Hornung, K.; Schlenker, Y.; Förster, M.; Witte, O.W.; Redecker, C. Effect of skilled reaching training and enriched environment on generation of oligodendrocytes in the adult sensorimotor cortex and corpus callosum. BMC Neurosci. 2017, 18, 31. [Google Scholar] [CrossRef]
- Ahlfeld, J.; Filser, S.; Schmidt, F.; Wefers, A.K.; Merk, D.J.; Glaß, R.; Herms, J.; Schüller, U. Neurogenesis from Sox2 expressing cells in the adult cerebellar cortex. Sci. Rep. 2017, 7, 6137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kempermann, G. Environmental enrichment, new neurons and the neurobiology of individuality. Nat. Rev. Neurosci. 2019, 20, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Leal-Galicia, P.; Chávez-Hernández, M.E.; Mata, F.; Mata-Luévanos, J.; Rodríguez-Serrano, L.M.; Tapia-de-Jesús, A.; Buenrostro-Jáuregui, M.H. Adult Neurogenesis: A Story Ranging from Controversial New Neurogenic Areas and Human Adult Neurogenesis to Molecular Regulation. Int. J. Mol. Sci. 2021, 22, 11489. [Google Scholar] [CrossRef] [PubMed]
- Wojcinski, A.; Lawton, A.K.; Bayin, N.S.; Lao, Z.; Stephen, D.N.; Joyner, A.L. Cerebellar granule cell replenishment postinjury by adaptive reprogramming of Nestin+ progenitors. Nat. Neurosci. 2017, 20, 1361–1370. [Google Scholar] [CrossRef]
- Kokoeva, M.V.; Yin, H.; Flier, J.S. Evidence for constitutive neural cell proliferation in the adult murine hypothalamus. J. Comp. Neurol. 2007, 505, 209–220. [Google Scholar] [CrossRef]
- Pierce, A.A.; Xu, A.W. De novo neurogenesis in adult hypothalamus as a compensatory mechanism to regulate energy balance. J. Neurosci. 2010, 30, 723–730. [Google Scholar] [CrossRef]
- Kuhn, H.G.; Toda, T.; Gage, F.H. Adult Hippocampal Neurogenesis: A Coming-of-Age Story. J. Neurosci. 2018, 38, 10401–10410. [Google Scholar] [CrossRef]
- Boldrini, M.; Fulmore, C.A.; Tartt, A.N.; Simeon, L.R.; Pavlova, I.; Poposka, V.; Rosoklija, G.B.; Stankov, A.; Arango, V.; Dwork, A.J.; et al. Human Hippocampal Neurogenesis Persists throughout Aging. Cell Stem Cell 2018, 22, 589–599.e5. [Google Scholar] [CrossRef]
- Lim, D.A.; Alvarez-Buylla, A. The Adult Ventricular-Subventricular Zone (V-SVZ) and Olfactory Bulb (OB) Neurogenesis. Cold Spring Harb. Perspect. Biol. 2016, 8, a018820. [Google Scholar] [CrossRef]
- Abbott, L.C.; Nigussie, F. Adult neurogenesis in the mammalian dentate gyrus. Anat. Histol. Embryol. 2020, 49, 3–16. [Google Scholar] [CrossRef]
- Dumbuya, J.S.; Chen, L.; Wu, J.Y.; Wang, B. The role of G-CSF neuroprotective effects in neonatal hypoxic-ischemic encephalopathy (HIE): Current status. J. Neuroinflammation 2021, 18, 55. [Google Scholar] [CrossRef] [PubMed]
- Weise, G.; Pösel, C.; Möller, K.; Kranz, A.; Didwischus, N.; Boltze, J.; Wagner, D.C. High-dosage granulocyte colony stimulating factor treatment alters monocyte trafficking to the brain after experimental stroke. Brain Behav. Immun. 2017, 60, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Dale, D.C.; Crawford, J.; Klippel, Z.; Reiner, M.; Osslund, T.; Fan, E.; Morrow, P.K.; Allcott, K.; Lyman, G.H. A systematic literature review of the efficacy, effectiveness, and safety of filgrastim. Support. Care Cancer 2018, 26, 7–20. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kozole, J.; Heydn, R.; Wirkert, E.; Küspert, S.; Aigner, L.; Bruun, T.-H.; Bogdahn, U.; Peters, S.; Johannesen, S. Direct Potential Modulation of Neurogenic Differentiation Markers by Granulocyte-Colony Stimulating Factor (G-CSF) in the Rodent Brain. Pharmaceutics 2022, 14, 1858. https://doi.org/10.3390/pharmaceutics14091858
Kozole J, Heydn R, Wirkert E, Küspert S, Aigner L, Bruun T-H, Bogdahn U, Peters S, Johannesen S. Direct Potential Modulation of Neurogenic Differentiation Markers by Granulocyte-Colony Stimulating Factor (G-CSF) in the Rodent Brain. Pharmaceutics. 2022; 14(9):1858. https://doi.org/10.3390/pharmaceutics14091858
Chicago/Turabian StyleKozole, Judith, Rosmarie Heydn, Eva Wirkert, Sabrina Küspert, Ludwig Aigner, Tim-Henrik Bruun, Ulrich Bogdahn, Sebastian Peters, and Siw Johannesen. 2022. "Direct Potential Modulation of Neurogenic Differentiation Markers by Granulocyte-Colony Stimulating Factor (G-CSF) in the Rodent Brain" Pharmaceutics 14, no. 9: 1858. https://doi.org/10.3390/pharmaceutics14091858
APA StyleKozole, J., Heydn, R., Wirkert, E., Küspert, S., Aigner, L., Bruun, T. -H., Bogdahn, U., Peters, S., & Johannesen, S. (2022). Direct Potential Modulation of Neurogenic Differentiation Markers by Granulocyte-Colony Stimulating Factor (G-CSF) in the Rodent Brain. Pharmaceutics, 14(9), 1858. https://doi.org/10.3390/pharmaceutics14091858