


Bovine Lactoferrin Suppresses Tumor Angiogenesis through NF-κB Pathway Inhibition by Binding to TRAF6
 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Cell Culture
2.2. Proliferation Assay
2.3. Wound Healing Assay
2.4. Migration Assay
2.5. Tube Formation Assay
2.6. Detection of Apoptosis Using Flow Cytometry Analysis
2.7. Western Blot Analysis
2.8. Gene Expression Experiments
2.9. Quantitative RT-PCR
2.10. Small Interfering RNA (siRNA) Gene Knockdown
2.11. In Vivo Experiment
2.12. Immunohistochemical Staining
2.13. Statistical Analysis
3. Results
3.1. bLF Significantly Inhibits Angiogenic Ability of TECs but Not NECs
3.2. bLF Suppresses Tube Formation of TECs but Promotes That of NECs
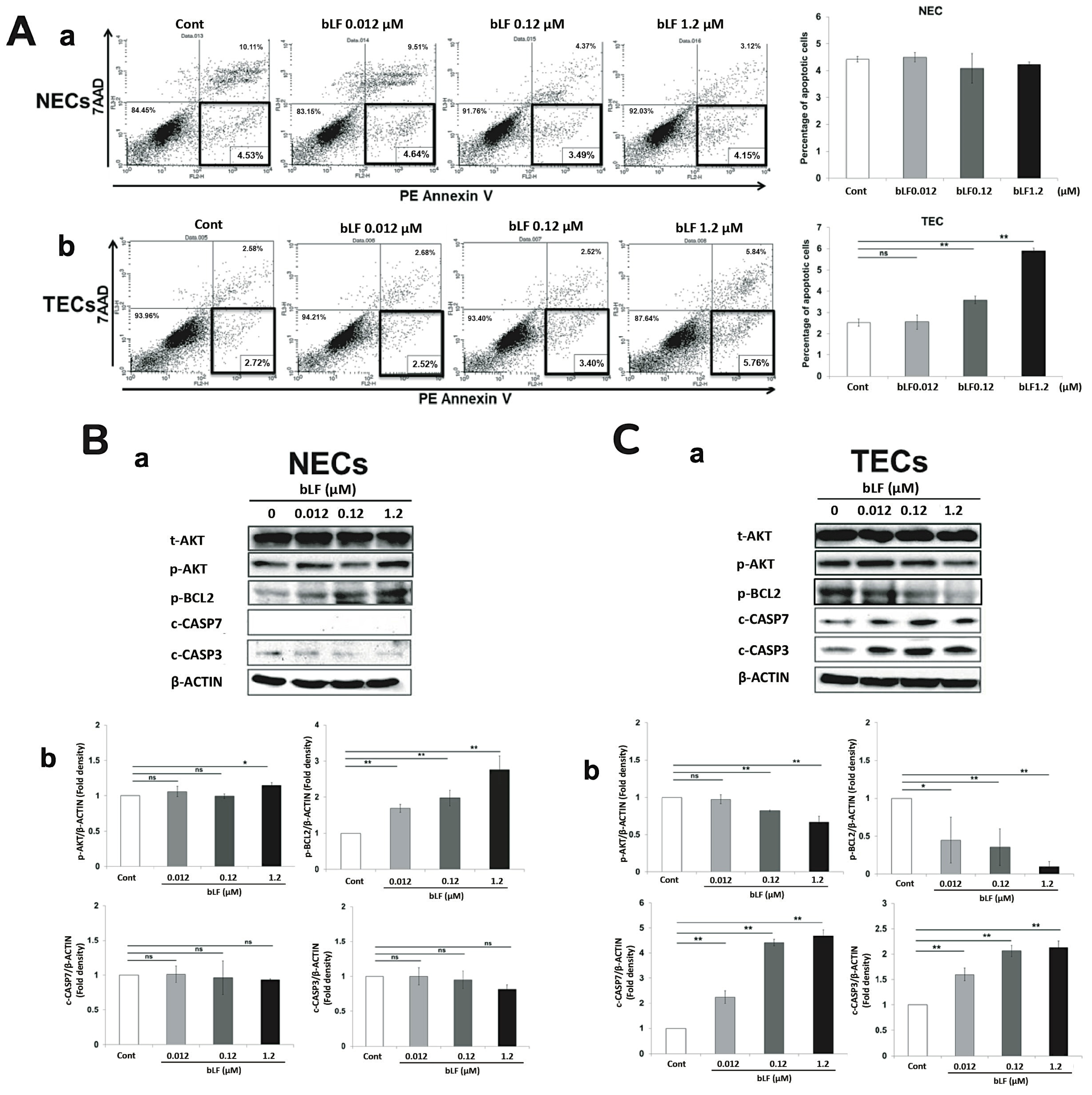
3.3. bLF Induces Apoptosis of TECs through Activation of Apoptotic Effector Caspases
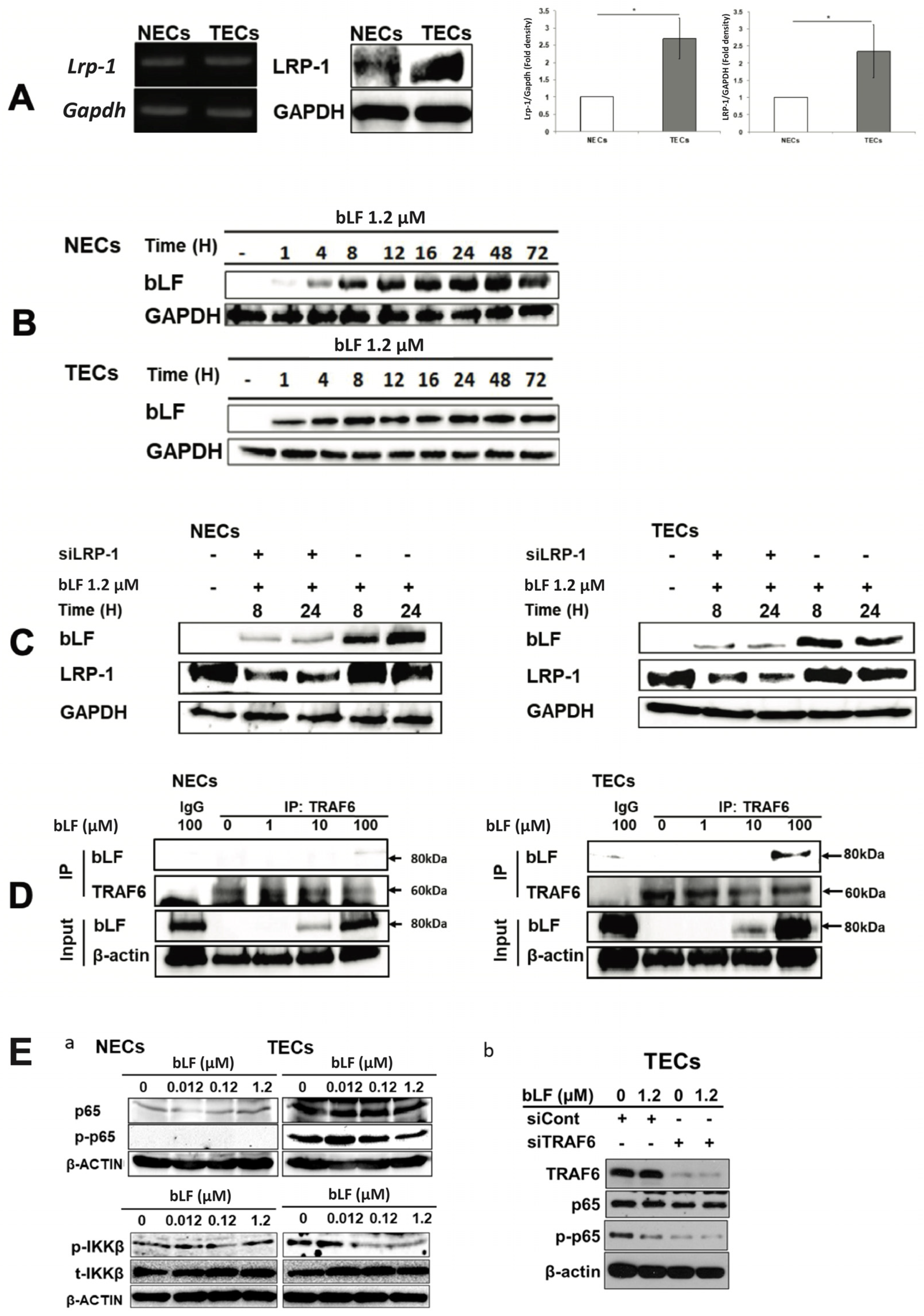
3.4. bLF Is Internalized in NECs and TECs through LRP-1
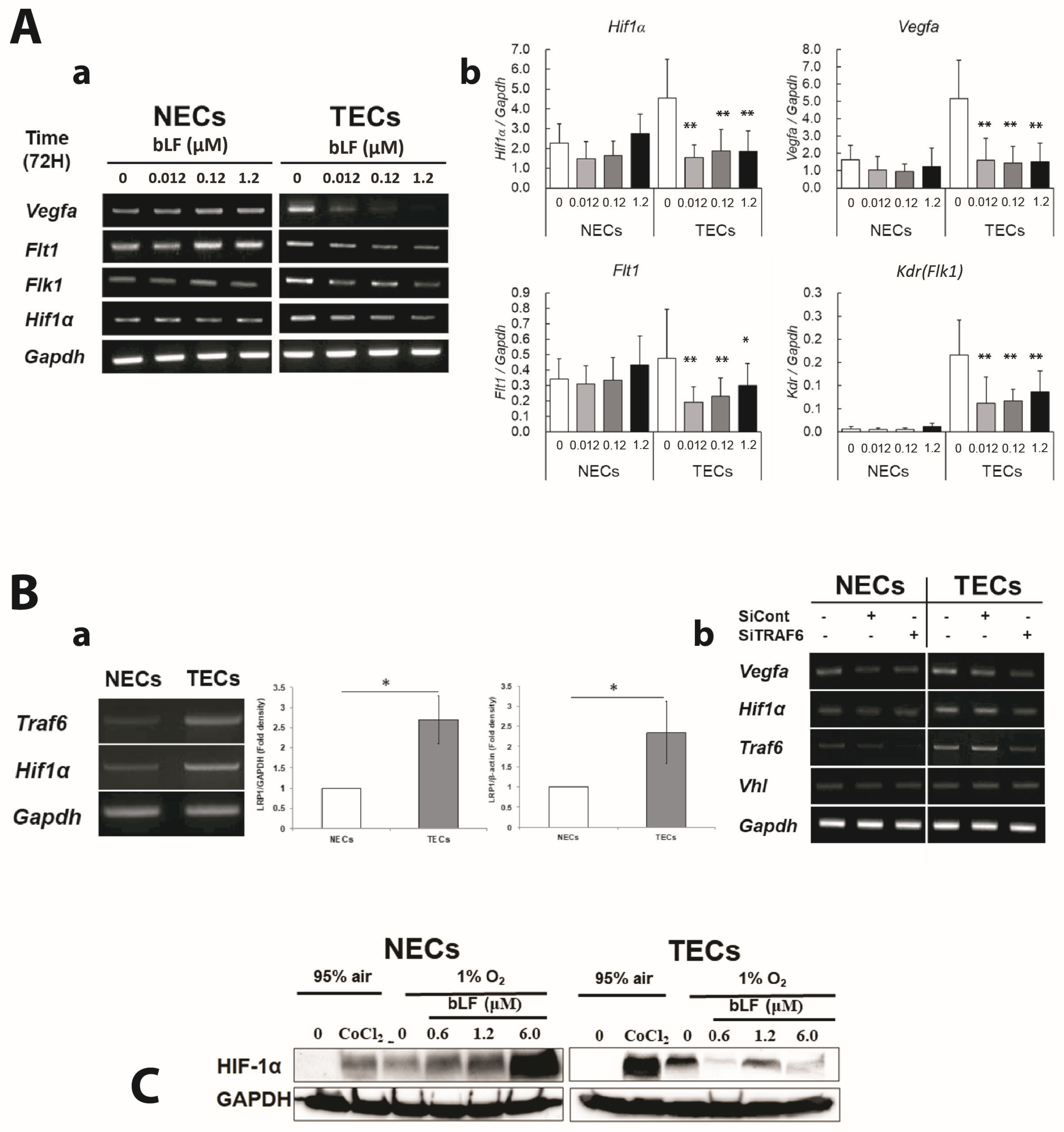
3.5. bLF down Regulates Proangiogenic Genes and Its Transcription Factor through Inhibiting NF-κB Activation in TECs
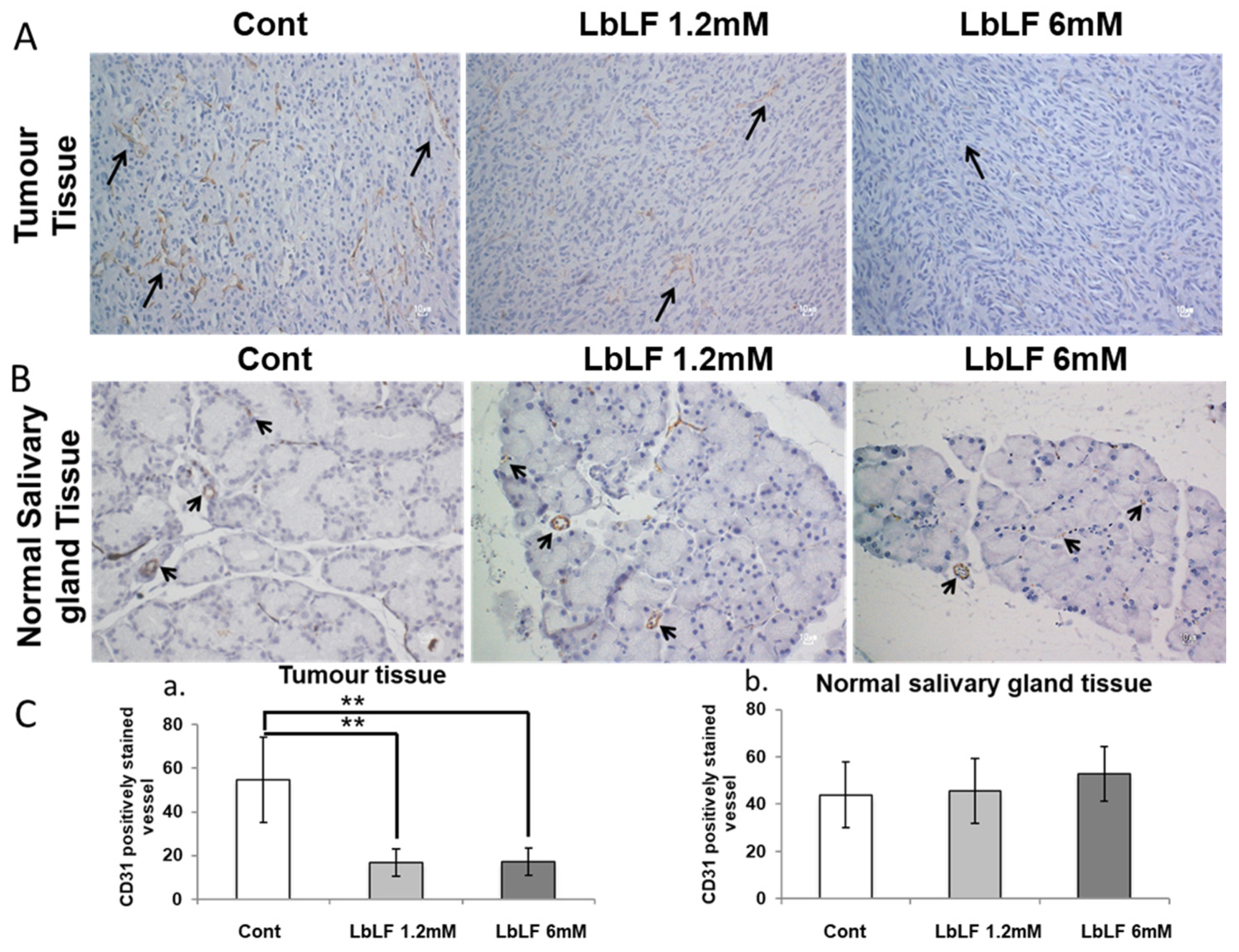
3.6. Effects of Orally Administrated LbLF on Microvascular Density in the In Vivo OSCC Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Folkman, J. Clinical Applications of Research on Angiogenesis. N. Engl. J. Med. 1995, 333, 1757–1763. [Google Scholar] [CrossRef]
- Hanahan, D.; Folkman, J. Patterns and Emerging Mechanisms of the Angiogenic Switch during Tumorigenesis. Cell 1996, 86, 353–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Risau, W. Mechanisms of Angiogenesis. Nature 1997, 386, 671–674. [Google Scholar] [CrossRef] [PubMed]
- Folkman, J. Tumor Angiogenesis: Therapeutic Implications. N. Engl. J. Med. 1971, 285, 1182–1186. [Google Scholar] [CrossRef] [PubMed]
- Folkman, J. Angiogenesis. Annu. Rev. Med. 2006, 57, 1–18. [Google Scholar] [CrossRef]
- Kerbel, R.S.; Yu, J.; Tran, J.; Man, S.; Viloria-Petit, A.; Klement, G.; Coomber, B.L.; Rak, J. Possible Mechanisms of Acquired Resistance to Anti-Angiogenic Drugs: Implications for the Use of Combination Therapy Approaches. Cancer Metastasis Rev. 2001, 20, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N. VEGF-A: A Critical Regulator of Blood Vessel Growth. Eur. Cytokine Netw. 2009, 20, 158–163. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, N.; Gerber, H.-P.; LeCouter, J. The Biology of VEGF and Its Receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef]
- Shibuya, M.; Claesson-Welsh, L. Signal Transduction by VEGF Receptors in Regulation of Angiogenesis and Lymphangiogenesis. Exp. Cell Res. 2006, 312, 549–560. [Google Scholar] [CrossRef]
- Bluff, J.E.; Menakuru, S.R.; Cross, S.S.; Higham, S.E.; Balasubramanian, S.P.; Brown, N.J.; Reed, M.W.; Staton, C.A. Angiogenesis Is Associated with the Onset of Hyperplasia in Human Ductal Breast Disease. Br. J. Cancer 2009, 101, 666–672. [Google Scholar] [CrossRef]
- Guo, S.; Colbert, L.S.; Fuller, M.; Zhang, Y.; Gonzalez-Perez, R.R. Vascular Endothelial Growth Factor Receptor-2 in Breast Cancer. Biochim. Biophys. Acta 2010, 1806, 108–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsson, A.-K.; Dimberg, A.; Kreuger, J.; Claesson-Welsh, L. VEGF Receptor Signalling—In Control of Vascular Function. Nat. Rev. Mol. Cell Biol. 2006, 7, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Waltenberger, J.; Claesson-Welsh, L.; Siegbahn, A.; Shibuya, M.; Heldin, C.H. Different Signal Transduction Properties of KDR and Flt1, Two Receptors for Vascular Endothelial Growth Factor. J. Biol. Chem. 1994, 269, 26988–26995. [Google Scholar] [CrossRef] [PubMed]
- Zachary, I. Vascular Endothelial Growth Factor: How It Transmits Its Signal. Exp. Nephrol. 1998, 6, 480–487. [Google Scholar] [CrossRef]
- Semenza, G.L. Hypoxia, Clonal Selection, and the Role of HIF-1 in Tumor Progression. Crit. Rev. Biochem. Mol. Biol. 2000, 35, 71–103. [Google Scholar] [CrossRef] [PubMed]
- Chaudary, N.; Hill, R.P. Hypoxia and Metastasis in Breast Cancer. Breast Dis. 2006, 26, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; Davis-Smyth, T. The Biology of Vascular Endothelial Growth Factor. Endocr. Rev. 1997, 18, 4–25. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Hou, M.; Guan, Y.; Jiang, M.; Yang, Y.; Gou, H. Expression of HIF-1alpha and VEGF in Colorectal Cancer: Association with Clinical Outcomes and Prognostic Implications. BMC Cancer 2009, 9, 432. [Google Scholar] [CrossRef] [Green Version]
- Tamano, S.; Sekine, K.; Takase, M.; Yamauchi, K.; Iigo, M.; Tsuda, H. Lack of Chronic Oral Toxicity of Chemopreventive Bovine Lactoferrin in F344/DuCrj Rats. Asian Pac. J. Cancer Prev. 2008, 9, 313–316. [Google Scholar]
- Sekine, K.; Watanabe, E.; Nakamura, J.; Takasuka, N.; Kim, D.J.; Asamoto, M.; Krutovskikh, V.; Baba-Toriyama, H.; Ota, T.; Moore, M.A.; et al. Inhibition of Azoxymethane-Initiated Colon Tumor by Bovine Lactoferrin Administration in F344 Rats. Jpn. J. Cancer Res. 1997, 88, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-Y.; Li, M.; Luo, C.-C.; Wang, J.-Q.; Zheng, N. Lactoferrin Exerts Antitumor Effects by Inhibiting Angiogenesis in a HT29 Human Colon Tumor Model. J. Agric. Food Chem. 2017, 65, 10464–10472. [Google Scholar] [CrossRef]
- Rodriguez-Ochoa, N.; Cortes-Reynosa, P.; Rodriguez-Rojas, K.; de la Garza, M.; Salazar, E.P. Bovine Holo-Lactoferrin Inhibits Migration and Invasion in MDA-MB-231 Breast Cancer Cells. Mol. Biol. Rep. 2022; Online ahead of print. [Google Scholar] [CrossRef]
- Chea, C.; Miyauchi, M.; Inubushi, T.; Febriyanti Ayuningtyas, N.; Subarnbhesaj, A.; Nguyen, P.T.; Shrestha, M.; Haing, S.; Ohta, K.; Takata, T. Molecular Mechanism of Inhibitory Effects of Bovine Lactoferrin on the Growth of Oral Squamous Cell Carcinoma. PLoS ONE 2018, 13, e0191683. [Google Scholar] [CrossRef] [Green Version]
- Roseanu, A.; Florian, P.E.; Moisei, M.; Sima, L.E.; Evans, R.W.; Trif, M. Liposomalization of Lactoferrin Enhanced Its Anti-Tumoral Effects on Melanoma Cells. Biometals 2010, 23, 485–492. [Google Scholar] [CrossRef]
- Cutone, A.; Rosa, L.; Ianiro, G.; Lepanto, M.S.; Bonaccorsi di Patti, M.C.; Valenti, P.; Musci, G. Lactoferrin’s Anti-Cancer Properties: Safety, Selectivity, and Wide Range of Action. Biomolecules 2020, 10, 456. [Google Scholar] [CrossRef] [Green Version]
- Kozu, T.; Iinuma, G.; Ohashi, Y.; Saito, Y.; Akasu, T.; Saito, D.; Alexander, D.B.; Iigo, M.; Kakizoe, T.; Tsuda, H. Effect of Orally Administered Bovine Lactoferrin on the Growth of Adenomatous Colorectal Polyps in a Randomized, Placebo-Controlled Clinical Trial. Cancer Prev. Res. 2009, 2, 975–983. [Google Scholar] [CrossRef] [Green Version]
- Iigo, M.; Alexander, D.B.; Xu, J.; Futakuchi, M.; Suzui, M.; Kozu, T.; Akasu, T.; Saito, D.; Kakizoe, T.; Yamauchi, K.; et al. Inhibition of Intestinal Polyp Growth by Oral Ingestion of Bovine Lactoferrin and Immune Cells in the Large Intestine. Biometals 2014, 27, 1017–1029. [Google Scholar] [CrossRef] [Green Version]
- Hida, K.; Klagsbrun, M. A New Perspective on Tumor Endothelial Cells: Unexpected Chromosome and Centrosome Abnormalities. Cancer Res. 2005, 65, 2507–2510. [Google Scholar] [CrossRef] [Green Version]
- Tsumita, T.; Maishi, N.; Annan, D.A.-M.; Towfik, M.A.; Matsuda, A.; Onodera, Y.; Nam, J.-M.; Hida, Y.; Hida, K. The Oxidized-LDL/LOX-1 Axis in Tumor Endothelial Cells Enhances Metastasis by Recruiting Neutrophils and Cancer Cells. Int. J. Cancer 2022, 151, 944–956. [Google Scholar] [CrossRef]
- Inubushi, T.; Kawazoe, A.; Miyauchi, M.; Kudo, Y.; Ao, M.; Ishikado, A.; Makino, T.; Takata, T. Molecular Mechanisms of the Inhibitory Effects of Bovine Lactoferrin on Lipopolysaccharide-Mediated Osteoclastogenesis. J. Biol. Chem. 2012, 287, 23527–23536. [Google Scholar] [CrossRef] [Green Version]
- van Uden, P.; Kenneth, N.S.; Rocha, S. Regulation of Hypoxia-Inducible Factor-1alpha by NF-KappaB. Biochem. J. 2008, 412, 477–484. [Google Scholar] [CrossRef]
- Sun, H.; Li, X.-B.; Meng, Y.; Fan, L.; Li, M.; Fang, J. TRAF6 Upregulates Expression of HIF-1α and Promotes Tumor Angiogenesis. Cancer Res. 2013, 73, 4950–4959. [Google Scholar] [CrossRef] [Green Version]
- Folkman, J.; Klagsbrun, M. Angiogenic Factors. Science 1987, 235, 442–447. [Google Scholar] [CrossRef]
- Theodoropoulos, V.E.; Lazaris, A.C.; Sofras, F.; Gerzelis, I.; Tsoukala, V.; Ghikonti, I.; Manikas, K.; Kastriotis, I. Hypoxia-Inducible Factor 1 Alpha Expression Correlates with Angiogenesis and Unfavorable Prognosis in Bladder Cancer. Eur. Urol. 2004, 46, 200–208. [Google Scholar] [CrossRef]
- Tung, Y.-T.; Chen, H.-L.; Yen, C.-C.; Lee, P.-Y.; Tsai, H.-C.; Lin, M.-F.; Chen, C.-M. Bovine Lactoferrin Inhibits Lung Cancer Growth through Suppression of Both Inflammation and Expression of Vascular Endothelial Growth Factor. J. Dairy Sci. 2013, 96, 2095–2106. [Google Scholar] [CrossRef] [Green Version]
- Sagar, S.M.; Yance, D.; Wong, R.K. Natural Health Products That Inhibit Angiogenesis: A Potential Source for Investigational New Agents to Treat Cancer-Part 1. Curr. Oncol. 2006, 13, 14–26. [Google Scholar] [CrossRef]
- Hida, K.; Hida, Y.; Amin, D.N.; Flint, A.F.; Panigrahy, D.; Morton, C.C.; Klagsbrun, M. Tumor-Associated Endothelial Cells with Cytogenetic Abnormalities. Cancer Res. 2004, 64, 8249–8255. [Google Scholar] [CrossRef] [Green Version]
- Harada, H.; Itasaka, S.; Kizaka-Kondoh, S.; Shibuya, K.; Morinibu, A.; Shinomiya, K.; Hiraoka, M. The Akt/MTOR Pathway Assures the Synthesis of HIF-1alpha Protein in a Glucose- and Reoxygenation-Dependent Manner in Irradiated Tumors. J. Biol. Chem. 2009, 284, 5332–5342. [Google Scholar] [CrossRef] [Green Version]
- Westphal, D.; Dewson, G.; Czabotar, P.E.; Kluck, R.M. Molecular Biology of Bax and Bak Activation and Action. Biochim. Biophys. Acta 2011, 1813, 521–531. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Stockard, C.R.; Harkins, L.; Lott, P.; Salih, C.; Yuan, K.; Buchsbaum, D.; Hashim, A.; Zayzafoon, M.; Hardy, R.W.; et al. Immunohistochemistry in the Evaluation of Neovascularization in Tumor Xenografts. Biotech. Histochem. Off. Publ. Biol. Stain Comm. 2008, 83, 179–189. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayuningtyas, N.F.; Chea, C.; Ando, T.; Saninggar, K.E.; Tanimoto, K.; Inubushi, T.; Maishi, N.; Hida, K.; Shindoh, M.; Miyauchi, M.; et al. Bovine Lactoferrin Suppresses Tumor Angiogenesis through NF-κB Pathway Inhibition by Binding to TRAF6. Pharmaceutics 2023, 15, 165. https://doi.org/10.3390/pharmaceutics15010165
Ayuningtyas NF, Chea C, Ando T, Saninggar KE, Tanimoto K, Inubushi T, Maishi N, Hida K, Shindoh M, Miyauchi M, et al. Bovine Lactoferrin Suppresses Tumor Angiogenesis through NF-κB Pathway Inhibition by Binding to TRAF6. Pharmaceutics. 2023; 15(1):165. https://doi.org/10.3390/pharmaceutics15010165
Chicago/Turabian StyleAyuningtyas, Nurina Febriyanti, Chanbora Chea, Toshinori Ando, Karina Erda Saninggar, Keiji Tanimoto, Toshihiro Inubushi, Nako Maishi, Kyoko Hida, Masanobu Shindoh, Mutsumi Miyauchi, and et al. 2023. "Bovine Lactoferrin Suppresses Tumor Angiogenesis through NF-κB Pathway Inhibition by Binding to TRAF6" Pharmaceutics 15, no. 1: 165. https://doi.org/10.3390/pharmaceutics15010165
APA StyleAyuningtyas, N. F., Chea, C., Ando, T., Saninggar, K. E., Tanimoto, K., Inubushi, T., Maishi, N., Hida, K., Shindoh, M., Miyauchi, M., & Takata, T. (2023). Bovine Lactoferrin Suppresses Tumor Angiogenesis through NF-κB Pathway Inhibition by Binding to TRAF6. Pharmaceutics, 15(1), 165. https://doi.org/10.3390/pharmaceutics15010165