
New Insights into the Behavior of NHC-Gold Complexes in Cancer Cells
 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
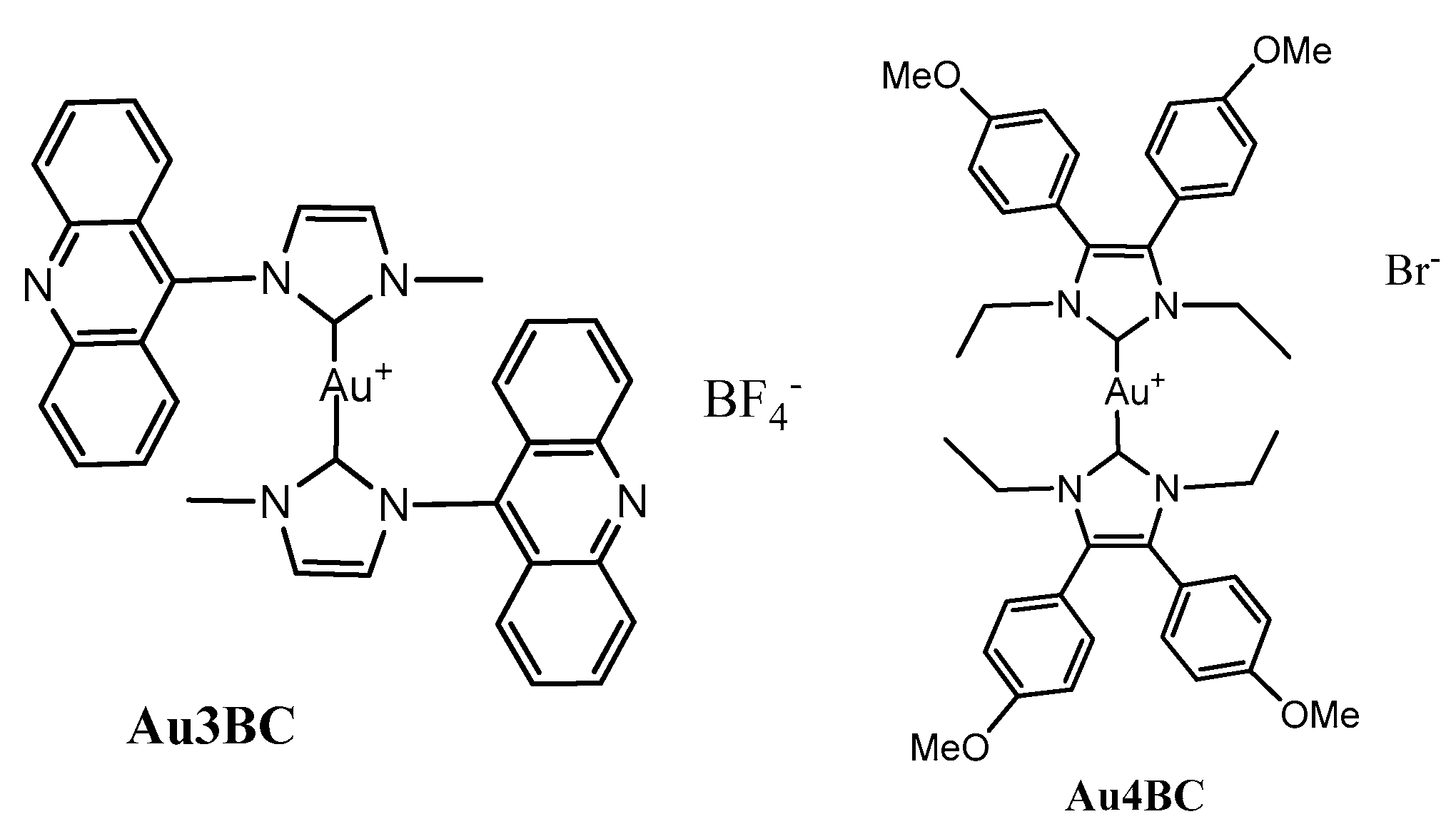
2.1. Synthesis of Compound Bis[1-methyl-3-acridineimidazolin-2-ylidene]gold(I) Tetrafluoroborate (Au3BC)
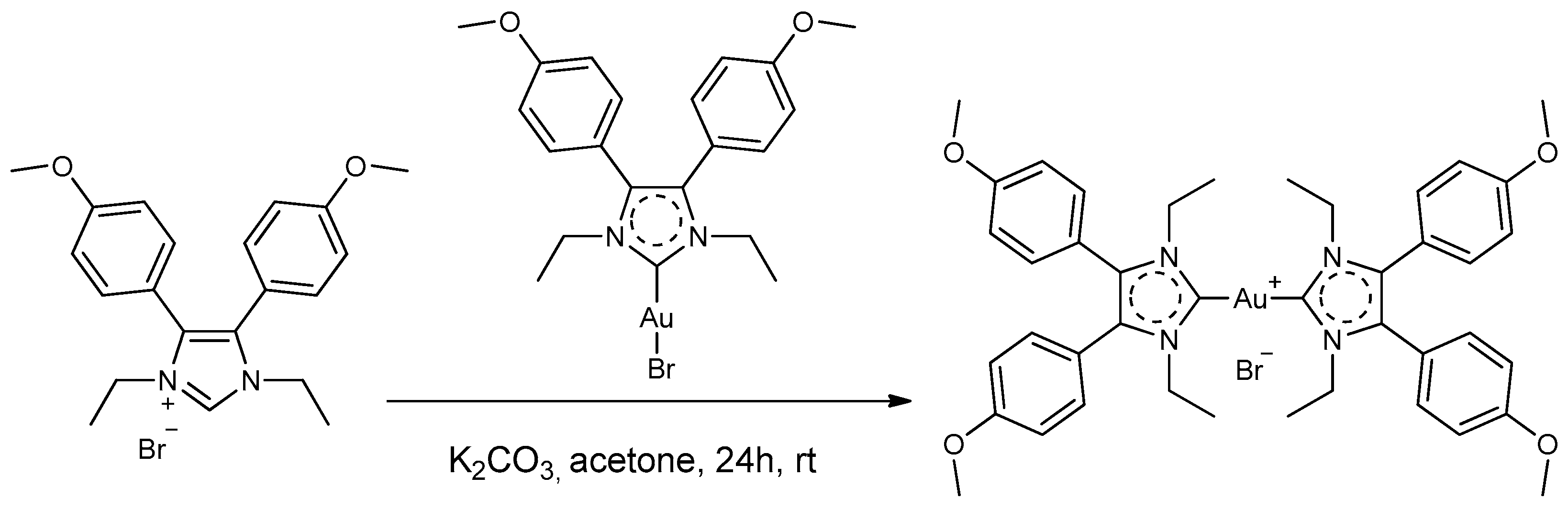
2.2. Synthesis of Compound Bis[1,3-diethyl-4,5-bis(4-methoxyphenyl)imidazol-2-ylidene]gold(I) Bromide (Au4BC)
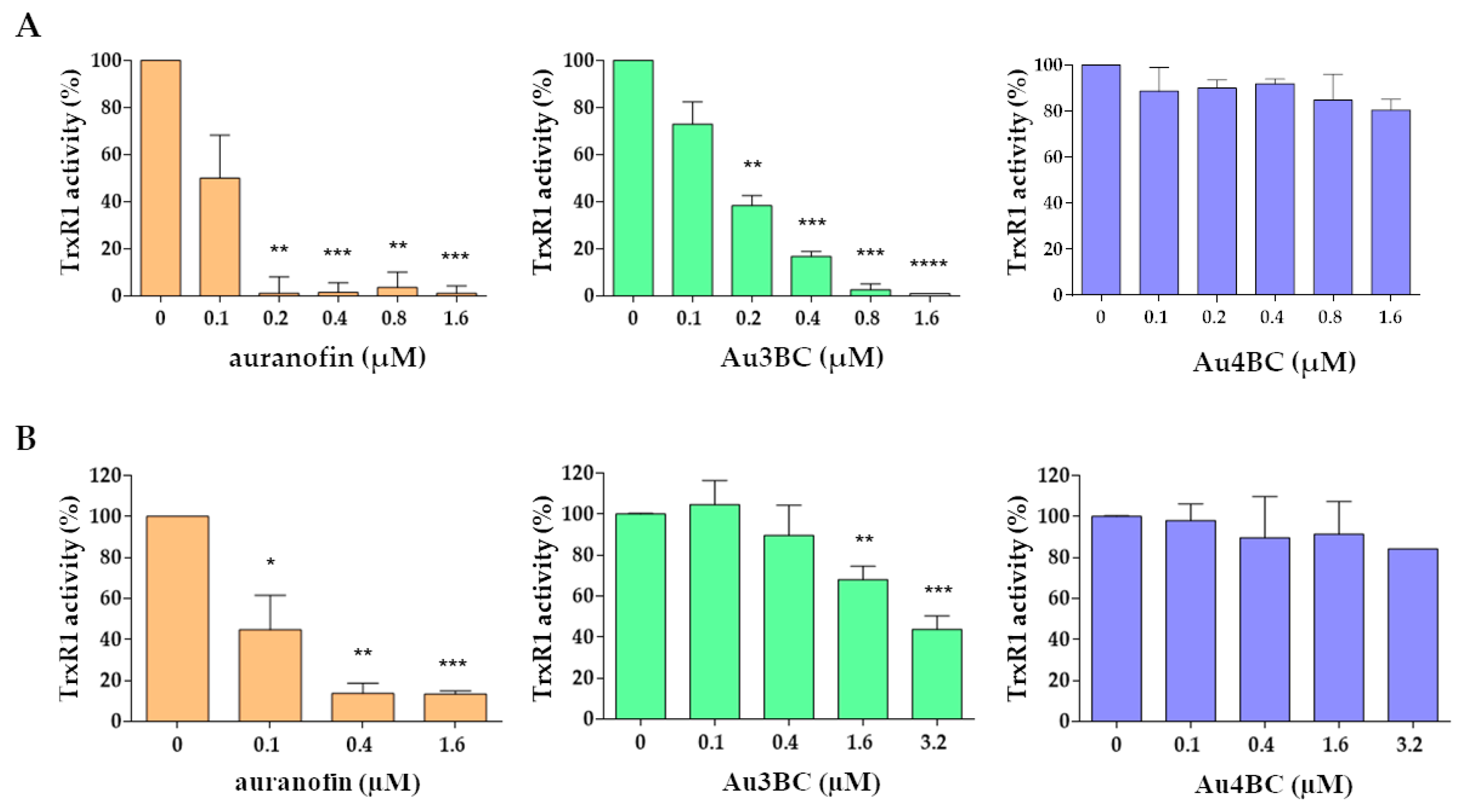
2.3. Measurement of TrxR Activity
2.4. Measurement of Trx1 Activity
2.5. Cell Lines and Cell Culture
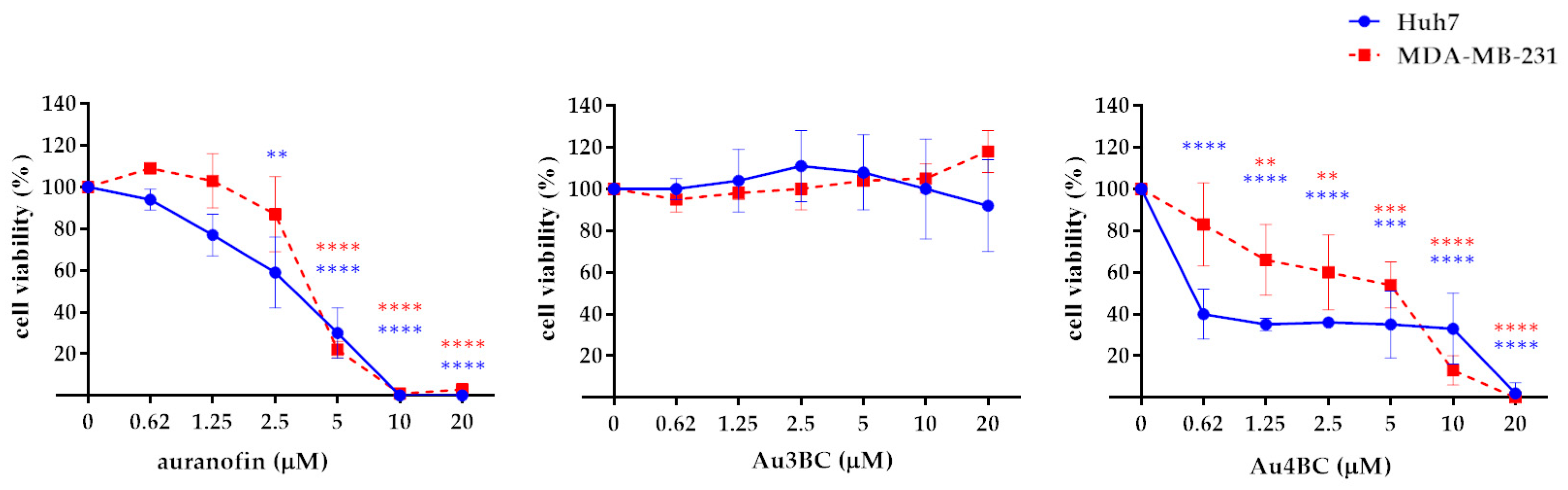
2.6. Cell Viability
2.7. Intracellular ROS Detection
2.8. Western Blotting
2.9. Cell Transfection
2.10. Statistical Analyses
3. Results
3.1. Selection and Synthesis of Gold Complexes
3.2. Biological Assays
3.2.1. Thioredoxin Reductase 1 (TrxR1) Activity
3.2.2. Effect of Au3BC and Au4BC on Cell Viability of HCC and BC Cell Lines
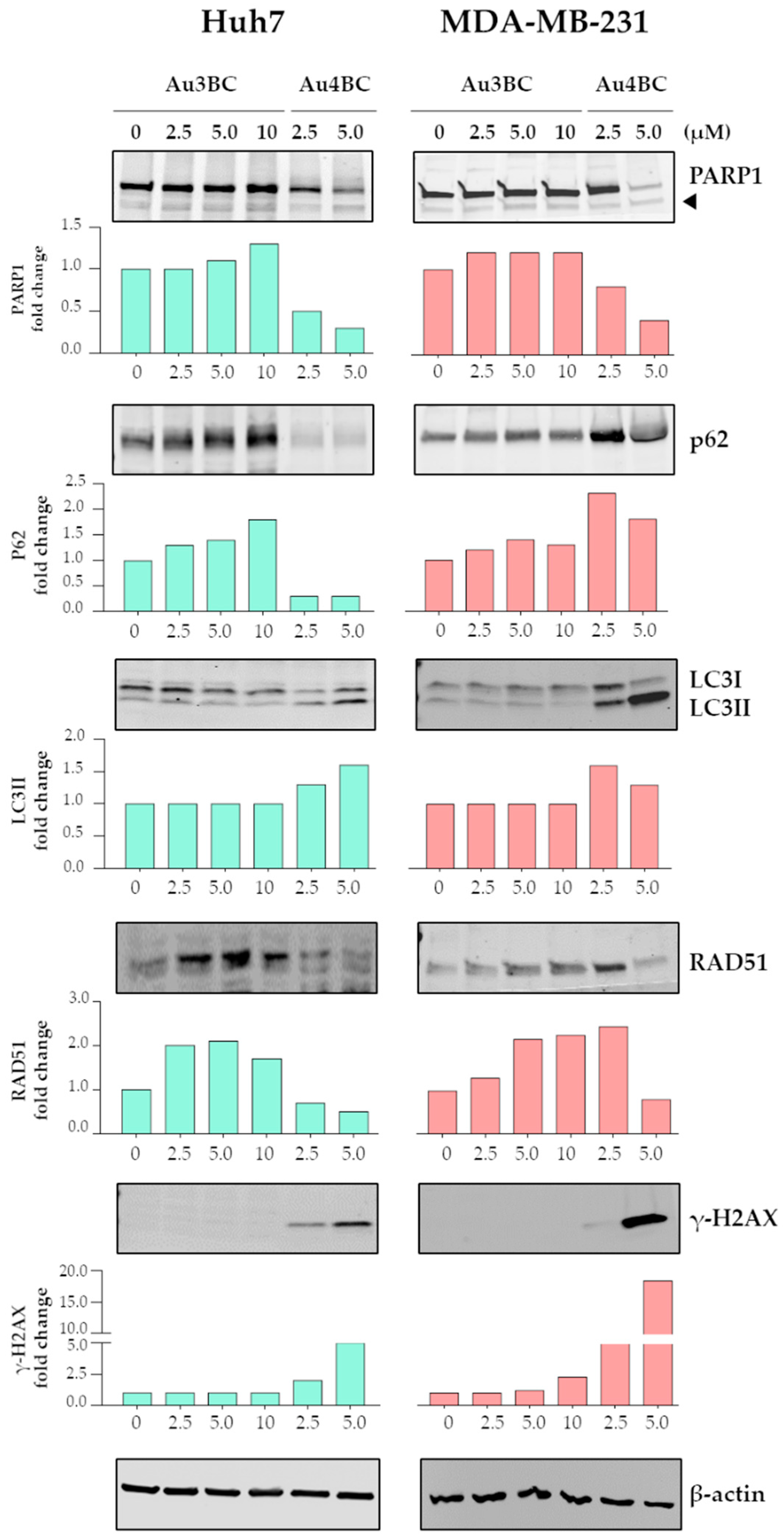
3.2.3. Effect of Au3BC and Au4BC on the Activation of the Apoptosis and Autophagy in HCC and BC Cells
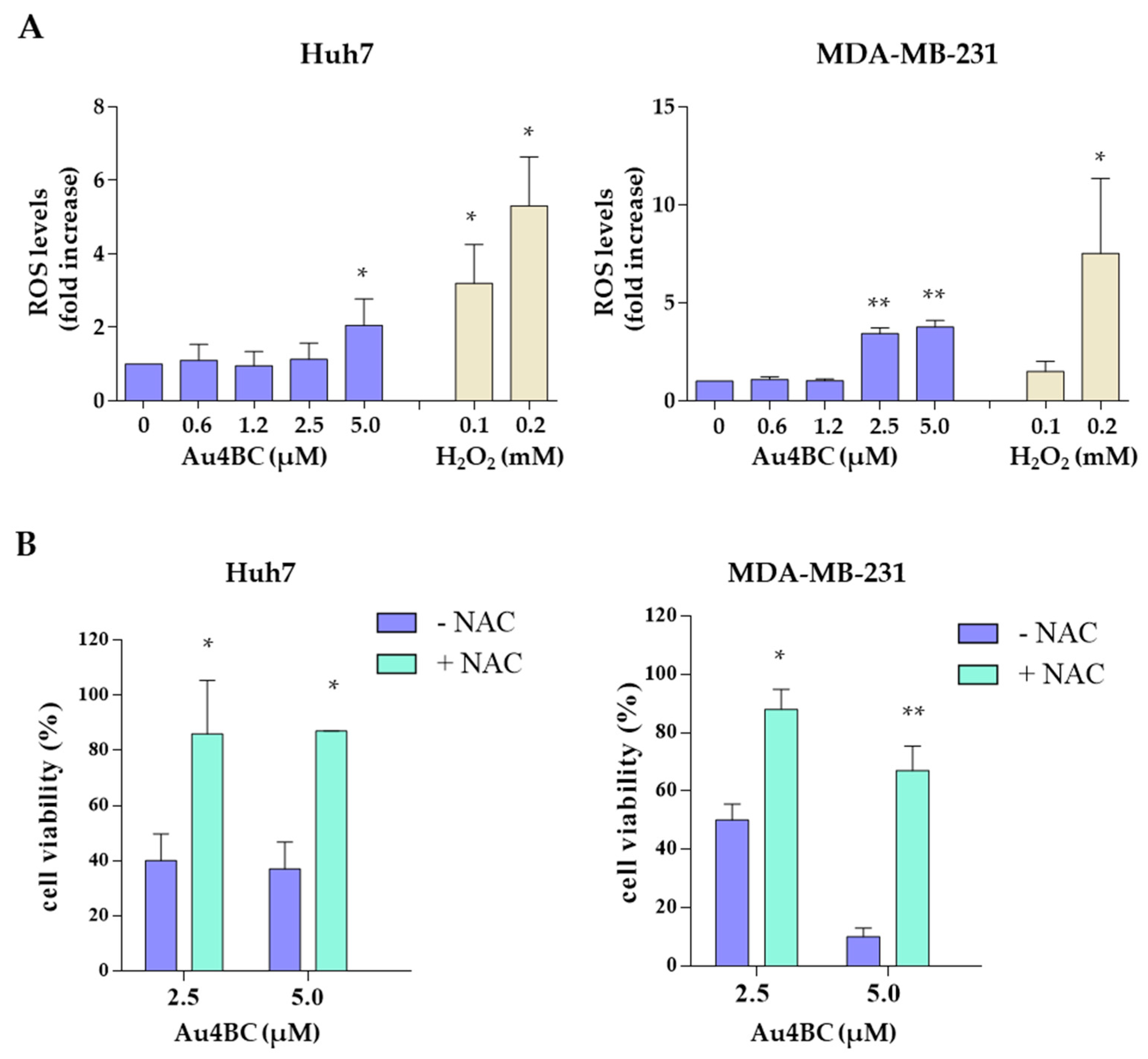
3.2.4. Au4BC Induced Intracellular Reactive Oxygen Species (ROS) Production
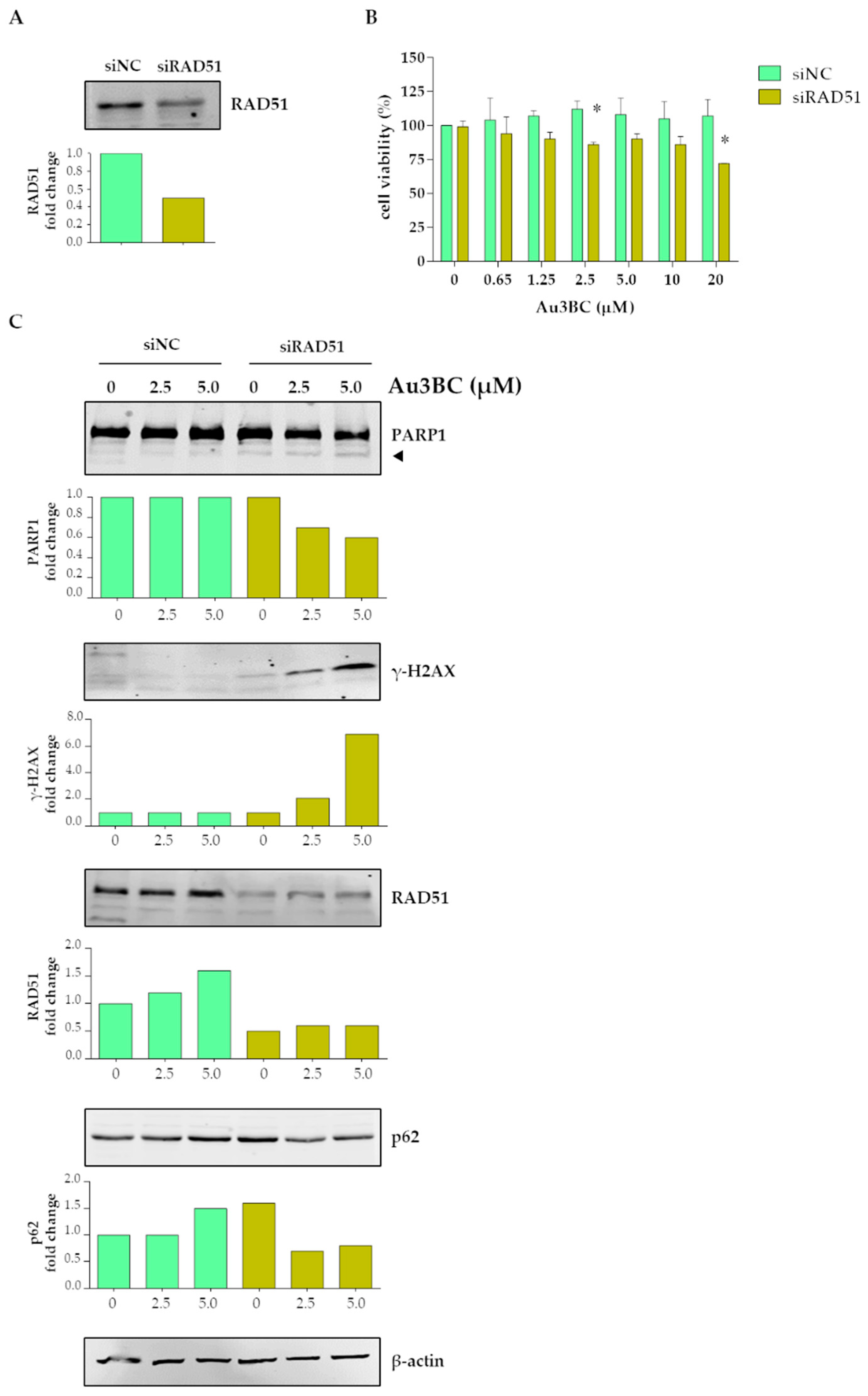
3.2.5. siRNA-Mediated Knockdown (KD) of RAD51 Gene Expression Potentiates the Sensitivity of Huh7 Cells to Au3BC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mjos, K.D.; Orvig, C. Metallodrugs in Medicinal Inorganic Chemistry. Chem. Rev. 2014, 114, 4540–4563. [Google Scholar] [CrossRef] [PubMed]
- Miranda, V.M. Medicinal inorganic chemistry: An updated review on the status of metallodrugs and prominent metallodrug candidates. Rev. Inorg. Chem. 2022, 42, 29–52. [Google Scholar] [CrossRef]
- Kelland, L. The resurgence of platinum-based cancer chemotherapy. Nat. Rev. Cancer 2007, 7, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Clarke, M.J.; Zhu, F.; Frasca, D.R. Non-platinum chemotherapeutic metallopharmaceuticals. Chem. Rev. 1999, 99, 2511–2533. [Google Scholar] [CrossRef] [PubMed]
- Komeda, S.; Casini, A. Next-generation anticancer metallodrugs. Curr. Top. Med. Chem. 2012, 12, 219–235. [Google Scholar] [CrossRef]
- Ott, I. On the medicinal chemistry of gold complexes as anticancer drugs. Coord. Chem. Rev. 2009, 253, 1670–1681. [Google Scholar] [CrossRef]
- Roder, C.; Thomson, M.J. Auranofin: Repurposing an Old Drug for a Golden New Age. Drugs R&D 2015, 15, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Available online: https://clinicaltrials.gov/ct2/show/NCT01419691 (accessed on 18 January 2016).
- Chiappetta, G.; Gamberi, T.; Faienza, F.; Limaj, X.; Rizza, S.; Messori, L.; Filomeni, G.; Modesti, A.; Vinh, J. Redox proteome analysis of auranofin exposed ovarian cancer cells (A2780). Redox Biol. 2022, 52, 102294. [Google Scholar] [CrossRef]
- Jovanović, M.; Podolski-Renić, A.; Krasavin, M.; Pešić, M. The Role of the Thioredoxin Detoxification System in Cancer Progression and Resistance. Front. Mol. Biosci. 2022, 9, 883297. [Google Scholar] [CrossRef]
- Fu, B.; Meng, W.; Zeng, X.; Zhao, H.; Liu, W.; Zhang, T. TXNRD1 Is an Unfavorable Prognostic Factor for Patients with Hepatocellular Carcinoma. Bio. Med. Res. Int. 2017, 2017, 4698167. [Google Scholar] [CrossRef]
- Lee, D.; Xu, I.M.; Chiu, D.K.; Leibold, J.; Tse, A.P.; Bao, M.H.; Yuen, V.W.; Chan, C.Y.; Lai, R.K.; Chin, D.W.; et al. Induction of Oxidative Stress Through Inhibition of Thioredoxin Reductase 1 Is an Effective Therapeutic Approach for Hepatocellular Carcinoma. Hepatology 2019, 69, 1768–1786. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhao, Y.; Wei, Z.; Tao, L.; Sheng, X.; Wang, S.; Chen, J.; Ruan, J.; Liu, Z.; Cao, Y.; et al. Targeting Thioredoxin System with an Organosulfur Compound, Diallyl Trisulfide (DATS), Attenuates Progression and Metastasis of Triple-Negative Breast Cancer (TNBC). Cell. Physiol. Biochem. 2018, 50, 1945–1963. [Google Scholar] [CrossRef]
- Bhatia, M.; McGrath, K.L.; Di Trapani, G.; Charoentong, P.; Shah, F.; King, M.M.; Clarke, F.M.; Tonissen, K.F. The thioredoxin system in breast cancer cell invasion and migration. Redox Biol. 2016, 8, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Gencheva, R.; Arnér, E. Thioredoxin Reductase Inhibition for Cancer Therapy. Annu. Rev. Pharmacol. Toxicol. 2022, 62, 177–196. [Google Scholar] [CrossRef] [PubMed]
- Karaca, Ö.; Scalcon, V.; Meier-Menches, S.M.; Bonsignore, R.; Brouwer, J.M.J.L.; Tonolo, F.; Folda, A.; Rigobello, M.P.; Kühn, F.E.; Casini, A. Characterization of Hydrophilic Gold(I) N-Heterocyclic Carbene (NHC) Complexes as Potent TrxR Inhibitors Using Biochemical and Mass Spectrometric Approaches. Inorg. Chem. 2017, 56, 14237–14250. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, C.; Albrecht, L.; Balasupramaniam, S.; Misgeld, R.; Karge, B.; Brönstrup, M.; Prokop, A.; Baumann, K.; Reichl, S.; Ott, I.A. A gold(I) biscarbene complex with improved activity as a TrxR inhibitor and cytotoxic drug: Comparative studies with different gold metallodrugs. Metallomics 2019, 11, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Rubbiani, R.; Salassa, L.; de Almeida, A.; Casini, A.; Ott, I. Cytotoxicgold(I) N-heterocyclic carbine complexes with phosphane ligands as potent enzyme inhibitors. Chem. Med. Chem. 2014, 9, 1205–1210. [Google Scholar] [CrossRef] [PubMed]
- Bazzicalupi, C.; Ferraroni, M.; Papi, F.; Massai, L.; Bertrand, R.; Messori, L.; Gratteri, P.; Casini, A. Determinants for Tight and Selective Binding of a Medicinal Dicarbene Gold(I) Complex to a Telomeric DNA G-Quadruplex: A Joint ESI MS and XRD Investigation. Angew. Chem. Int. Ed. 2016, 55, 4256–4259. [Google Scholar] [CrossRef]
- Wragg, D.; de Almeida, A.; Bonsignore, R.; Kuehn, F.E.; Leoni, S.; Casini, A. On the Mechanism of Gold/NHC Compounds Binding to DNA G Quadruplexes: Combined Metadynamics and Biophysical Methods. Angew. Chem. Int. Ed. 2018, 57, 14524–14528. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.H.; Shih, W.C.; Chang, H.C.; Kuo, Y.Y.; Hung, W.C.; Ong, T.G.; Li, S.W. Preparation and Characterization of Amino-Linked Heterocyclic Carbene Palladium, Gold, and Silver Complexes and Their Use as Anticancer Agents That Act by Triggering Apoptotic Cell Death. J. Med. Chem. 2011, 54, 5245–5249. [Google Scholar] [CrossRef]
- Concepción Gimeno, M.; Laguna, A.; Visbal, R.N. Heterocyclic Carbene Coinage Metal Complexes as Intense Blue-Green Emitters. Organometallics 2012, 31, 7146–7157. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Bensdorf, K.; Proetto, M.; Hagenbach, A.; Abram, U.; Gust, R. Synthesis, characterization, and in vitro studies of bis[1,3-diethyl-4,5-diarylimidazol-2-ylidene]gold(I/III) complexes. J. Med. Chem. 2012, 55, 3713–3724. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Bensdorf, K.; Hagenbach, A.; Abram, U.; Niu, B.; Mariappan, A.; Gust, R. Synthesis and biological studies of silver N-heterocyclic carbene complexes derived from 4,5-diarylimidazole. Eur. J. Med. Chem. 2011, 46, 5927–5934. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Bensdorf, K.; Proetto, M.; Abram, U.; Hagenbach, A.; Gust, R. NHC gold halide complexes derived from 4,5-diarylimidazoles: Synthesis, structural analysis, and pharmacological investigations as potential antitumor agents. J. Med. Chem. 2011, 54, 8605–8615. [Google Scholar] [CrossRef]
- Cusimano, A.; Puleio, R.; D’Alessandro, N.; Loria, G.R.; McCubrey, J.A.; Montalto, G.; Cervello, M. Cytotoxic activity of the novel small molecule AKT inhibitor SC66 in hepatocellular carcinoma cells. Oncotarget 2015, 6, 1707–1722. [Google Scholar] [CrossRef] [Green Version]
- Porchia, M.; Pellei, M.; Marinelli, M.; Tisato, F.; Del Bello, F.; Santini, C. New insights in Au-NHCs complexes as anticancer agents. Eur. J. Med. Chem. 2018, 146, 709–746. [Google Scholar] [CrossRef]
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Zucman-Rossi, J.; Finn, R.S. Hepatocellular carcinoma. Nature Rev. Dis. Primers 2021, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Smolarz, B.; Nowak, A.Z.; Romanowicz, H. Breast Cancer-Epidemiology, Classification, Pathogenesis and Treatment (Review of Literature). Cancers 2022, 14, 2569. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Guarra, F.; Pratesi, A.; Gabbiani, C.; Biver, T. A focus on the biological targets for coinage metal-NHCs as potential anticancer complexes. J. Inorg. Biochem. 2021, 217, 111355. [Google Scholar] [CrossRef] [PubMed]
- Mora, M.; Gimeno, M.C.; Visbal, R. Recent advances in gold-NHC complexes with biological properties. Chem. Soc. Rev. 2019, 48, 447–462. [Google Scholar] [CrossRef] [PubMed]
- Visbal, R.; Fernández-Moreira, V.; Marzo, I.; Laguna, A.; Gimeno, M.C. Cytotoxicity and biodistribution studies of luminescent Au(I) and Ag(I) N-heterocyclic carbenes. Searching for new biological targets Dalton Trans. 2016, 45, 15026–15033. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Augello, G.; Azzolina, A.; Rossi, F.; Prencipe, F.; Mangiatordi, G.F.; Saviano, M.; Ronga, L.; Cervello, M.; Tesauro, D. New Insights into the Behavior of NHC-Gold Complexes in Cancer Cells. Pharmaceutics 2023, 15, 466. https://doi.org/10.3390/pharmaceutics15020466
Augello G, Azzolina A, Rossi F, Prencipe F, Mangiatordi GF, Saviano M, Ronga L, Cervello M, Tesauro D. New Insights into the Behavior of NHC-Gold Complexes in Cancer Cells. Pharmaceutics. 2023; 15(2):466. https://doi.org/10.3390/pharmaceutics15020466
Chicago/Turabian StyleAugello, Giuseppa, Antonina Azzolina, Filomena Rossi, Filippo Prencipe, Giuseppe Felice Mangiatordi, Michele Saviano, Luisa Ronga, Melchiorre Cervello, and Diego Tesauro. 2023. "New Insights into the Behavior of NHC-Gold Complexes in Cancer Cells" Pharmaceutics 15, no. 2: 466. https://doi.org/10.3390/pharmaceutics15020466
APA StyleAugello, G., Azzolina, A., Rossi, F., Prencipe, F., Mangiatordi, G. F., Saviano, M., Ronga, L., Cervello, M., & Tesauro, D. (2023). New Insights into the Behavior of NHC-Gold Complexes in Cancer Cells. Pharmaceutics, 15(2), 466. https://doi.org/10.3390/pharmaceutics15020466