1. Introduction
Orchids (Orchidaceae Juss.) is one of the largest families of flowering plants, with 736 genera and more than 28,000 species [
1,
2]. The genus belongs to the subfamily Epidendroideae, tribe Arethuseae, subtribe Coelogyninae and is divided into 23 sections [
3]. According to different authors, it includes between 120 and 200 species [
3,
4,
5]. Representatives of the genus are distributed in Southeast Asia and the Pacific Islands, with centers of species diversity in the Himalayas and on the islands of Java and Kalimantan [
4]. Members of the genus show a wide range of ecological preferences and include epiphytes, lithophytes, and terrestrial species. The main habitats are humid montane tropical forests. Some species climb mountains up to 3300 m above sea level, while others grow near ocean coasts [
6].
An interesting feature of the genus is the structural and rhythmological diversity of shoots, with a synchronous development of the vegetative and generative parts or advanced development of the generative part of the vegetative-generative shoot [
6,
7]. Some Himalayan species from regions with alternating wet and dry cool periods develop intercalary (multiseasonal) inflorescences. The flowers are resupinant, and the inflorescence always turns lip-down. Some species of
Coelogyne with large flowers are commercially used, and new ornamental hybrids have been obtained [
8]. The medicinal properties of
Coelogyne are also in demand. The shoots and leaves of many species are used in traditional oriental and modern medicine. They contain the stibenoids coelogin, coeloginin, ochrone, ochrolic acid, and ochrolone [
3,
9]. Substances with bactericidal and fungicidal properties have been isolated from the leaves of
Coelogyne speciosa [
10,
11]. At present, methods are being developed for cultivating the
Coelogyne medicinal species in vitro [
12], including the method of incomplete somatic embryogenesis. However, prolonged fruit ripening (from 1.5 to 2.5 years) and the lack of information regarding the initial stages of ovule development affect the efficiency of the biotechnological work [
12].
The objectives of this study were to document, in detail, the megasporogenesis and megagametogenesis of the Coelogyne speciosa subsp. fimbriata (J.J.Sm.) Gravendeel and establish the time of fertilization and embryo development.
2. Materials and Methods
We used plants from the collection of the Fund Greenhouse of the N.V. Tsitsin Main Botanical Garden of Russian Academy of Sciences (Moscow, Russia). The plants were grown as epiphytes in pots and baskets with a substrate of bark and sphagnum under natural light in a greenhouse with a cold temperature regime (10–18 °C) and high relative air humidity (more than 60%). Flowering was observed between July and January, and the flowers were pollinated by the geitonogamous method. Artificial pollination was carried out between the flowers of different plants of the same clone as they bloomed (geitonogamous method). A total of 40 flowers were pollinated. The fruits were harvested in 2020–22, every 5 days, after pollination (DAP). A total of 25 fruits were collected.
Fragments of immature fruits of different ages were fixed in 2% paraformaldehyde in 0.05 M phosphate buffer pH 7.4 at 4 °C and stored until use (at least 1 week). The ovules were stained on a glass slide for 1–2 h in a humid chamber to prevent drying out. For staining, water solutions of each of the fluorescent dyes were used: 100 µg/mL berberine (BRB, Sigma-Aldrich, Burlington, MA, USA), 1–5 mg/mL Calcofluor (CF, Fluorescent Brighter 28, Sigma-Aldrich), or 50 µg/mL propidium iodide (PI, Sigma-Aldrich). The stained samples were washed several times with distilled water on a glass slide, placed in 50% (w/v) glycerol, covered with a cover glass, and stored at 4 °C until use (usually at least 1–2 weeks). Four replicates were examined at each step: three with fluorescent dyes and one unstained preparation (autofluorescence). A total of 100 preparations were made.
The samples were viewed under a confocal laser scanning microscope (Olympus FV1000D, Shinjuku, Tokyo, Japan). Lasers with wavelengths of 405 nm, 473 nm, and 560 nm were used, individually or in combination, according to the absorption spectra of the dies to excite fluorescence. The stained samples were examined using 5–20% laser intensity. The signal was registered in the blue (425–460 nm), green (485–530 nm), and red (560–660 nm) channels that corresponded to the settings of the channel parameters for DAPI (4.6-diamidino-2-phenylindole), Alexa Fluor 488, and rhodamine, respectively. The figures show the confocal fluorescence images either alone or merged with the differential interference contrast (DIC).
3. Results
The generative shoots of C. speciosa subsp. Fimbriata are up to 5–7 cm long, drooping, bears 2–4 large flowers. The bracts are sharp, ovate-oblong, and fall off simultaneously with the opening of the flowers. The flowers are resupinant (the lip of the flower is turned to the lower position), sessile, up to 8 cm in diameter, with greenish-yellow petals and sepals and a white three-lobed lip with chocolate-brown longitudinal ridges. The column is curved and possesses pterygoid processes in the distal part. The ovary is inferior, 3.4–3.5 cm long. The ovary is formed by three fused carpels that develop as six valves. Three fertile valves oppose the petals and form the placenta, and three sterile valves oppose the sepals. The pollinated ovary develops into a dehiscence fruit (capsule).
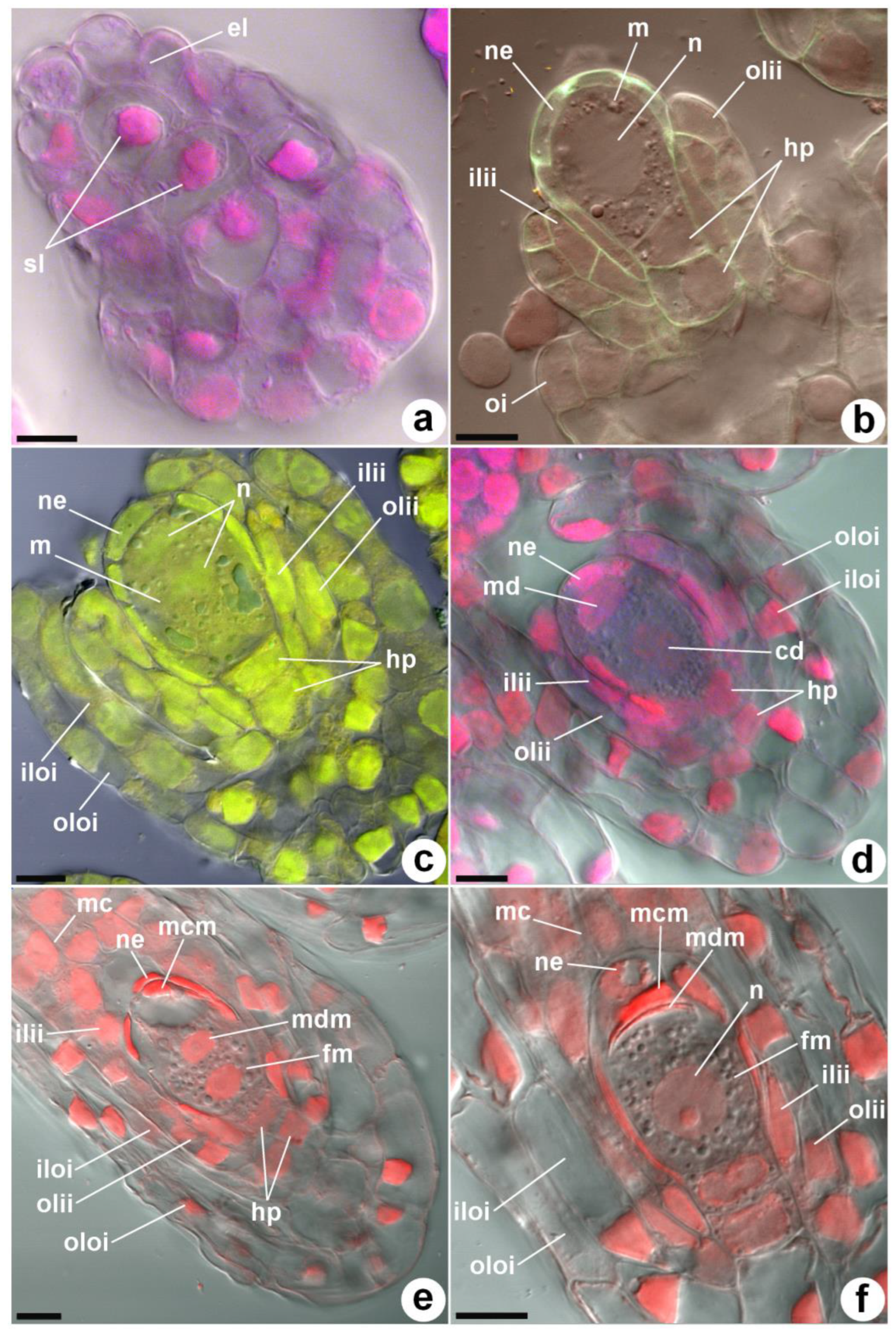
Before the pollination of the flower, the primordia of the ovule are already differentiated. After pollination, the ovule primordia on the placenta gradually lengthen and branch. The subepidermal axial row of cells (nucellar filament) in the ovule primordia is short and consists of three cells (
Figure 1a). The upper subepidermal cell increases in size and transforms into an archesporial cell. The two lower cells of the nucellar filament become hypostasis cells. At 45–50 days after pollination (DAP), the archesporial cells transform into megasporocytes with a denser cytoplasm and a large nucleus.
Figure 1b shows a large subepidermal archesporium cell and two hypostasis cells below it. The number of nucellus cells on a longitudinal section is six. The inner integument half surrounds the archesporium (megasporocyte) cell. In the epidermal layer, 2-cell initials of the external integument are visible.
The initials of the inner integument are laid first by periclinal divisions of one of the cells of the nucellus epidermis, and slightly later, the initials of the outer integument are laid. The growth of the ovule integument occurs in the same order: first, the inner integument lengthens; then, the outer integument lengthens. Both integuments, gradually lengthening, surround the megasporocyte, and the ovule assumes an anatropic position relative to the placenta. The inner and outer integuments of the ovule each consist of two layers of cells, with the exception of the 3-layer edge of the outer integument from the side of the funiculus.
After 58–60 DAP, the first meiotic division of the megasporocyte occurs (
Figure 1c), resulting in the formation of unequal megaspore dyads. The distal megaspore of the dyad dies off (
Figure 1d), and a brighter glow of its callous membranes is observed during confocal microscopy (
Figure 1e). Then, the chalazal megaspore undergoes a second meiotic division, and callose is also present in the cell wall of the second degenerating megaspore. At the stage of meiotic division, the integuments completely surround the nucellus of the ovule. Two meiotic divisions at the 63–65 DAP stage result in a linear triad, consisting of one functional chalazal megaspore and two degenerating micropylar megaspores (
Figure 1f). The nucellus and two hypostasis cells are preserved until the stage of the degeneration of the micropylar and median megaspores. As a result, a monosporic embryo sac is formed.
From 65–70 DAP, a female gametophyte is formed from a functional megaspore. The nucellus surrounding the megasporocyte begins to degenerate by the time of the first division of the functional megaspore nucleus (
Figure 2a). However, the lowest chalazal row of the nucellus epidermis, adjacent to the upper cell of the hypostasis, is preserved. The first two mitotic divisions form four nuclei that diverge in pairs towards the micropylar and chalazal ends of the embryo sac (
Figure 2b). The third mitotic division of each of these nuclei leads to the formation of 8-nuclear (
Figure 2d–f) embryo sacs, which make up the majority. In a smaller number of embryo sacs, the third mitotic division occurs in only one of the chalazal and micropylar nuclei, resulting in the formation of mature 6-nuclear embryo sacs, with a reduction in the nuclei at both the micropylar and chalazal ends (
Figure 2c). In most embryo sacs, a large vacuole is formed, which prevents the advancement and connection of the polar nuclei in the center of the embryo sac (
Figure 2c,d) or pushes the polar nuclei away from the walls of the chalazal zone, towards the center of the embryo sac.
The formation of mature embryo sacs was observed after 85 DAP (
Figure 2f). In 8-nuclear embryo sacs, one of the micropylar nuclei forms an egg, whilst the other two are converted into two synergids, and the fourth (upper polar nucleus) remains a free nucleus without a cell membrane. The chalazal complex of nuclei does not form cell membranes and contains one lower polar nucleus and three chalazal nuclei. The connection of the micropylar and chalazal polar nuclei can occur in different positions, relative to the large central vacuole. This usually occurs in the center of the embryo sac or closer to its micropylar half, but can sometimes occur in the chalazal part of the embryo sac. In some cases, the polar nuclei merges with the formation of a diploid central nucleus; in others, the polar nuclei simply approaches and remains connected, but does not merge. Endosperm formation does not occur. At 85–100 DAP, fertilization and an increase in the size of the ovules occur. At this stage, the epidermis of the nucellus is completely destroyed, and the inner layer of the inner integument becomes much thinner.
We found that the micropyle of the C. speciosa subsp. fimbriata is turned towards the placenta (anatropic), and the nucellus is eliminated at the stage of the mature embryo sac (tenuinucellate). The micropylar and chalazal ends of the embryo sac have an equal number of nuclei (bitegmal), and the funiculus is formed (funicular). The embryo sac is monosporic, Polygonum-type, and 8-nuclear, with a normal distribution of nuclei (3 + 2 + 3) or 6-nuclear with a reduction in the nuclei in the chalazal and micropylar ends of the embryo sac (3 + 3). Endosperm formation was not detected.
4. Discussion
Orchids are characterized by the morphogenetic transformations of placental structures, megasporangium and female gametophytes, associated with mycosymbiotrophic ways of life. A number of embryonic features distinguish them from other flowering plants. A reduction in some of the stages of female gametophyte formation includes a decrease in the megasporangium mass, the absence or underdevelopment of the endosperm [
13], the loss of conductive tissue in the funiculus [
14], the simplification of the integument structure [
15], and the absence of cotyledons and germinal roots [
16].
Most orchid species have a three-carpel unilocular paracarp ovary; the placental ridges of the ovary are usually activated only after flower pollination [
17,
18,
19]. The ovule primordium begins to develop after the first periclinal division of one of the subepidermal cells of the placenta. The upper two cells become archesporial, and from the lower cell, by several more periclinal divisions, an axial row of cells is formed. Thus, the cells of the axial row differentiate later than the archesporial cells [
20].
In orchids, the development of the axial cell row has primarily been studied in temperate species [
20,
21,
22]; less attention has been given to tropical species. An interesting study is the work of Ochora [
23], in which the number of cells of the axial row (nucellar filament) was calculated in various orchid species from tropical Africa. The author found that the shortest axial row possessed seven cells (tribes Cymbidieae, Malaxideae and Vandeae, subtribe Disinae Sw.). In some monopodial growing orchids, as well as in the subtribe Polystachyiinae and in
Ansellia africana Lindl., it included nine cells. Thirteen cells were found in
Vanilla polylepis Summerh. In the primitive terrestrial orchid species
Neuwiedia veratrifolia Blume, six cells were found in the axial row [
24]. According to our data, in
C. speciosa subsp.
fimbriata, the axial row consists of only three cells, the upper of which becomes an archesporial cell.
The question of the archesporium transformation into the mother cell of megaspores was previously discussed in the embryological literature [
15,
23]. Some authors believe that orchids bypass the archesporium stage, and the subepidermal cell of the axial row immediately becomes a megasporocyte [
17,
23]. However, most researchers believe that the subepidermal cell of the orchid nucellus first turns into an archesporial cell, which is accompanied by an increase in size and physiological transformations, and only then into the mother cell of megaspores [
15].
The product of two successive meiotic divisions is a structure called the megaspore tetrad. Of the four megaspore cells, only one becomes functioning, and the rest die off. In different families of flowering plants, chalazal, micropylar, subchalazal, or even any of the four megaspores, can transform into the embryo sac [
25]. In the orchid family, in addition to tetrads, the formation of megaspore triads was also found. Triad formation occurs in representatives of many orchid genera, such as
Dendrobium Sw. [
26],
Calanthe R. Br. [
15], and
Cymbidium Sw. [
27]. The formation of megaspore triads is due to the suppression of cytokinesis in the micropylar cell of the dyad. In this case, meiosis II occurs only in the chalazal cell of the dyad. The chalazal daughter cell of this division becomes a functional megaspore. Degenerating micropylar megaspores probably prevents the premature penetration of the pollen tubes into the ovule, which is not yet ready for fertilization. Their degeneration occurs simultaneously with the mitotic divisions of the nuclei of the embryo sac and the elimination of the micropylar part of the nucellus epidermis, which opens the entrance to the embryo sac for the pollen tubes.
The orchid ovule is traditionally defined as tenuinucellate, i.e., the nucellus consists only of the epidermis surrounding the megaspore mother cell [
28]. However, in some orchids, crassinucellate ovules with a lateral region of the nucellus (the nucellus is massive, multilayered, and long-lasting) were also found [
20,
29]. Shamrov [
20] proposed distinguishing one more type of ovule—medionucellate—which is characterized by the preservation of the podium (nucellar column) and part of the nucellus epidermis at the stage of the female gametophyte and the early stages of embryogenesis. The two cells underlying the archesporium, and then the megasporocyte, are not surrounded by the nucellus epidermis; therefore, we consider them to be hypostasis cells. Thus, we did not find a podium in the ovule of the
C. speciosa subsp.
fimbriata. At the stage of megasporogenesis, the hypostasis cells are topographically well expressed, and they have larger nuclei than the other cells of the embryo sac membranes. The absence of a podium in the nucellus, as well as the relatively short (up to the stage of early embryogenesis) existence of the chalazal cells of the nucellus, makes it possible to consider the ovule of the
C. speciosa subsp.
fimbriata tenuinucellate. Histochemical studies have shown that the cells of the hypostasis and epidermis of the orchid nucellus are physiologically active tissues with a significant content of proteins and dextrins, particularly during megagametogenesis [
20]. For example, the hypostasis cells of
Cymbidium sinense Willd. are positive with periodic acid-Schiff staining [
30].
A reduction in the number of nuclei in the orchid embryo sacs has been repeatedly observed [
20,
28,
31]. Usually, as a result of three mitoses of a megaspore, a bipolar 8-nuclear embryo sac is formed. With the reduction in one of the mitotic divisions, 6–7-nuclear embryo sacs are formed. Yeung and Law [
32] suggested that a decrease in the number of nuclei within the orchid embryo sac could be caused by abnormalities in the mitosis, the fusion of the mitotic spindles, or the degeneration of the resulting chalazal nuclei. In the only previous study of megasporogenesis in the genus
Coelogyne [
33], in the embryo sacs of
C. tomentosa Lindl. (synonym
C. massangeana Rchb.f.), eight nuclei were found. We usually observed the development of the
C. speciosa subsp.
fimbriata according to the classical scheme, with the formation of a bipolar 8-nuclear embryo sac. However, sometimes a reduction in the third mitotic division was observed, and 6-nuclear embryo sacs were formed, with a decrease in the number of nuclei in the chalazal and/or micropylar zones.
Technologies for seed reproduction in vitro, cryopreservation and the indirect somatic embryogenesis of orchids require the rational selection of plant material, taking into account the critical periods of ovule formation, the duration of the female gametophyte development and the timing of the onset of embryogenesis. We found that the opening of the fruits of the
C. speciosa subsp.
fimbriata occurs between 600 and 750 DAP [
34]. Megasporogenesis occurs after 65–70 DAP and is characterized by the suppression of cytokinesis in the chalazal cell of the macrospore dyad at the stage of meiosis II. The formation of mature normal monosporic 8-nuclear or reduced 6-nuclear embryo sacs and the subsequent fertilization occurs after 85–100 DAP. Orchid embryos can be successfully cultivated in vitro, not only in the mature state, but also at the stage of the formation of several cells and even a zygote [
15]. In the case of the
C. speciosa subsp.
fimbriata, the fruit maturation time for in vitro culture can be reduced from 600–750 to 85–100 DAP.
5. Conclusions
This study sheds light on some of the details of the propagation and hybridization of Coelogyne, the study of which was previously hindered by the difficulty of artificial pollination, the long period of fruit ripening, and the slow growth of seedlings. It seems important to elucidate these features due to the lack of information regarding the initial stages of Coelogyne ovule development. Thus, the study of megasporegenesis and megagametogenesis expands the knowledge that is of fundamental and practical importance.
The following features of megasporogenesis and megagametogenesis were observed in the C. speciosa subsp. fimbriata:
1. The axial row consists of 3 cells, the upper subepidermal cell becomes archesporium, and the two lower subepidermal cells become hypostasis.
2. The structure of the ovules is anatropic, the micropyle faces the placenta, and the ovule turns before meiosis I.
3. Meiosis II occurs only in the chalazal cell of the megaspore dyad. As a result, a triad is formed, and not a tetrad of megaspores.
4. In the tenuinucellate ovule without a podium, the chalazal row of cells of the nucellus epidermis is preserved until the stage of early embryogenesis.
5. The ovule is bitegmal; the integuments consist of two-layer inner and outer integuments. On the side of the funiculus, the outer integument consists of three layers. The inner integument is preserved until the stage of early embryogenesis.
6. The embryo sac is monosporic, 6- or 8-nuclear, uni- or bi-polar, Polygonum-type, and often has a large vacuole that prevents the polar nuclei from approaching.
7. The polar nuclei in the embryo sac join or fuse, but no endosperm is formed.
8. The main stages of megasporogenesis and megagametogenesis were the formation of a functional megaspore after 65–70 DAP and the formation of a mature embryo sac and fertilization after 85–100 DAP.







{kind=link}
{kind=link}

