Ultramicrobacteria from Nitrate- and Radionuclide-Contaminated Groundwater

 , ,
, ,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling of Groundwater
2.2. Microbial Enumeration and Isolation
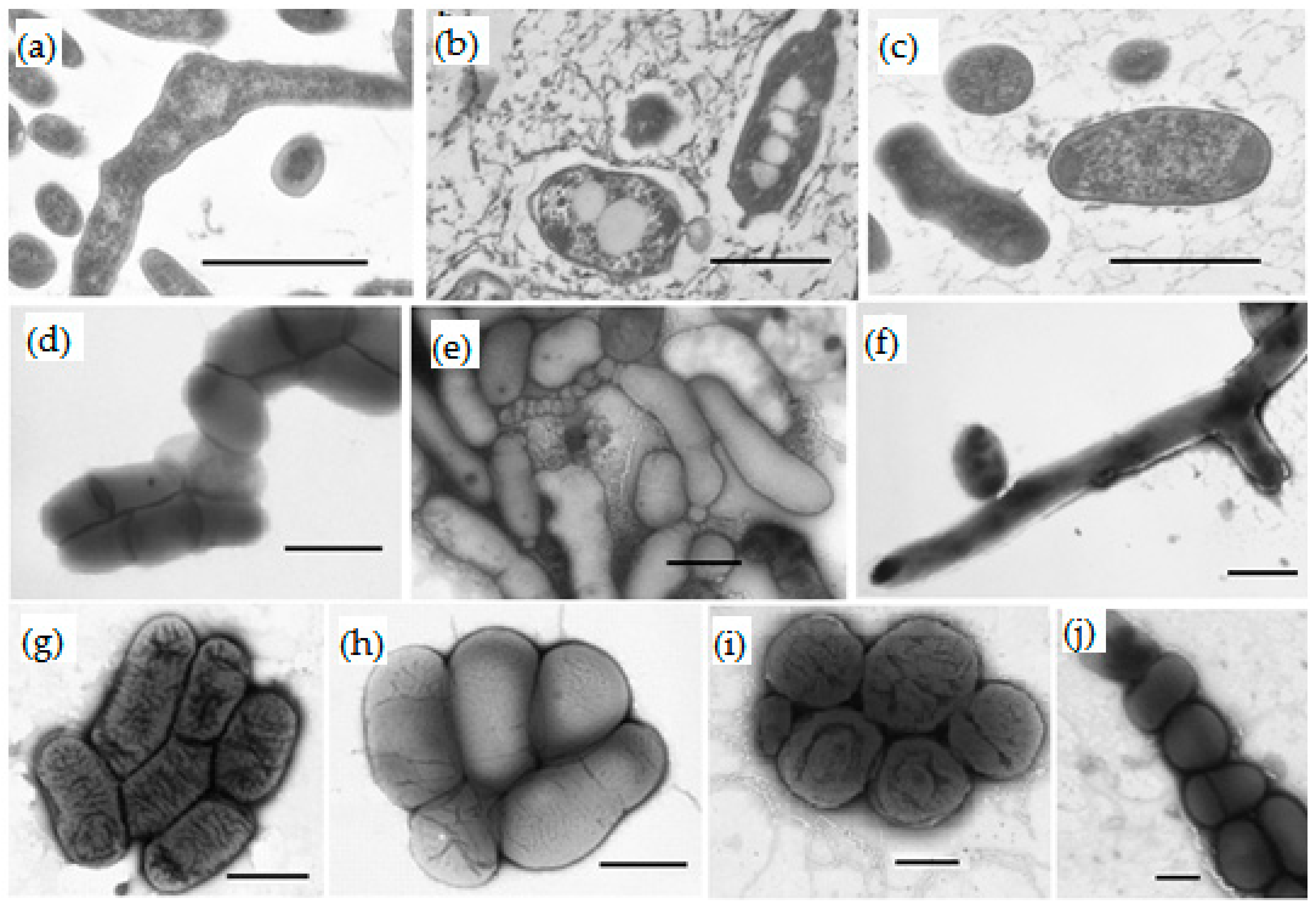
2.3. Microscopy
2.4. Identification of Pure Cultures
2.5. Analytical Methods
2.6. Radiotracer Methods and Tolerance to Heavy Metals and Ionizing Radiation
2.7. Construction and Analysis of the 16S rRNA Gene Clone Libraries
2.8. Genome Analysis
2.9. Nucleotide Sequence Accession Numbers
3. Results
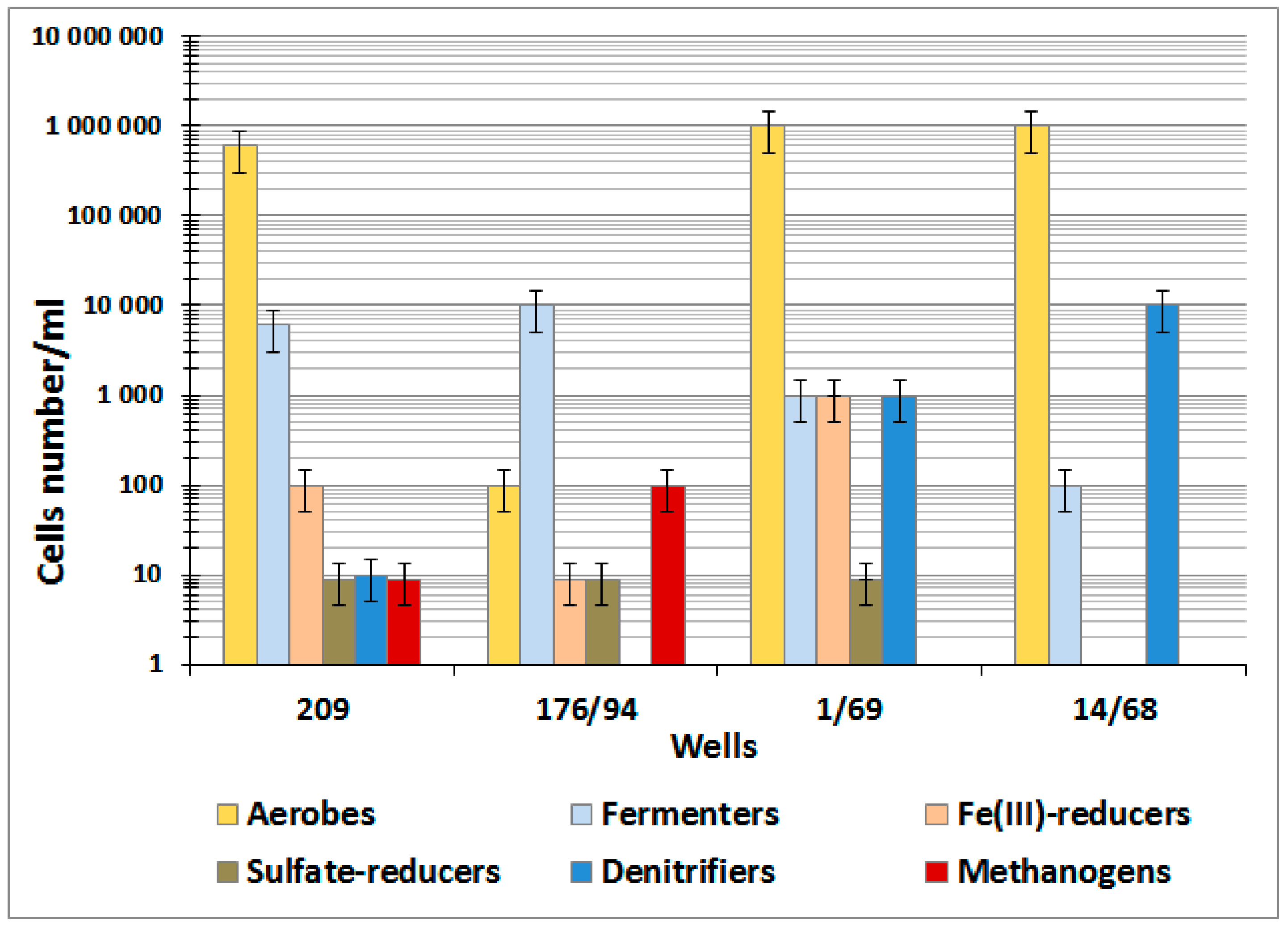
3.1. Physicochemical, Radiochemical, and Microbiological Characteristics of Groundwater
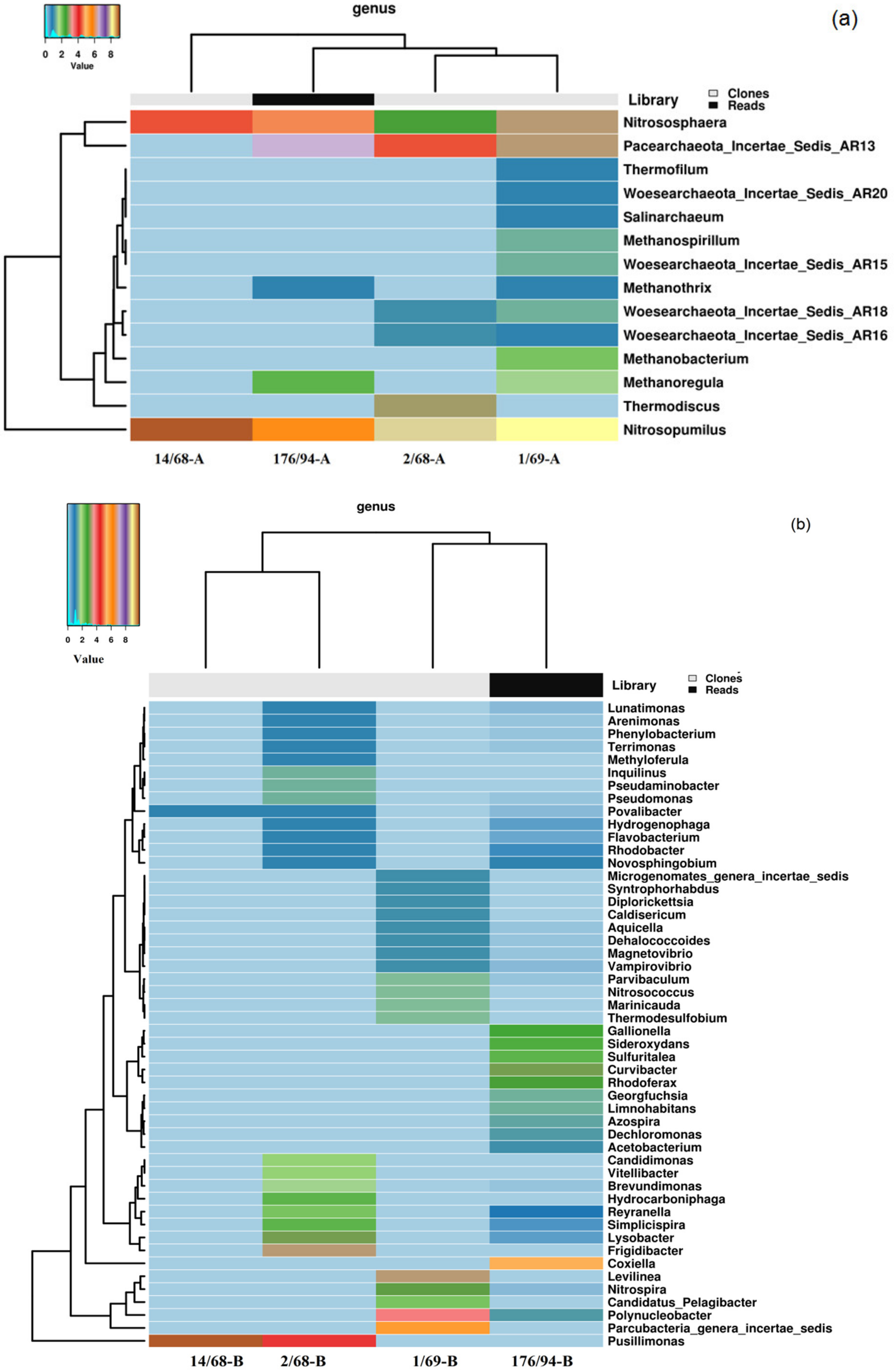
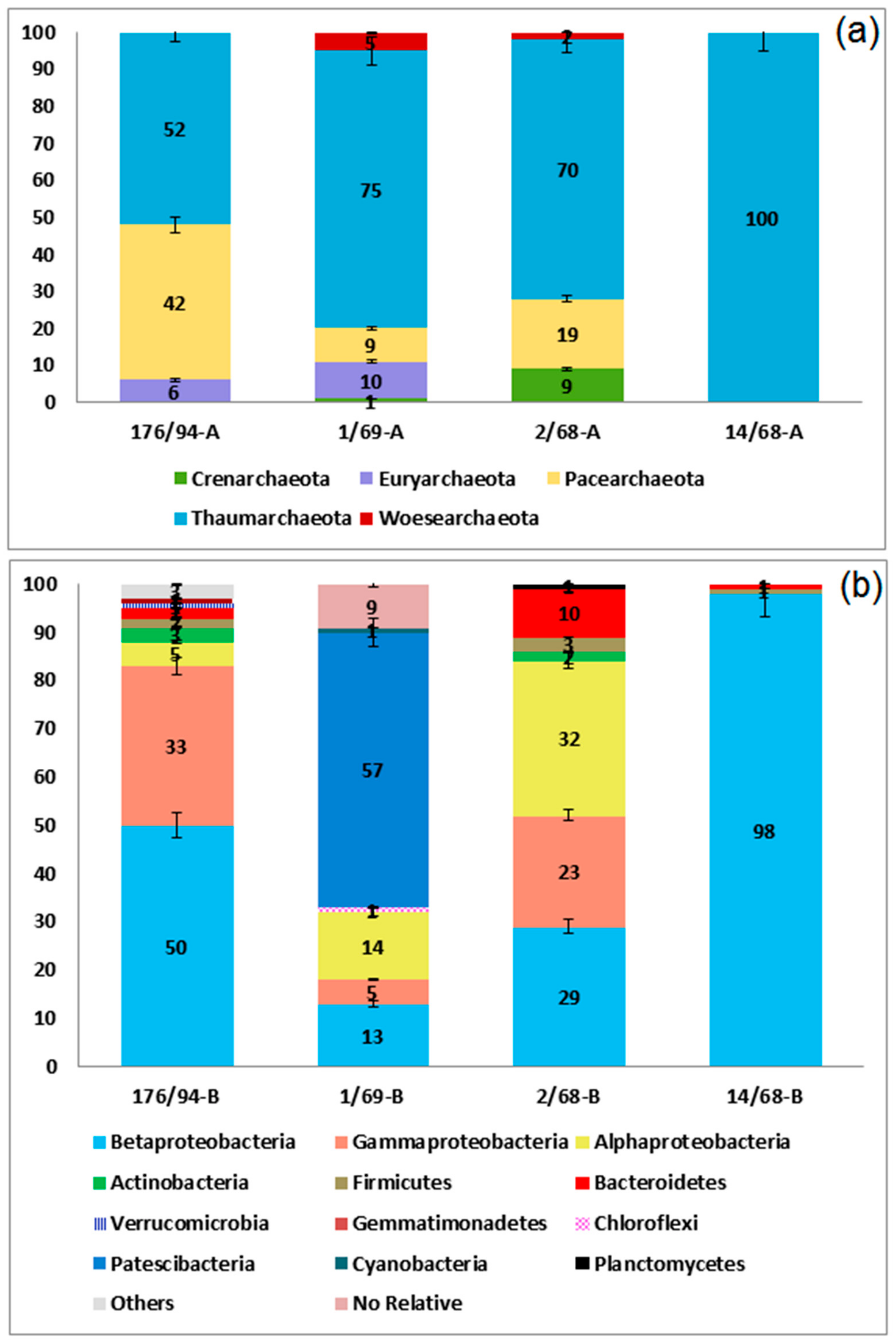
3.2. Phylogenetic Diversity of Archaea and Bacteria in Groundwater Samples
3.3. Isolation and Taxonomy of Bacteria from Groundwater
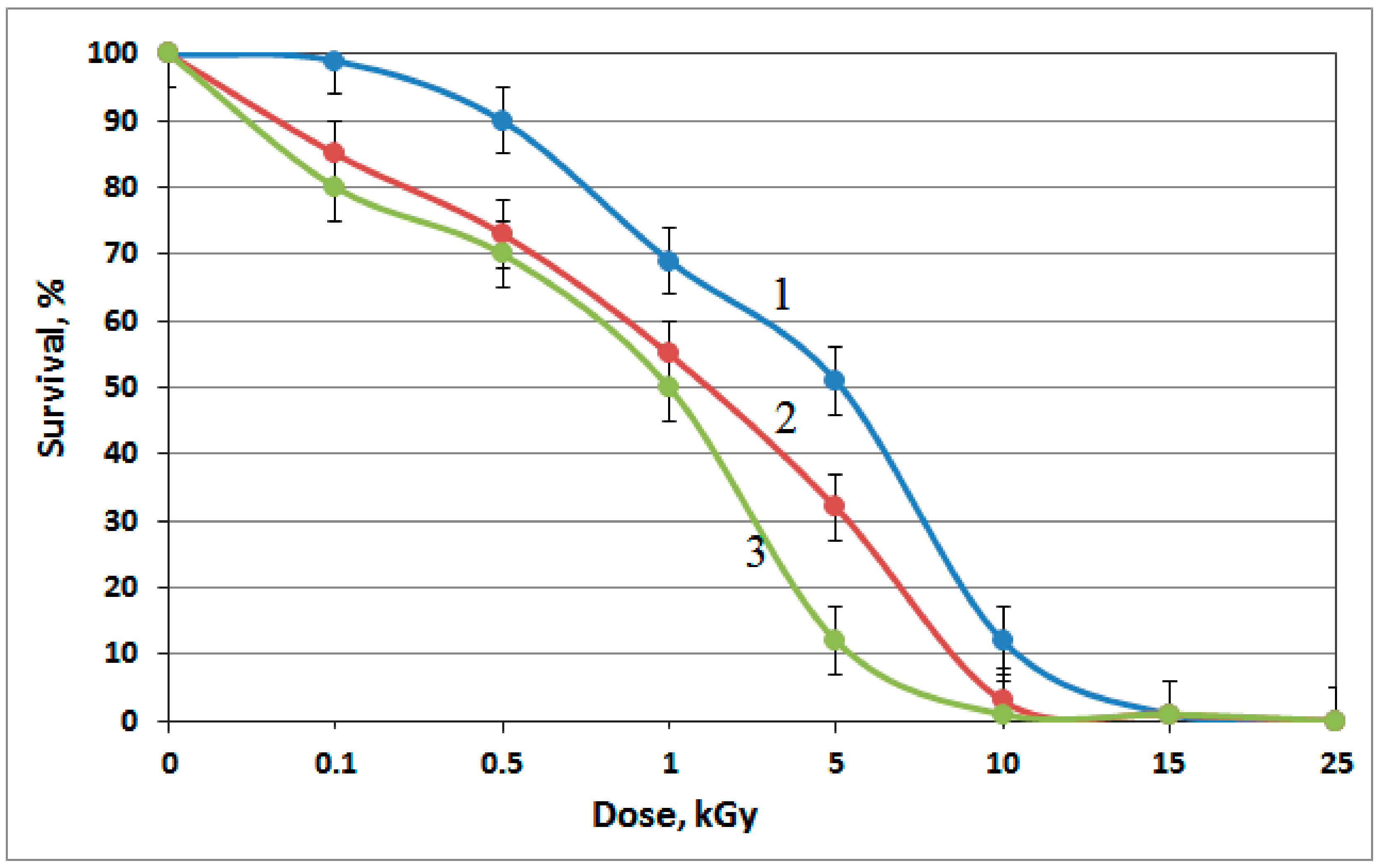
3.4. Tolerance of the Isolates to Heavy Metals and Ionizing Radiation
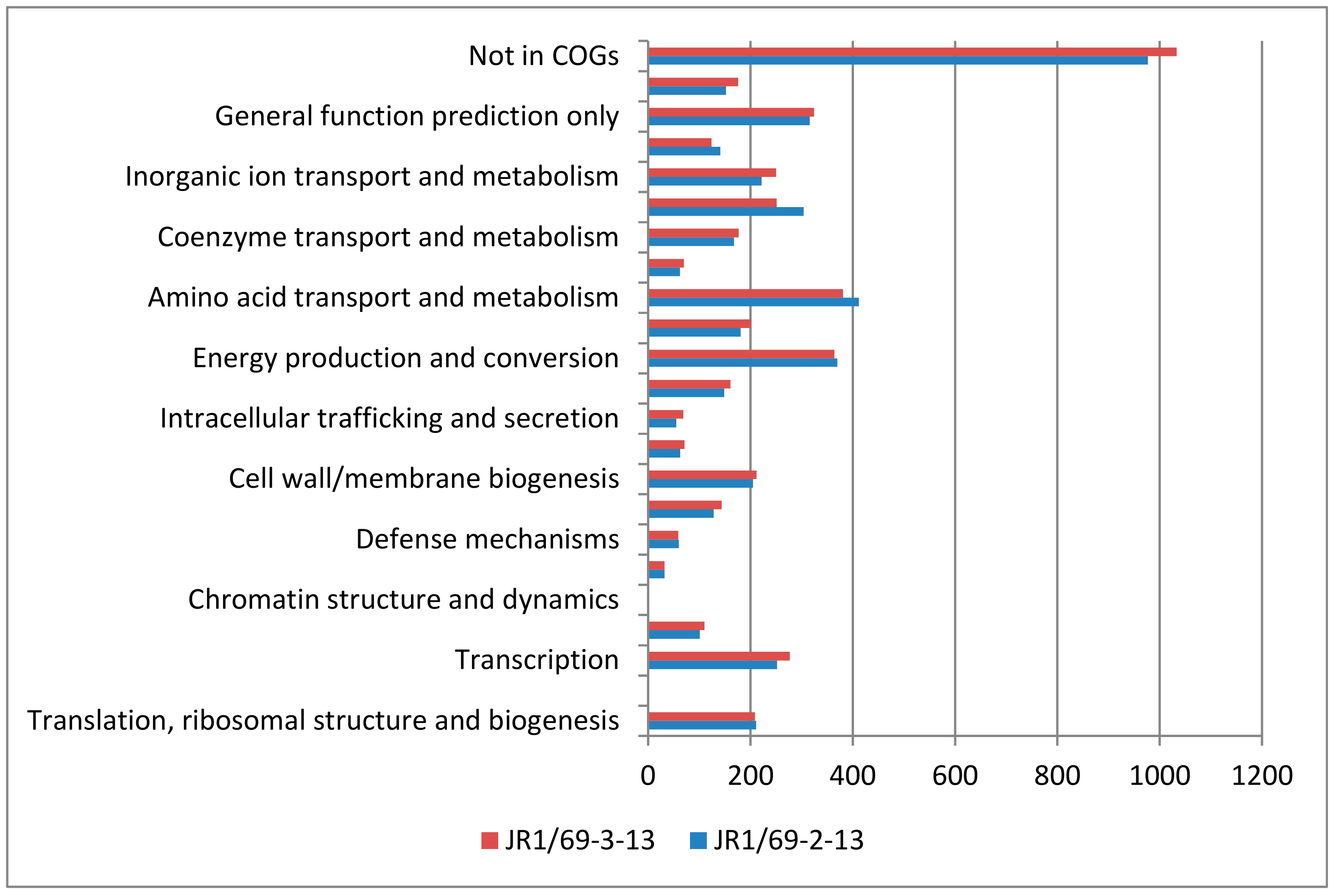
3.5. Metabolic Potential of Pusillimonas and Roseomonas Strains, as Assessed by Physiology and Genomes Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Super-Kingdom | Phylum, Class | Genus | Share of the Number of Sequences in the Library * from Well, % | |||
|---|---|---|---|---|---|---|
| 176/94 | 2/68 | 1/69 | 14/68 | |||
| Archaea | Crenarchaeota | Thermodiscus | 0 | 8.7 | 0 | 0 |
| Euryarchaeota | 5.8 | 0 | 9.7 | 0 | ||
| Methanobacterium | 0 | 0 | 4.0 | 0 | ||
| Methanoregula | 4.8 | 0 | 2.4 | 0 | ||
| Methanospirillum | 0 | 0 | 1.6 | 0 | ||
| Methanothrix | 1.0 | 0 | 0 | 0 | ||
| Pacearchaeota | Incertae_Sedis_AR13 | 42.3 | 19.4 | 8.9 | 0 | |
| Thaumarchaeota | 51.9 | 69.9 | 75.8 | 100 | ||
| Nitrosopumilus | 30.7 | 63.4 | 66.9 | 81.0 | ||
| Nitrososphaera | 21.2 | 6.5 | 8.9 | 19.0 | ||
| Woesearchaeota | 0 | 1.1 | 4.8 | 0 | ||
| Incertae_Sedis_AR15 | 0 | 0 | 2.4 | 0 | ||
| Incertae_Sedis_AR18 | 0 | 1.1 | 1.6 | 0 | ||
| Bacteria | Proteobacteria | 90.7 | 84.0 | 35.2 | 97.1 | |
| Alphaproteobacteria | 5.2 | 32.0 | 15.4 | 0 | ||
| Novosphingobium | 1.1 | 0 | 0 | 0 | ||
| Reyranella | 0 | 5.1 | 0 | 0 | ||
| Brevundimonas | 0 | 3.0 | 0 | 0 | ||
| Parvibaculum | 0 | 0 | 2.4 | 0 | ||
| Candidatus Pelagibacter | 0 | 0 | 4.8 | 0 | ||
| Marinicauda | 0 | 0 | 2.4 | 0 | ||
| Frigidibacter | 0 | 11.1 | 0 | 0 | ||
| Inquilinus | 0 | 2.0 | 0 | 0 | ||
| Pseudaminobacter | 0 | 2.0 | 0 | 0 | ||
| Betaproteobacteria | 51.5 | 29.0 | 14.3 | 97.1 | ||
| Gallionella | 6.9 | 0 | 0 | 0 | ||
| Curvibacter | 9.2 | 0 | 0 | 0 | ||
| Rhodoferax | 7.9 | 0 | 0 | 0 | ||
| Sulfuritalea | 5.6 | 0 | 0 | 0 | ||
| Limnohabitans | 2.0 | 0 | 0 | 0 | ||
| Dechloromonas | 1.6 | 0 | 0 | 0 | ||
| Polynucleobacter | 1.5 | 0 | 14.3 | 0 | ||
| Sideroxydans | 6.4 | 0 | 0 | 0 | ||
| Simplicispira | 0 | 6.1 | 0 | 0 | ||
| Pusillimonas | 0 | 18.2 | 0 | 97.1 | ||
| Candidimonas | 0 | 4.0 | 0 | 0 | ||
| Georgfuchsia | 2.1 | 0 | 0 | 0 | ||
| Azospira | 1.8 | 0 | 0 | 0 | ||
| Gammaproteobacteria | 34.0 | 23.0 | 5.5 | 0 | ||
| Uncultured Coxiellaceae | 30.9 | 0 | 0 | 0 | ||
| Pseudomonas | 0 | 2.0 | 0 | 0 | ||
| Nitrosococcus | 0 | 0 | 2.4 | 0 | ||
| Lysobacter | 0 | 9.1 | 0 | 0 | ||
| Hydrocarboniphaga | 0 | 6.1 | 0 | 0 | ||
| Nitrospirae | Nitrospira | 0 | 0 | 8.3 | 0 | |
| Bacteroidetes | 2.1 | 10.0 | 0 | 0 | ||
| Vitellibacter | 0 | 4.0 | 0 | 0 | ||
| Parcubacteria | Incertae_sedis | 0 | 0 | 34.5 | 0 | |
| Chloroflexi | Levilinea | 0 | 0 | 10.7 | 0 | |
| Firmicutes | 1.4 | 0 | 3.5 | 0 | ||
| Thermodesulfobium | 0 | 0 | 2.4 | 0 | ||
| Acetobacterium | 1.3 | 0 | 0 | 0 | ||
| Gemmatimonadetes | Gemmatimonas | 1.0 | 0 | 0 | 0 | |
| Parameter | Libraries from Observation Wells | |||||||
|---|---|---|---|---|---|---|---|---|
| 176/94 | 1/69 * | 2/68 | 14/68 | |||||
| Year of Sampling | 2012 | 2011 | 2010 | 2010 | ||||
| Depth of Sampling, m | 60 | 44 | 80 | 80 | ||||
| Library | 176/94-A | 176/94-B | 1/69-A | 1/69-B | 2/68-A | 2/68-B | 14/68-A | 14/68-B |
| Number of sequences in the library | 104 | 17,685 | 104 | 57 | 93 | 99 | 84 | 103 |
| Number of OTUs | 15 | 927 | 14 | 16 | 21 | 60 | 5 | 4 |
| Shannon-Weaver diversity index (H) | 2.21 | 5.08 | 1.27 | 2.58 | 2.54 | 3.65 | 0.85 | 0.16 |
| Simpson’s diversity index (1/S) | 6.11 | 35.50 | 1.97 | 11.0 | 9.0 | 20.80 | 1.70 | 1.10 |
| Evenness | 0.82 | 0.74 | 0.92 | 0.92 | 0.83 | 0.89 | 0.53 | 0.12 |
| Berger-Parker Dominance index (D) | 0.32 | 0.13 | 0.70 | 0.21 | 0.23 | 0.15 | 0.75 | 0.97 |
| Gene | Locus Tag | GenBank Identifier (pBLAST Best Hit) | % Identity | ||
|---|---|---|---|---|---|
| JR1/69-2-13 | JR1/69-3-13 | JR1/69-2-13 | JR1/69-3-13 | ||
| narG | - | CR159_07195 | WP_073102996.1 Bordetella petrii | - | 91 |
| narH | - | CR159_07190 | WP_008166145.1 Achromobacter arsenitoxydans | - | 91 |
| narI | - | CR159_07185 | WP_084658162.1 Chitinimonas taiwanensis | - | 72 |
| narJ | - | CR159_07180 | WP_050873229.1 Comamonas testosteroni | - | 81 |
| narX | CR159_07165 | WP_088588207.1 Achromobacter marplatensis | - | 64 | |
| narL | CR159_07160 | WP_076411578.1 Achromobacter xylosoxidans | - | 77 | |
| narK1 | CR159_07205 | WP_072427369.1 Chitinimonas taiwanensis | - | 80 | |
| narK2 | CR159_07200 | WP_028353905.1 Bordetella petrii | - | 80 | |
| nirK | CR155_06925 | CR159_13410 | WP_084135858.1 Candidimonas bauzanensis | 88 | 87 |
| nirK | CR155_06925 | CR159_19240 | WP_073101649.1 Candidimonas bauzanensis | 87 | 88 |
| norC | CR155_05890 | CR159_19250 | WP_073101466.1 Candidimonas bauzanensis | 82 | 84 |
| norB | CR155_05895 | CR159_19255 | WP_084135643.1 Candidimonas bauzanensis | 91 | 90 |
| norQ | CR155_05900 | CR159_19256 | WP_084135672.1 Candidimonas bauzanensis | 85 | 86 |
| norD | CR155_05905 | CR159_19257 | WP_073101467.1 Candidimonas bauzanensis | 74 | 76 |
| norZ | CR155_19235 | CR159_02315 | WP_073106495.1 Candidimonas bauzanensis | 91 | 90 |
| norA | CR155_19230 | CR159_02310 | WP_073106498.1 Candidimonas bauzanensis | 83 | 84 |
| norR | CR155_19225 | CR159_02305 | WP_073106501.1 Candidimonas bauzanensis | 89 | 89 |
| nosR | CR155_08950 | CR159_05710 | WP_073103310.1 Candidimonas bauzanensis | 80 | 81 |
| nosZ | CR155_08945 | CR159_05705 | WP_088147579.1 Achromobacter denitrificans | 89 | 90 |
| nosD | CR155_08940 | CR159_05700 | WP_073103312.1 Candidimonas bauzanensis | 79 | 80 |
| nosF | CR155_08935 | CR159_05695 | WP_073103313.1 Candidimonas bauzanensis | 84 | 83 |
| nosY | CR155_08930 | CR159_05690 | WP_073103314.1 Candidimonas bauzanensis | 89 | 88 |
| nosL | CR155_08925 | CR159_05685 | WP_084135995.1 Candidimonas bauzanensis | 72 | 66 |
| nosX | CR155_08920 | CR159_05680 | WP_073103316.1 Candidimonas bauzanensis | 66 | 68 |




References
- Morita, R.Y. Starvation and miniaturisation of heterotrophs, with special emphasis on maintenance of the starved viable state. In Bacteria in Their Natural Environment; Fletcher, M., Floodgate, G.D., Eds.; Academic Press: London, UK, 1985; pp. 111–130. [Google Scholar]
- Duda, V.I. Ultramicrobacteria. In Els; John Wiley & Sons Ltd.: Chichester, UK, 2011; pp. 1–15. [Google Scholar] [CrossRef]
- Schut, F.; Prins, R.A.; Gottschal, J.C. Oligotrophy and pelagic marine bacteria: Facts and fiction. Aquat. Microb. Ecol. 1997, 12, 177–202. [Google Scholar] [CrossRef] [Green Version]
- Ghiorse, W.C.; Wilson, J.T. Microbial ecology of the terrestrial subsurface. Adv. Appl. Microbiol. 1988, 33, 107–172. [Google Scholar] [PubMed]
- Wrighton, K.C.; Thomas, B.C.; Sharon, I.; Miller, C.S.; Castelle, C.J.; VerBerkmoes, N.C.; Wilkins, M.J.; Hettich, R.; Lipton, M.S.; Williams, K.; et al. Fermentation, hydrogen, and sulfur metabolism in multiple uncultivated bacterial phyla. Science 2012, 337, 1661–1665. [Google Scholar] [CrossRef] [Green Version]
- Kantor, R.S.; Wrighton, K.C.; Handley, K.M.; Sharon, I.; Hug, L.A.; Castelle, C.J.; Thomas, B.C.; Banfield, J.F. Small genomes and sparse metabolisms of sediment-associated bacteria from four candidate phyla. MBio 2013, 4, e00708–e00713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyoshi, T.; Iwatsuki, T.; Naganuma, T. Phylogenetic characterization of 16 S rRNA gene clones from deep groundwater microorganisms that pass through 0.2-micrometer-pore-size filters. Appl. Environ. Microbiol. 2005, 71, 1084–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, J.K.; Kelley, S.T.; Pace, N.R. New perspective on uncultured bacterial phylogenetic division OP11. Appl. Environ. Microbiol. 2004, 70, 845–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinke, C.; Schwientek, P.; Sczyrba, A.; Ivanova, N.N.; Anderson, I.J.; Cheng, J.F.; Darling, A.; Malfatti, S.; Swan, B.K.; Gies, E.A.; et al. Insights into the phylogeny and coding potential of microbial dark matter. Nature 2013, 499, 431–437. [Google Scholar] [CrossRef] [Green Version]
- Zachara, J.M.; Long, P.E.; Bargar, J.; Davis, J.A.; Fox, P.; Fredrickson, J.K.; Freshley, M.D.; Konopka, A.E.; Liu, C.; McKinley, J.P.; et al. Persistence of uranium groundwater plumes: Contrasting mechanisms at two DOE sites in the groundwater–river interaction zone. J. Contam. Hydrol. 2013, 147, 45–72. [Google Scholar] [CrossRef]
- Nelson, W.C.; Stegen, J.C. The reduced genomes of Parcubacteria (OD1) contain signatures of a symbiotic lifestyle. Front. Microbiol. 2015, 6, 713. [Google Scholar] [CrossRef] [Green Version]
- May, H.D.; Sowers, K.R. “Dehalobium chlorocoercia” DF-1—from Discovery to Application. In Organohalide-Respiring Bacteria; Adrian, L., Löffler, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar] [CrossRef]
- Castelle, C.J.; Wrighton, K.C.; Thomas, B.C.; Hug, L.A.; Brown, C.T.; Wilkins, M.J.; Frischkorn, K.R.; Tringe, S.G.; Singh, A.; Markillie, L.M.; et al. Genomic expansion of domain archaea highlights roles for organisms from new phyla in anaerobic carbon cycling. Curr. Biol. 2015, 25, 690–701. [Google Scholar] [CrossRef] [Green Version]
- Nazina, T.N.; Luk’yanova, E.A.; Zakharova, E.V.; Konstantinova, L.I.; Kalmykov, S.N.; Poltaraus, A.B.; Zubkov, A.A. Microorganisms in a disposal site for liquid radioactive wastes and their influence on radionuclides. Geomicrobiol. J. 2010, 27, 473–486. [Google Scholar] [CrossRef]
- Rybal’chenko, A.I.; Pimenov, M.K.; Kostin, P.P.; Balukova, V.D.; Nosukhin, A.V.; Mikerin, E.I.; Egorov, N.N.; Kaimin, E.P.; Kosareva, I.M.; Kurochkin, V.M. Deep Injection Disposal of Liquid Radioactive Waste in Russia; Battelle Press: Columbus, OH, USA, 1998; p. 206. [Google Scholar]
- Novikov, A.P.; Pavlotskaya, F.I.; Goryachenkova, T.A.; Posokhov, A.K.; Kazinskaya, I.E.; Emel’yanov, V.V. Radionuclide content in undeground waters from observation wells around Karachai Lake. Radiokhimiya 1998, 40, 484–490. [Google Scholar]
- Samsonova, L.M.; Drozhko, E.G. Migration of high-density industrial waste solutions through fresh groundwaters. In Deep Injection Disposal of Hazardous and Industrial Waste: Scientific and Engineering Aspects; Apps, J.A., Tsang, C.-F., Eds.; Academic Press: Cambridge, MA, USA, 1996; Chapter 41; pp. 669–680. [Google Scholar]
- Senko, J.M.; Istok, J.D.; Suflita, J.M.; Krumholz, L.R. In-situ evidence for uranium immobilization and remobilization. Environ. Sci. Technol. 2002, 36, 1491–1496. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.-M.; Carley, J.; Green, S.J.; Luo, J.; Kelly, S.D.; Nostrand, J.; Lowe, K.; Mehlhorn, T.; Carroll, S.; Boonchayanant, B.; et al. Effects of nitrate on the stability of uranium in a bioreduced region of the subsurface. Environ. Sci. Technol. 2010, 44, 5104–5111. [Google Scholar] [CrossRef]
- Rumynin, V.G. Radioactive brine migration at the Lake Karachai site (South Urals, Russian Federation). In Subsurface Solute Transport Models and Case Histories. Theory and Applications of Transport in Porous Media; Springer: Dordrecht, The Netherlands, 2011; Volume 25. [Google Scholar] [CrossRef]
- Nazina, T.N.; Babich, T.L.; Kostryukova, N.K.; Sokolova, D.S.; Abdullin, R.R.; Tourova, T.P. Microbial diversity and possible activity in nitrate- and radionuclide-contaminated groundwater. In Behavior of Radionuclides in the Environment I. Function of Particles in Aquatic System; Kato, K., Kalmykov, S.N., Konoplev, A., Eds.; Springer-Nature: Tokyo, Japan, 2020; Volume I, Chapter 2; ISBN 978-981-15-0678-9. [Google Scholar]
- Carini, P.; Steindler, L.; Beszteri, S.; Giovannoni, S.J. Nutrient requirements for growth of the extreme oligotroph ‘Candidatus Pelagibacter ubique’ HTCC1062 on a defined medium. ISME J. 2012, 7, 592–602. [Google Scholar] [CrossRef] [Green Version]
- Smibert, R.M.; Krieg, N.R. Phenotypic characterization. In Methods for General and Molecular Bacteriology; Gerhardt, P., Murray, R.G.E., Wood, W.A., Krieg, N.R., Eds.; ASM Press: Washington, DC, USA, 1994; pp. 607–654. [Google Scholar]
- Reynolds, E.S. The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J. Cell. Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef] [Green Version]
- Nazina, T.N.; Shestakova, N.M.; Grigor’yan, A.A.; Mikhailova, E.M.; Tourova, T.P.; Poltaraus, A.B.; Feng, C.; Ni, F.; Belyaev, S.S. Phylogenetic diversity and activity of anaerobic microorganisms of high-temperature horizons of the Dagang oil field P R China. Microbiology 2006, 75, 55–65. [Google Scholar] [CrossRef]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Trüper, H.G.; Schlegel, H.G. Sulfur metabolism in Thiorhodaceae. I. Quantitative measurements on growing cells of Chromatium okenii. J. Microbiol. Serol. 1964, 30, 321–323. [Google Scholar]
- Garrett, R.H.; Nason, A. Further purification and properties of Neurospora nitrate reductase. J. Biol. Chem. 1969, 244, 2870–2882. [Google Scholar]
- Safonov, A.V.; Babich, T.L.; Sokolova, D.S.; Grouzdev, D.S.; Tourova, T.P.; Poltaraus, A.B.; Zakharova, E.V.; Merkel, A.Y.; Novikov, A.P.; Nazina, T.N. Microbial community and in situ bioremediation of groundwater by nitrate removal in the zone of a radioactive waste surface repository. Front. Microbiol. 2018, 9, 1985. [Google Scholar] [CrossRef] [PubMed]
- Myasoedov, B.F.; Novikov, A.P. Radiochemical procedures for speciation of actinides in the environment. Methodology and data obtained in contaminated by radionuclides regions of Russia. In Proceedings of the Speciation Work Shop, Tokai-Mura, Japan, 25–28 October 1999; pp. 3–21. [Google Scholar]
- Nazina, T.N.; Kosareva, I.M.; Petrunyaka, V.V.; Savushkina, M.K.; Kudriavtsev, E.G.; Lebedev, V.A.; Ahunov, V.D.; Revenko Yu, A.; Khafizov, R.R.; Osipov, G.A.; et al. Microbiology of groundwaters from the deep repository of liquid radioactive wastes Severnyi. FEMS Microbiol. Ecol. 2004, 49, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cheng, H.; Wang, F.; Wei, D.; Wang, X. An improved 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) reduction assay for evaluating the viability of Escherichia coli cells. J. Microbiol. Meth. 2010, 82, 330–333. [Google Scholar] [CrossRef] [PubMed]
- Plakunov, V.K.; Mart’yanov, S.V.; Teteneva, N.A.; Zhurina, M.V. A universal method for quantitative characterization of growth and metabolic activity of microbial biofilms in static models. Microbiology 2016, 85, 509–513. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley & Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Edwards, U.; Rogall, T.; Bloeker, H.; Ende, M.D.; Boeettge, E.C. Isolation and direct complete nucleotide determination of entire genes, characterization of gene coding for 16S ribosomal RNA. Nucleic Acids Res. 1989, 17, 7843–7853. [Google Scholar] [CrossRef] [Green Version]
- Groβkopf, R.; Janssen, P.H.; Liesack, W. Diversity and structure of the methanogenic community in anoxic rice paddy soil microcosms as examined by cultivation and direct 16S rRNA gene sequence retrieval. Appl. Environ. Microbiol. 1998, 64, 960–969. [Google Scholar] [CrossRef] [Green Version]
- Kolganova, T.V.; Kuznetsov, B.B.; Tourova, T.P. Selection and testing of oligonucleotide primers for amplification sequencing of archaeal 16S rRNA genes. Microbiology 2002, 71, 243–246. [Google Scholar] [CrossRef]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulaam-Syed-Mohideen, A.S.; McGarrell, D.M.; Marsh, T.; Garrity, G.M.; et al. The Ribosomal Database Project: Improved alignments and new tools for rRNA analysis. Nucleic Acids Res. 2009, 37, D141–D145. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods. 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]
- Grouzdev, D.S.; Tourova, T.P.; Babich, T.L.; Shevchenko, M.A.; Sokolova, D.S.; Abdullin, R.R.; Poltaraus, A.B.; Toshchakov, S.V.; Nazina, T.N. Whole-genome sequence data and analysis of type strains ‘Pusillimonas nitritireducens’ and ‘Pusillimonas subterraneus’ isolated from nitrate- and radionuclide-contaminated groundwater in Russia. Data Brief 2018, 21, 882–887. [Google Scholar] [CrossRef] [PubMed]
- Grouzdev, D.S.; Babich, T.L.; Tourova, T.P.; Sokolova, D.S.; Abdullin, R.R.; Poltaraus, A.B.; Schevchenko, M.A.; Toshchakov, S.V.; Nazina, T.N. Draft genome sequence of Roseomonas aestuarii strain JR1/69-1-13 isolated from nitrate- and radionuclide-contaminated groundwater in Russia. Genome Announc. 2018, 6, e00583-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Ciufo, S.; Li, W. Prokaryotic genome annotation pipeline. In The NCBI Handbook [Internet], 2nd ed.; NCBI: Bethesda, MD, USA, 2013. Available online: http://www.ncbi.nlm.nih.gov/books/NBK174280 (accessed on 10 December 2013).
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid annotations using subsystems technology. BMC Genomics. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Markowitz, V.M.; Mavromatis, K.; Ivanova, N.N.; Chen, I.M.A.; Chu, K.; Kyrpides, N.C. IMG ER: A system for microbial genome annotation expert review and curation. Bioinformatics 2009, 25, 2271–2278. [Google Scholar] [CrossRef] [Green Version]
- Pal, C.; Bengtsson-Palme, J.; Rensing, C.; Kristiansson, E.; Larsson, D.G.J. BacMet: Antibacterial biocide and metal resistance genes database. Nucleic Acids Res. 2014, 42, D737–D743. [Google Scholar] [CrossRef] [Green Version]
- Solodov, I.N.; Zotov, A.V.; Khoteev, A.D. Geochemistry of natural and contaminated underground waters in fissured bed bedbedrocks of the Lake Karachai area. Appl. Geochem. 1998, 13, 921–939. [Google Scholar] [CrossRef]
- Kim, M.; Oh, H.S.; Park, S.C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef]
- Glöckner, F.O.; Zaichikov, E.; Belkova, N.; Denissova, L.; Pernthaler, J.; Pernthaler, A.; Amann, R. Comparative 16S rRNA analysis of lake bacterioplankton reveals globally distributed phylogenetic clusters including an abundant group of Actinobacteria. Appl. Environ. Microbiol. 2000, 66, 5053–5065. [Google Scholar] [CrossRef] [Green Version]
- Konstantinidis, K.T.; Ramette, A.; Tiedje, J.M. The bacterial species definition in the genomic era. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 1929–1940. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [Green Version]
- Wall, J.D.; Krumholz, L.R. Uranium reduction. Annu. Rev. Microbiol. 2006, 60, 149–166. [Google Scholar] [CrossRef] [Green Version]
- North, N.; Dollhopf, S.L.; Petrie, L.; Istok, J.D.; Balkwill, D.L.; Kostka, J.E. Change in bacterial community structure during in situ biostimulation of subsurface sediment cocontaminated with uranium and nitrate. Appl. Environ. Microbiol. 2004, 70, 4911–4920. [Google Scholar] [CrossRef] [Green Version]
- Green, S.J.; Prakash, O.; Jasrotia, P.; Overholt, W.A.; Cardenas, E.; Hubbard, D.; Tiedje, J.M.; Watson, D.B.; Schadt, C.W.; Brooks, S.C.; et al. Denitrifying bacteria from the genus Rhodanobacter dominate bacterial communities in the highly contaminated subsurface of a Nuclear Legacy Waste Site. Appl. Environ. Microbiol. 2012, 78, 1039–1047. [Google Scholar] [CrossRef] [Green Version]
- Newsome, L.; Morris, K.; Lloyd, J.R. The biogeochemistry and bioremediation of uranium and other priority radionuclides. Chem. Geol. 2014, 363, 164–184. [Google Scholar] [CrossRef]
- Alexakhin, A.I.; Glagolev, A.V.; Drozhko, E.G.; Zinin, A.I.; Zinina, G.A.; Ivanov, I.A.; Mokrov, Y.G.; Orlova, E.I.; Samsonov, B.G.; Samsonova, L.M.; et al. Reservoir-9—Storage of Liquid Radioactive Waste and Its Impact on the Geological Environment; Drozhko, E.G., Samsonov, B.G., Eds.; Atomic Energy Agency: Moscow, Russia, 2007; p. 250. ISBN 978-5-9901363-1-1. [Google Scholar]
- Weidler, G.W.; Gerbl, F.W.; Stan-Lotter, H. Crenarchaeota and their role in the nitrogen cycle in a subsurface radioactive thermal spring in the Austrian Central Alps. Appl. Environ. Microbiol. 2008, 74, 5934–5942. [Google Scholar] [CrossRef] [Green Version]
- Hemme, C.L.; Tu, Q.; Shi, Z.; Qin, Y.; Gao, W.; Deng, Y.; Nostrand, J.D.; Wu, L.; He, Z.; Chain, P.S.; et al. Comparative metagenomics reveals impact of contaminants on groundwater microbiomes. Front. Microbiol. 2015, 6, 1205. [Google Scholar] [CrossRef]
- Spain, A.M.; Peacock, A.D.; Istok, J.D.; Elshahed, M.S.; Najar, F.Z.; Roe, B.A.; White, D.C.; Krumholz, L.R. Identification and isolation of a Castellaniella species important during biostimulation of an acidic nitrate- and uranium-contaminated aquifer. Appl. Environ. Microbiol. 2007, 73, 4892–4904. [Google Scholar] [CrossRef] [Green Version]
- Hahn, M.W. Isolation of strains belonging to the cosmopolitan Polynucleobacter necessarius cluster from freshwater habitats located in three climatic zones. Appl. Environ. Microbiol. 2003, 69, 5248–5254. [Google Scholar] [CrossRef] [Green Version]
- Wrighton, K.C.; Castelle, C.J.; Wilkins, M.J.; Hug, L.A.; Sharon, I.; Thomas, B.C.; Handley, K.M.; Mullin, S.W.; Nicora, C.D.; Singh, A.; et al. Metabolic interdependencies between phylogenetically novel fermenters and respiratory organisms in an unconfined aquifer. ISME J. 2014, 8, 1452–1463. [Google Scholar] [CrossRef] [Green Version]
- Lever, M.A.; Rogers, K.L.; Lloyd, K.G.; Overmann, J.; Schink, B.; Thauer, R.K.; Tori, M.; Hoehler, T.M.; Jørgensen, B.B. Life under extreme energy limitation: A synthesis of laboratory- and field-based investigations. FEMS Microbiol. Rev. 2015, 39, 688–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nies, D.H. Microbial heavy-metal resistance. Appl. Microbiol. Biotechnol. 1999, 51, 730–750. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Pan, C.; Xiao, A.; Yang, X.; Zhang, G. Isolation, identification, and environmental adaptability of heavy-metal-resistant bacteria from ramie rhizosphere soil around mine refinery. 3 Biotech. 2017, 7, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kothari, A.; Wu, Y.-W.; Chandonia, J.-M.; Charrier, M.; Rajeev, L.; Rocha, A.M.; Joyner, D.C.; Hazen, T.C.; Singer, S.W.; Mukhopadhyay, A. Large circular plasmids from groundwater plasmidomes span multiple incompatibility groups and are enriched in multimetal resistance genes. MBio 2019, 10, e02899-18. [Google Scholar] [CrossRef] [Green Version]


| Parameters | Standard Error | Observation Wells | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 2/68 * | 14/68 | 1/69 * | 1/69 | 176/94 | |||||
| Distance from the Karachai, m | 2100 | 2000 | 3200 | 2500 | |||||
| Year of Analyses | 2010 | 2010 | 2013 | 2011 | 2013 | 2012 | |||
| Depth of Sampling, m | 80 | 80 | 20–30 | 60 | 44 | 20–30 | 60 | 60 | |
| pH | ±0.1 | 8.0 | 7.6 | 7.7 | 7.2 | 7.9 | 7.3 | 6.9 | 7.8 |
| Eh, mV | ±1.0 | 310 | 270 | 110 | 120 | 200 | 110 | 420 | 120 |
| Na+ + K+, mg L−1: | ±0.02 | 9071 | 1099 | 340 | 3230 | 145 | 370 | 2610 | 356 |
| Ca2+ + Mg2+, mg L−1: | ±0.09 | 6712 | 3102 | - | - | 302 | - | - | 2460 |
| Fetotal, mg L−1: | ±0.004 | 12.1 | 0.86 | - | - | 3.3 | - | - | 0.75 |
| Cl−, mg L−1: | ±0.004 | - | - | - | - | 140 | - | - | 215 |
| SO42−, mg L−1: | ±0.02 | - | - | - | - | 110 | - | - | 80 |
| NO3−, mg L−1: | ±0.09 | 42,700 | 13,600 | 120 | 2900 | 120 | 200 | 2300 | 2090 |
| CO32−/HCO3−, mg L−1: | ±0.23 | 1310 | 267 | 80 | 120 | 360 | 59 | 200 | 1820 |
| Acetate, mg L−1: | ±0.5 | 921 | 795 | - | - | 26 | - | - | - |
| Radioactivity, Bq L−1 | - | - | - | - | - | - | - | - | |
| 239,240Pu | 0.01 | 5.78 | - | 0.03 ± 0.01 | 0.18 ± 0.04 | 0.14 | <0.02 | 0.12 ± 0.03 | 0.8 |
| 90Sr | ±0.001 | - | - | 220 ± 30 | 3300 ± 300 | 12.4 | 155 ± 20 | 2200 ± 250 | 300 |
| 137Cs | ±0.001 | - | - | <0.2 | 2.9 ± 0.4 | <0.2 | <0.2 | - | |
| 241Am | ±0.001 | 3.01 | - | <0.02 | 0.06 ± 0.01 | 0.8 | <0.02 | 0.20 ± 0.04 | 0.7 |
| 237Np | ±0.001 | 7.2 | - | <0.02 | 1.0 ± 0.3 | 0 | <0.02 | 3.7 ± 1.2 | <0.5 |
| 238U, Bq·L−1 or mg L−1 ** | 39 | - | 3.1 ± 0.7 ** | 12.1 ± 2.0 ** | 2.6 | 0.3 ± 0.1 ** | 1.1 ± 0.2 ** | 45 | |
| SR rate, µg S2−/(L day) | ±0.001 | - | - | - | - | 0.01 | - | - | 0 |
| MG rate, µg CH4/(L day) | ±0.001 | - | - | - | - | 0 | - | - | 0 |
| Strain | Closest Cultivated Microorganism, Accession No. of 16S rRNA Gene | Accession No.* of 16S rRNA Gene | 16S rRNA Similarity, % | Genome Size, Mb |
|---|---|---|---|---|
| JR1/69-1-13 | Roseomonas aestuarii JC17T, NR_116913 | MG205609 | 99.6 | 5.2 |
| JR1/69-2-13 | Pusillimonas harenae B201, GQ232740 | MG205613 | 98.6 | 4.3 |
| JR1/69-3-13 | Pusillimonas harenae B201, GQ232740 | MG205614 | 98.5 | 4.5 |
| J1-13Tb | Chryseobacterium haifense H38T, NR_044167 | MG205639 | 98.2 | 2.8 ** |
| JR3/2-4 | Rhodococcus qingshengii JCM 15477T, DQ090961 | MG205632 | 99.7 | 7.2 ** |
| J1-4 | Rhodococcus fascians ATCC 12974T, NR_119126 | MG205633 | 100 | 5.8 ** |
| JR2/2-1 | Mycolicibacterium iranicum M05T, NR_117909 | MG205634 | 100 | 6.3 ** |
| JR43/78-4-13 | Janibacter limosus DSM 11140T, NR_026362 | MG205635 | 99.7 | 3.4 ** |
| JR2/1-6 | Salinibacterium amurskyense KMM 3673T, AF539697 | MG205636 | 98.5 | 2.8 ** |
| JR3/2-13 | Microbacterium oxydans DSM 20578T, NR_044931 | MG205637 | 99.6 | 3.9 ** |
| JR1/1-8b | Paeniglutamicibacter sulfureus DSM 20167T, NR_026237 | MG205638 | 99.3 | ND *** |
| Characteristic | Strain | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| JR3/2-13 | J1-13Tb | JR43/78-4-13 | JR2/1-6 | JR2/2-1 | JR3/2-4 | J1-4 | JR1/1-8b | JR1/69-2-13 | JR1/69-3-13 | JR1/69-1-13 | |
| Cells morphology | Rods | Rods, cocci | Cocci, rods | Rods | Rods | Pleomorphic | Pleomorphic | Cocci, rods | Rods | Rods | Coccobacilli |
| Cell size, µm | 0.1–0.2 × 0.4–0.9 | 0.4–0.9 × 1.3–2.5; 0.3 | Cocci 0.3–1.2; rods 0.9 × 1.3 | 0.3 × 1.3–2.0 | 0.8–1.5 × 1.6–2.4 | 0.9–1.2 × 1.2–4.7; 0.9–2.5 | 1.1–1.2 × 1.5–3.1; 0.9–1.5 | Cocci 0.7–1.6 | 0.8–1.2 × 1.5–2.1 | 0.7–1.3 × 0.9–2.1 | 1.2–1.6 × 1.5–2.4 |
| Motility | + | − | − | − | − | − | − | − | + | + | + |
| Substrates for aerobic growth | |||||||||||
| Acetate | + | W | + | + | + | + | + | + | + | − | |
| Oxalate | − | + | − | − | − | − | − | − | − | ||
| Glucose | + | + | − | + | + | + | + | + | + | + | + |
| Sucrose | + | + | − | + | + | + | + | + | W | W | + |
| Ethanol | W | − | + | + | + | W | − | − | − | ||
| Alanine | W | + | − | − | − | + | + | − | − | ||
| Lactate | + | + | + | − | + | + | W | + | W | + | |
| Acetate + Fe3+→Fe2+ | − | − | − | − | − | − | − | − | + | ||
| Acetate + NO3−→NO2− | − | − | + | − | − | − | W | + | + | + | + |
| Acetate + NO3−→N2 | − | − | − | − | − | − | − | + | + | − | − |
| Temperature, range/optimum, °C | 5–42/30 | 5–40/30–32 | 4–50/28 | 5–37/25–28 | 13–42/35 | 21–42/33 | 13–35/28 | 28–38/28 | 5–35/30 | 5–35/30 | 13–42/30 |
| NaCl, range/optimum (%, w/v) | 0–7/0.5–3 | 0–2/0–1.5 | 0–4 | 0–10/1–2 | 0–5(7)/0–2 | 0–5(7)/0–2 | 0–7(9)/ 0–3 | 0–8/0.5–2 | 0–3/0.5–1 | 0–5 0.5–3 | 0–3/0.5–1 |
| Strain | LD50, LD100 | Radiation Dose (kGy) | Maximum Concentration (mg L−1) | ||||
|---|---|---|---|---|---|---|---|
| Zn2+ | Cr6+ | Cu2+ | Cd2+ | U6+ | |||
| R. aestuarii JR1/69-1-13 | LD50 | 5.0 | 10 | 25 | 50 | 50 | 10 |
| LD100 | 15.0 | 100 | 100 | >200 | 200 | 100 | |
| Pusillimonas sp. JR1/69-2-13 | LD50 | 1.0 | 50 | 25 | 100 | 100 | 250 |
| LD100 | 10.0 | 200 | 200 | >200 | 300 | >400 | |
| Pusillimonas sp. JR1/69-3-13 | LD50 | 1.0 | 50 | 100 | 100 | 100 | 250 |
| LD100 | 10.0 | 200 | 400 | >200 | 300 | >400 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nazina, T.; Babich, T.; Kostryukova, N.; Sokolova, D.; Abdullin, R.; Tourova, T.; Kadnikov, V.; Mardanov, A.; Ravin, N.; Grouzdev, D.; et al. Ultramicrobacteria from Nitrate- and Radionuclide-Contaminated Groundwater. Sustainability 2020, 12, 1239. https://doi.org/10.3390/su12031239
Nazina T, Babich T, Kostryukova N, Sokolova D, Abdullin R, Tourova T, Kadnikov V, Mardanov A, Ravin N, Grouzdev D, et al. Ultramicrobacteria from Nitrate- and Radionuclide-Contaminated Groundwater. Sustainability. 2020; 12(3):1239. https://doi.org/10.3390/su12031239
Chicago/Turabian StyleNazina, Tamara, Tamara Babich, Nadezhda Kostryukova, Diyana Sokolova, Ruslan Abdullin, Tatyana Tourova, Vitaly Kadnikov, Andrey Mardanov, Nikolai Ravin, Denis Grouzdev, and et al. 2020. "Ultramicrobacteria from Nitrate- and Radionuclide-Contaminated Groundwater" Sustainability 12, no. 3: 1239. https://doi.org/10.3390/su12031239
APA StyleNazina, T., Babich, T., Kostryukova, N., Sokolova, D., Abdullin, R., Tourova, T., Kadnikov, V., Mardanov, A., Ravin, N., Grouzdev, D., Poltaraus, A., Kalmykov, S., Safonov, A., Zakharova, E., Novikov, A., & Kato, K. (2020). Ultramicrobacteria from Nitrate- and Radionuclide-Contaminated Groundwater. Sustainability, 12(3), 1239. https://doi.org/10.3390/su12031239