1. Introduction
Microalgae are unicellular photosynthetic organisms that use light and carbon dioxide, with higher photosynthetic efficiency than plants, for the production of biomass. Some microalgae species can also grow and multiply heterotrophically in the absence of light if an organic carbon source becomes available [
1]. Τhe main advantage of heterotrophic growth is higher biomass growth rates and biomass production as, unlike autotrophic growth, heterotrophic growth is not limited by light transmission through the growth medium. Another advantage of heterotrophic growth is the potential of achieving higher lipid content and, as a result, higher lipid productivities. This is needed if microalgal cultivation is to be useful for biodiesel production. Disadvantages of heterotrophic growth are the susceptibility to contamination, which requires that all parts of the bioreactors, as well as all growth media, must be carefully sterilized in order to minimize the risk of contamination, and the cost of organic carbon which must be provided to the growth medium.
The microalgae growth rate, as well as the protein and lipid content, are influenced by many parameters, such as the cultivated species, the temperature, the concentration of carbon, the ratio of carbon to nitrogen, the medium pH, the concentration of potassium, phosphorus and micronutrients, dissolved oxygen availability, and the mixing rate [
2]. For biodiesel production, except for a high lipid productivity a favorable fatty acid profile of the lipids is needed as well. Currently, biodiesel is mainly produced from used oils and various seed oils such as sunflower and cotton seed oils. Vegetable oils contain fatty acid mostly from C14:0 to about C22:0. Used oils contain a small amount of animal fats which have a lower fatty acid chain length.
Depending on the cultivated species and the cultivation mode and parameters, the microalgal biomass can be directed towards specific uses, as various components such as proteins, lipids, antioxidants, and other compounds can be maximized [
3]. Therefore, the microalgal biomass composition can be altered by altering the growth conditions and furthermore microalgal cultivation can be carried out on an industrial scale using basic biochemical engineering principles. In addition to the production of biodiesel, the biomass can be used in a variety of applications. The biomass of microalgae is rich in antioxidants and some organic substances of pharmaceutical value, so some applications are in the cosmetic and pharmaceutical industry [
4]. The high protein and lipid content can be of value in aquaculture for the replacement of fishmeal in fish feeds and in the poultry industry [
5].
In heterotrophic cultivation mode, microalgae use organic carbon and oxygen under dark conditions for growth and biomass production [
6]. A number of review papers focus on the heterotrophic growth of several microalgal species, and the trend is that heterotrophic growth enhances both the biomass and lipid productivity [
7,
8]. Different sources of carbon can be used for microalgae growth, such as glucose, sucrose, fructose, mannose, glycerol, lactose, or galactose [
9]. Additionally, various industrial effluents and by-products, such as anaerobic digestate, can be used as a carbon and nitrogen source. Studies on the heterotrophic cultivation of different microalgal species using glucose as carbon source have been published [
10,
11], while work on the cultivation of microalgae using glycerol is limited. It should be mentioned that studies on cultivation using industrial effluents and by-products, such as anaerobic digestate are lacking.
In the heterotrophic cultivation of
Chlorella sorokiniana, the growth rate was higher using glucose and sodium acetate as the carbon source compared to fructose [
12]. In
Chlorella vulgaris, optimal cell growth and lipid productivity were attained using glucose at 1% (
w/
v). Growth of
C. vulgaris on glycerol had similar dose effects as those from glucose [
13]. Ogbonna et al. [
14] report that the heterotrophic cultivation of
Euglena gracilis was enhanced when cultivated with glucose in comparison with cultivation with ethanol. In
Chlorella protothecoides, corn powder hydrolysate was used instead of glucose as an organic carbon source in heterotrophic culture medium in fermenters, in order to increase the biomass and reduce the cost of cultivation [
15]. O’Grady and Morgan [
16] studied the heterotrophic growth of
Chlorella protothecoides with different carbon sources, specifically glycerol, glucose, and a glucose/glycerol mixture. They found that the specific growth rate and lipid yields when using glycerol and a glucose/glycerol mixture were higher in comparison when using only glucose. Similarly, Kong et al. [
17] found that the growth rate of
C. vulgaris, as well as the biomass production of the species, was enhanced when cultivated with a mixture of glycerol and glucose.
Anaerobic digestate (AD) produced by biogas plants is mainly used for field fertilization in various crops such as corn. When there are no adequate fields available, or the land is not sufficiently flat, nitrification of underground and surface water and high soil salinity may result as excess anaerobic digestate is applied. Therefore, developing new applications for the efficient use of anaerobic digestate is needed in order to utilize the macro- and micronutrient content of the AD in the framework of circular economy. A good solution is the use of anaerobic digestate for microalgae cultivation, as it is a good source of nitrogen, phosphorus, and potassium, as well as a source of over 30 different micronutrients. Potassium and phosphorus availability are projected to decrease in the future.
The microalgae species Chlorella sorokiniana was cultivated with glycerol as the main source of carbon and 0%, 15%, 30%, and 50% AD added to the growth media. In all four cultivation treatments using glycerol and AD, both initial carbon and nitrogen concentrations, as well as pH and temperature, were held constant in order to determine the effect of AD on the growth rate, the lipid and protein content of the biomass, and the FA distribution. The four FAME properties, namely SN, IV, CN, and HHV, were calculated from the FA distribution and empirical equations. Nitrogen was in the form of ammonium as this is the form found in AD. Potassium and phosphorus were held constant as well. In addition, Chlorella sorokiniana was cultivated without glycerol, (AD/water (v/v): 75:25, 50:50, and 25:75) in order to determine the rate of carbon uptake of the partly digested organic material present in the AD.
Most studies have been carried out with piggery and municipal wastewater, with or without glycerol, with autotrophic or mixotrophic cultivations, and using other than Chlorella sorokiniana species. There are no studies which examine the effect of anaerobic digestate on the fatty acid distribution of the lipids obtained from a heterotrophic cultivation, with or without glycerol. The purpose of the present study was to investigate the effect of different percentages of anaerobic digestate enriched with crude glycerol in the growth medium of Chlorella sorokiniana under heterotrophic growth conditions on the fatty acid distribution of the lipids obtained and the basic biodiesel properties.
2. Materials and Methods
2.1. Bioreactors
The cultivation of C. sorokiniana was carried out in glass cylindrical bioreactors of 42 L capacity each that were filled to 80% of their volume. Air was continuously provided to each bioreactor though a perforated network of piping placed at the bottom of the bioreactor tank. The temperature of the cultures was kept at 30 ± 1 °C with the use of temperature thermostats (Aquael 250 W heaters, Suwalki, Poland) and the pH was held constant at 7 ± 0.3. The pH was adjusted manually as needed two or three times daily with the use of HCl or NaOH solutions. The bioreactors, the glass tubing, and the culture medium were sterilized before use.
2.2. Inoculum Preparation
The microalgae species
C. sorokiniana (SAG strain 211-31) was obtained from Culture Collection of Algae from the University of Göttingen in Germany (EPSAG). The cultivation medium used for the inoculum growth was the Basal Medium (=ES “Erddekokt + Salze”) and each liter of it contained: 0.2 g KNO
3/L, 0.02 g K
2HPO
4/L, 0.02 g MgSO
4·7H
2O/L, 30 mL of soil extract/L, and 5 mL/L of a solution containing the following micronutrients: (1 mg ZnSO
4·7H
2O, 2 mg MnSO
4·4H
2O, 10 mg H
3BO
3, 1 mg Co(NO
3)
2.6H
2O, 1 mg MoO
4·2H
2O, 0.005 mg CuSO
4·5H
2O, 700 mg FeSO
4·7H
2O, and 800 mg EDTA)/L [
18].
The culture medium was inoculated with a standard quantity (250 mL of C. sorokiniana inoculum) which was prepared as follows: 1 L flasks, containing the necessary culture medium, were inoculated with C. sorokiniana culture directly obtained from EPSAG and cultivated in a sterile environment until they reached an optical density of about 0.5. The cultivation of the inoculum was always performed under the same conditions, namely at a temperature of 25 °C, under natural illumination, and by using an orbital shaker at 60 rpm in order to prevent sticking of algae to the surfaces of the flask.
2.3. Measurements
In all experiments, the organic carbon concentration was measured daily so that its uptake by the microalgae could be monitored. Initial nitrogen concentration (Co), in the form of ammonium, was also held constant. The organic carbon reduction kinetics (as rate of Carbon uptake) was determined from the slope of the experimental data of C(t) vs. t, by plotting the C(t) vs. t data and obtaining the average slope ΔC(t)/Δt.
The saponification number (
SN) and the iodine value (
IV) were calculated theoretically from the FA distribution using the equations suggested by Kalayasiri et al. [
19] and by Azam et al. [
20]. Similarly, the cetane number (
CN) was evaluated from the theoretical equation suggested by Krisnangkura [
21] and the higher heating value (
HHV) from the equation suggested by Demirbas [
22].
where %
Wi is the % weight of each FA,
Ndb is the number of double bonds, and
MWi is the molecular weight of the respective FAME.
2.4. Method of Analyses
The composition of the samples was determined according to AOAC [
23] methods.
Total nitrogen content in samples was measured with liquid digestion. Digestion is accomplished by boiling of the samples in concentrated sulphuric acid with catalyst potassium sulphate, copper sulphate, and selenium. After cooling, the digestion product was distilled with NaOH in a solution 2% H
3BO
3 in presence of indicator methyl red and was titrated with 0.1 N HCl [
24]. Total protein content of plant tissues was calculated from the total nitrogen using a conversion factor of 6.25 [
25].
For the determination of organic carbon, the method of Walkley–Black was used. The samples were first centrifuged and then filtered. According to this method, organic carbon was oxidized by a mixture of K2Cr2O7 and H2SO4 in a ratio of 1:2. The remaining K2Cr2O7 was titrated with 0.5 N FeSO4 7H2O.
The total lipid content was determined with extraction using co-solvents of n-hexane/isopropanol in the microalgal biomass in accordance with the method of Bian et al. [
26]. The solvent ratio of n-hexane to isopropanol was 3/2 (
v/
v) and the ratio of co-solvents to microalgal dry biomass was 10/1 (
v/
w). Extraction was carried out in horizontal-circular movement “tehtnica Zelezniki EV-402” machine. Stirring speed was 300 rpm, and the extraction time was 48 h. The total lipid extract was determined gravimetrically after filtration and evaporation of the solvents.
The bio-oil was subjected to transesterification and the fatty acid distribution was determined by gas chromatography. Specifically, the bio-oil was filtered and 100 mg of the bio-oil was reacted with a mixture of heptane and KOH/MeOH (2 N). The anhydrous MeOH was filtered. Following the separation of the two phases, 1 μL of the FAME (Fatty Acid Methyl Ester) phase was injected into an Agilent Gas Chromatographer Model 6890 N. Analysis of the FAME distribution was performed according to the EN 14103 method.
2.5. Materials
Crude glycerol was used as the carbon source. It was obtained from a local biodiesel manufacturing plant. Its methanol was removed, and its composition was 86% glycerin, 0.5% methanol, 7.5% water, 3.5% MONG (Non-Glycerin Organic Matter), and 2.5% ash. Anaerobic digestate was provided by a local biogas producing plant. It was first filtered through three successive sieves and then centrifuged at 4000 rpm for 10 min. It was then sterilized by boiling for about 10 min. Following its cooling, the macronutrient content of the AD was determined. Nitrogen in the AD is present in the form of ammonium ions while carbon comes from partly digested organic material. The ammonium concentration in the biofertilizer was measured at 1129 mg/L, which corresponds to 878.1 mg/L of elemental nitrogen. Therefore, at the maximum percentage of 50% biofertilizer, the concentration of elemental nitrogen is 439 mg/L. In the first 3 bioreactors the remaining nitrogen was added as NH4Cl. Organic carbon concentration in the AD was 1.95 g/L.
2.6. Initial Parameters of the Variable AD Experiment
Table 1 summarizes the initial concentrations of carbon and nitrogen, the Co/No ratio, and the AD used for the experiments where AD varied from 0% to 50% (
v/
v) in the growth media.
2.7. Statistical Analysis
Data were analyzed using the IBM SPSS Statistics 24 statistical package [
27]. The experiment had four replications. Analysis of variance was used to assess treatment effects. Mean separation was made using Tukey’s test when significant differences (
p = 0.05) between treatments were found. All presented numeric values are means of four measurements ± standard deviation (SD).
3. Results and Discussion
3.1. Growth and Carbon Uptake Rate Using Only Anaerobic Digestate
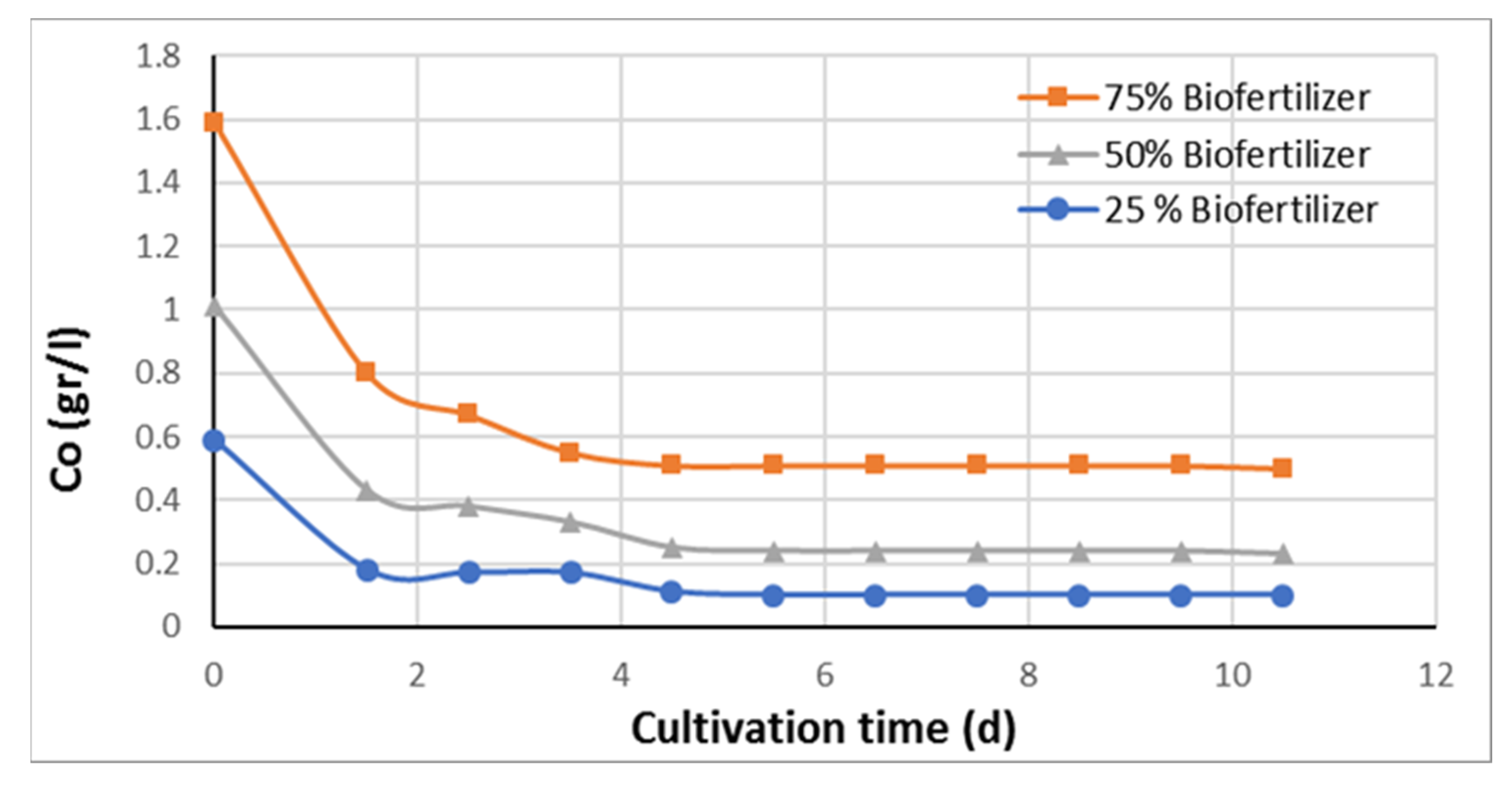
Figure 1 shows the carbon concentration as a function of cultivation time for
Chlorella sorokiniana cultivated only in diluted pure anaerobic digestate without any glycerol and other macro- or micronutrient added. The AD concentration was approximately 75%, 50%, and 25%, and water was used in order to dilute the AD. Therefore, both carbon and nitrogen are solely supplied by the AD. The only purpose of this experiment was to determine the utilization or uptake, by the microalgal species
Chlorella sorokiniana, of the organic carbon found as undigested or partly digested organic compounds in the AD. Carbon in AD is a mixture of unspecified fragments of various organic molecules present in the raw materials such as proteins, lipids, carbohydrates, hemi-cellulose, and cellulose.
The organic carbon and ammonium ion concentrations of the AD (100%) were determined and were equal to 1129 mg/L and 1.95 g/L, respectively. Therefore, the initial concentration of elemental nitrogen is equal to 878 mg/L and the initial Co/No is equal to 2.2. From the data of
Figure 1, it appears that the organic carbon in the AD is easily absorbed and utilized by
Chlorella sorokiniana, as there is a high percentage of carbon uptake in just 2 to 4 days, and 70% to almost 85% of the undigested organic carbon in the AD is utilized.
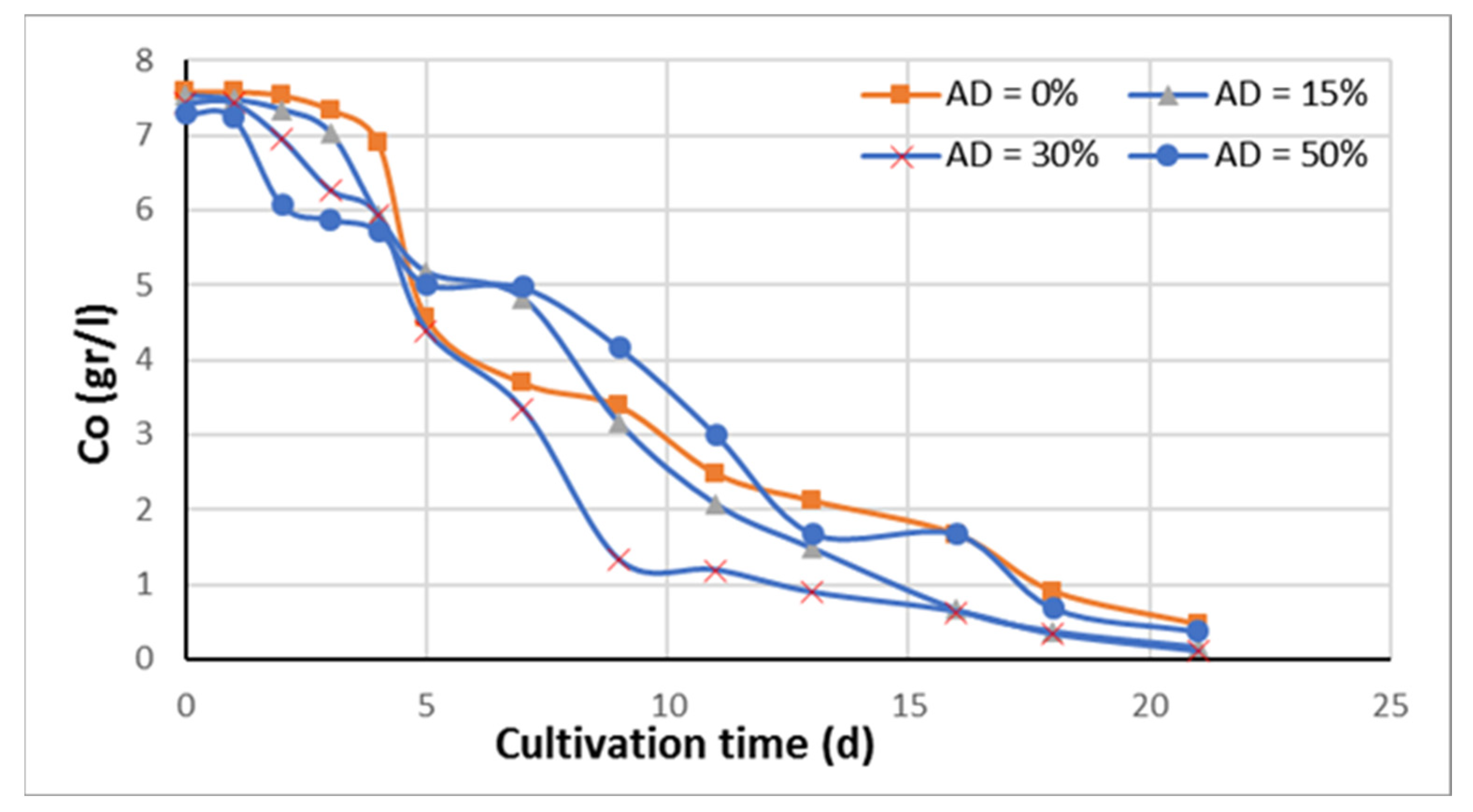
3.2. Kinetics of the Growth Rate Using Anaerobic Digestate and Glycerol
Figure 2 shows the rate of carbon decrease for four different AD concentrations in the growth medium equal to 0% (standard), 15%, 30%, and 50%, using glycerol as the main carbon source. The measured Co/No ratio was basically the same for all bioreactors and ranged from 18.2 to 19.0. Carbon was mostly from glycerol and the amount of glycerol added in each bioreactor was adjusted so as to take into account the undigested carbon found in the AD. Nitrogen in the form of ammonium came from the AD and ammonium chloride. Its initial concentration was constant in all bioreactors by adjusting the ammonium chloride added to the first three bioreactors. Its initial nitrogen concentration was 439 mg/L. Therefore, the purpose of this experiment was to (a) examine the effect of the AD on the growth rate, (b) examine the lipid and protein content of the biomass, and (c) determine the FA distribution of the bio-oil extracted from the biomass and estimate basic properties of the corresponding fatty acid methyl esters (FAME).
Carbon concentration fell to a value less than 0.5 g/L in about 18–20 days. The lag phase was from 1 to 2 days. During the intermediate growth of Chlorella sorokiniana, between about the 3rd and 12th day, the growth is fast, as is the carbon rate of decrease. This is the exponential gorwth phase which is very fast. In cultivations, there are realized four growth phases: (a) the lag phase in which the microorganisms adjust to the growth media environment, (b) the exponential growth phase, characterized by fast growth, (c) the stationary phase during which growth is slowed, and (d) the death phase occurring much later in time.
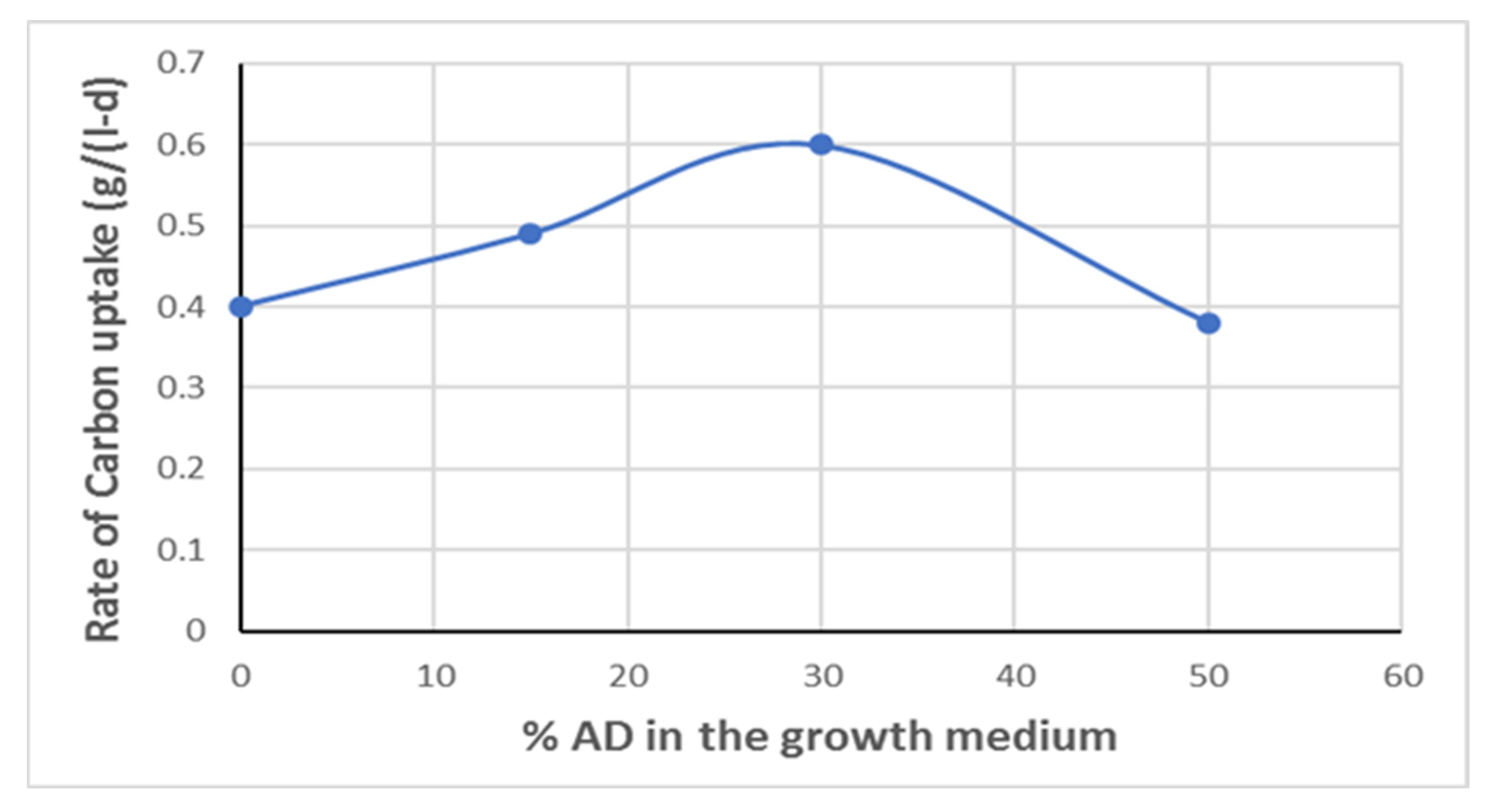
From the data of
Figure 2, the average carbon uptake rate in the exponential growth phase was calculated and shown in
Figure 3. It is seen that the carbon uptake rate goes through a maximum at 30% AD. It is higher by about 33% compared to 0% AD. Carbon concentration drops to less than 1 g/L in 13 days for the 30% AD, while the respective time for the 0% AD and 50% is about 18 days.
Anaerobic digestate is a very complex medium. It is used, as it contains all the macro- and micronutrients needed for the cultivation of microalgae. Specifically, it contains a great number of elements, basically all elements, as well as various organic compounds. To add to this complexity, other unknown compounds may be formed from the anaerobic digestion. Micronutrients present in the AD, especially cobalt, molybdenum, and iron, may be affecting carbon uptake kinetics; this is presently being investigated. Cobalt and especially molybdenum may also be affecting nitrogen uptake by the microalgal cells which utilize these micronutrients for cell growth and therefore may be directly involved in the carbon utilization process.
Several studies have been conducted investigating the use of anaerobic digestion as a nutrient source for the cultivation of a variety of microalgal sp. Chuka-ogwudea et al. [
28] reviewed research on microalgal cultivation using anaerobic digestate produced from food waste. Various food wastes and agricultural energy plants were used. Various
Chlorella sp., such as
pyrenoidosa,
vulgaris,
minutissima, and
sorokiniana, as well as various
Scenedesmus sp.,
Nannochloropsis sp., and
Tetraselmis sp. were used. Singh et al. [
29] studied the mixotrophic cultivation of
Chlorella sorokiniana using different growth media from anaerobic effluent from a poultry litter anaerobic digester. The lipid content varied between 8.2% and 12.4%, while the protein content varied between 37.9% and 39.2%. The biomass productivity was 63 mg L
−1 d
−1. Rajagopal et al. [
30] studied the autotrophic cultivation of
Chlorella vulgaris strain CPCC 90 using anaerobic digestate (AD) from chicken manure. They diluted the AD and prepared AD media containing AD from 10% up to 90% in increments of 20%. They found that, due to the high ammonia content of the AD, growth was inhibited in growth media containing AD. Sekine et al. [
31] studied the cultivation of
Chlorella sorokiniana strain NIES-2173 using undiluted anaerobic digestate and compared with the cultivation using a synthetic growth medium. The growth of
Chlorella sorokiniana strain NIES-2173 using undiluted anaerobic digestate was almost the same as that with using the synthetic growth medium. Dilution of the anaerobic digestate resulted in a decrease in the specific growth rate coefficient.
3.3. Lipid and Protein Content of the Biomass
The lipid and protein content of the biomass in the experiments where the concentration of the AD varied between 0% and 50% is shown in
Table 2. The protein content in the biomass of
Chlorella sorokiniana increased by the addition of 30% and 50% percentages of AD in the growth medium compared with the control (crude glycerol + 0% AD), while, by the addition of 15% of AD in the growth medium, the protein content in the biomass of
Chlorella sorokiniana did not show statistically significant differences compared with the control (
Table 2). In contrast, the lipid content in the biomass of
Chlorella sorokiniana decreased by the addition of 30% and 50% of AD in the growth medium compared with the control (crude glycerol + 0% AD), while the addition of 15% of AD in the growth medium did not show statistically significant differences in the lipid content in the biomass of
Chlorella sorokiniana compared with the control (
Table 2).
It is noted that up to 15% AD protein and lipid contents are basically unaltered. However, increasing the AD% to 30% and 50% decreases the lipid content by about 3% and 4%, respectively. However, smaller or equal to 15% percentages of AD can be used without a substantial reduction in lipid content of the biomass. Additionally, other modes of using the AD, such as semi-batch addition, may be used in order to improve the lipid content of the biomass, or the Co/No ratio can be increased, and this will lead to an increase in the lipid content of the biomass. The lipid content is relatively low as, although a relatively high Co was used, the high No value (439 mg/L), which was used due to the AD, favors protein synthesis instead.
3.4. Fatty Acid Distribution and Estimation of Biodiesel (FAME) Properties
Following collection of the biomass, the bio-oil was extracted. The Fatty Acid distribution is shown in
Table 3.
It is noted that, as the percentage of AD in the growth medium is increased, FA with longer chain length are produced. With respect to the chain length, it is observed that the predominant FA are C16:0, C16:1, C18:1, and C18:2. Of these, C18:1, i.e., oleic acid, is predominant. Oleic acid and C18:2, i.e., linoleic acid, decrease substantially at high AD percentages. In most vegetable and seed oils C18:2 is higher than C18:1, while in Chlorella sorokiniana at 0% and 15% AD, the C18:1/C18:2 ratio is about 2 at 30% AD and increases to about 4 at 50% AD. Additionally, palmitic acid, C16:0, is present in significant amounts up to 15% AD but decreases substantially at higher AD percentages. It can also be seen that chain lengths up to C26 are obtained, and in general increasing the amount of AD in the growth medium results in FA with higher chain length. FA with chain lengths higher than C20 are absent at 0% AD, but increase in chain length progressively as the percentage of AD is increased.
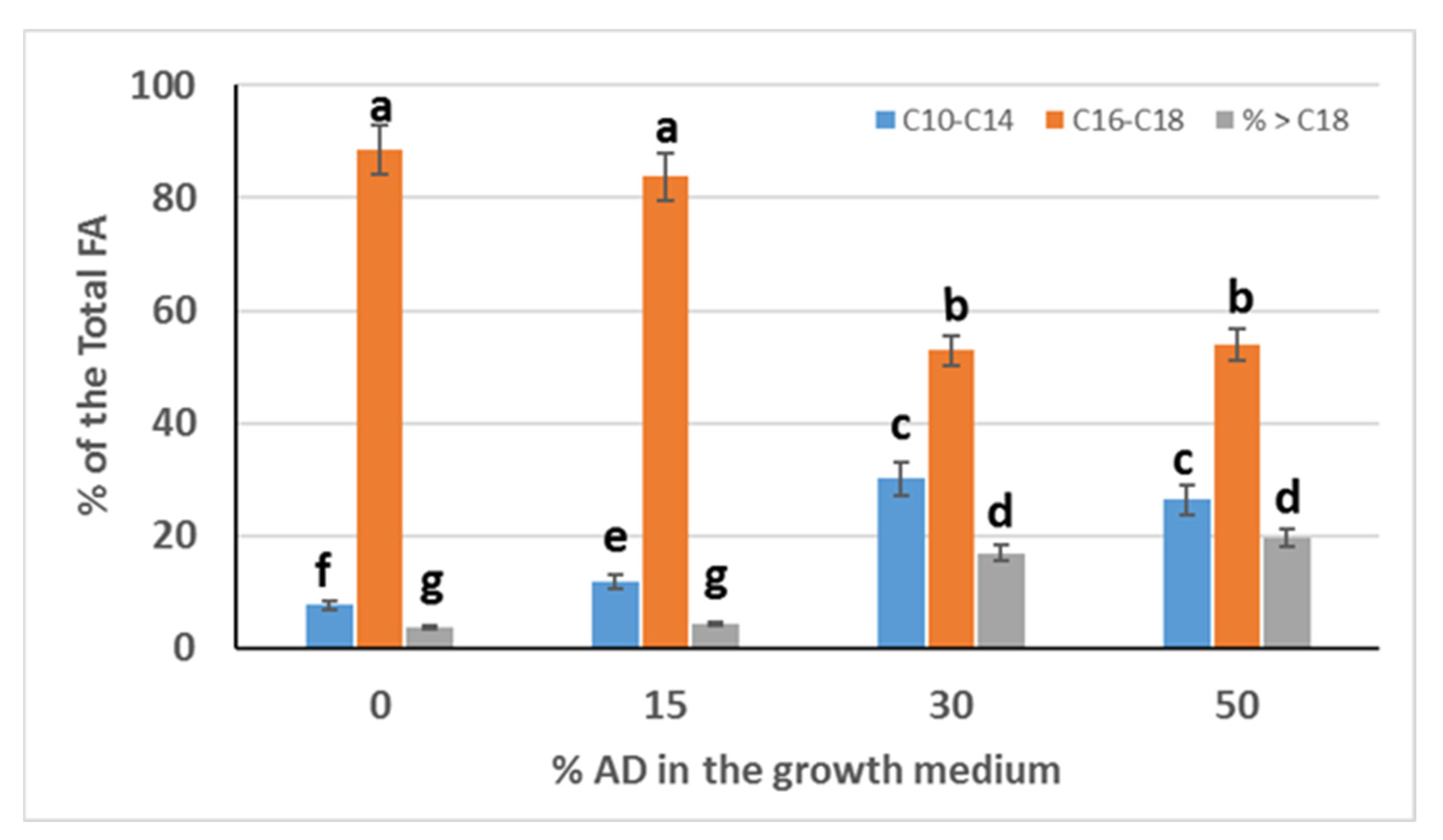
If the fatty acids are separated in three categories as: short chain FA from C10 to C14, medium chain FA from C16 to C18, and long chain FA higher than C18, then
Figure 4 shows a histogram of the distribution of short, medium, and long chain FA versus the percentage of AD used. It was observed that the use of AD has a profound effect on the chain length of the FA in the biomass of
Chlorella sorokiniana. In particular, the percentage of short chain length FA of the total FA increased by the addition of 15%, 30%, and 50% of AD in the growth medium compared with the control (crude glycerol + 0% AD). In contrast, the percentage of medium chain FA of the total FA decreased when AD was added to the growth medium at 30% and 50% compared with the control, while by the addition of 15% AD in the growth medium, the percentage of medium chain FA did not show statistically significant differences compared with the control. Additionally, the percentage of long chain FA of the total FA increased by the addition of 30% and 50% of AD in the growth medium compared with the control, while by the addition of 15% AD in the growth medium, the percentage of long chain FA did not show statistically significant differences compared with the control (
Figure 4). The medium chain length FA, so predominant up to 15% AD, decrease significantly from 85–89% of the total FA to 52–53% at 30% and 50% AD. Instead, at 30% and 50% AD, the percentage of short chain and long chain FA increases substantially from 7.6% to 30.16%, respectively. Long chain FA also increase from about 4% to about 17% at 30% AD and to about 20% at 50% AD. This re-arrangement of the distribution of the chain length of the FA affects the properties of the biodiesel as discussed in the following paragraph.
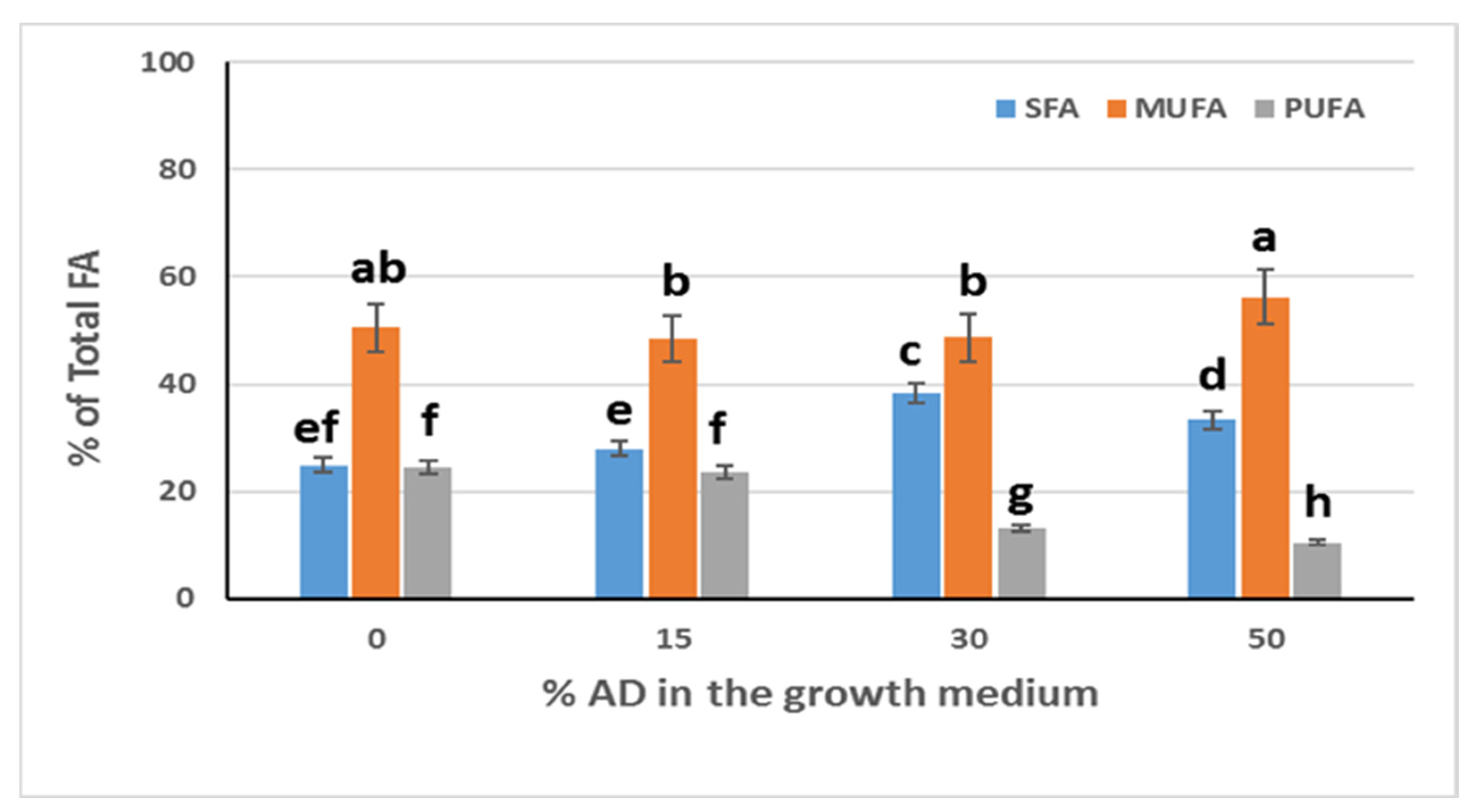
Figure 5 shows the distribution of SFA (Saturated FA), MUFA (Mono-Unsaturated FA), and PUFA (Poly-Unsaturated FA) versus the percentage of AD added to the growth medium. From
Figure 5, it is evident that the percentage of AD also has an effect on the relative distribution of SFA, MUFA, and PUFA. In particular, the percentage of SFA of the total FA increased by the addition of 30% and 50% of AD in the growth medium compared with the control (crude glycerol + 0% AD), while by the addition of 15% AD in the growth medium, the percentage of SFA did not show statistically significant differences compared with the control. The percentage of MUFA of the total FA by the addition of 15%, 30%, and 50% of AD in the growth medium did not show statistically significant differences compared with the control. Furthermore, the percentage of PUFA of the total FA decreased by the addition of 30% and 50% of AD in the growth medium compared with the control, while by the addition of 15% AD in the growth medium, the percentage of PUFA did not show statistically significant differences compared with the control.
This should affect the biodiesel properties. Increasing the degree of saturation leads to a decrease in ignition delay [
32]. The ignition delay is related to the cetane number (CN). As the ignition delay increases, the CN number decreases; the CN number is an indication of the quality of the biodiesel produced [
20]. The ignition delay is an important parameter which characterizes the initiation of combustion process and consequently its development in diesel engines. Diesel engines, compared to gasoline engines, typically produce lower carbon monoxide and partially burned hydrocarbons. However, nitric oxides and particulate matter are significant pollutants from diesel engines. Changes in ignition delay affect the exhaust emissions indirectly. A decrease in the ignition delay leads to a longer time for complete combustion to take place in the cylinder and this results in lower emissions. Reversely, increased ignition delay leads to insufficient combustion time and therefore to higher exhaust emission. Therefore, ignition delay affects the combustion performance of a diesel engine. Higher CN leads to better engine performance. Additionally, the degree of saturation affects the iodine number. As the degree of saturation increases the IV number decreases. However, low iodine values lead to biodiesel, which is more combustible but not suitable for colder climates as it has rather poor flow properties at low temperatures [
32].
3.5. Fatty Acid Methyl Ester (FAME) Properties
The saponification number (SN) is the weight (mg) of potassium hydroxide that is needed to convert one gram of fatty acid into soap. It relates to the average molecular weight, or the average chain length of all fatty acids present in the bio-oil. Therefore, longer chain FA give lower SN and vice versa. As the chain length becomes longer, the ratio of the mass of carboxylate (1) to the rest of the molecule of the FA decreases, hence less KOH is needed per g of FA.
The iodine value (IV) is the amount of iodine in grams absorbed by 100 mL of FA or FAME and is a measure of the degree of unsaturation of the FAs present in the oil or the respective FAME. Unsaturated fatty acid methyl esters are needed, as they improve the cold properties of the biodiesel, that is they prevent its solidification at low temperatures. As the degree of unsaturation increases, the IV of the biodiesel increases as well. However, a degree of unsaturation in the biodiesel leads to unwanted reactions with atmospheric oxygen leading to the formation of polymerization products in the internal combustion engine [
20]. Therefore, a higher IV equal to 120 g I/100 g FAME has been set.
The cetane number (CN) relates to the quality of the fuel. As the cetane number increases the ignition delay, which the time between the injection of the fuel into the cylinder and the onset of ignition, decreases. Therefore, a higher CN leads to a shorter ignition delay. A minimum CN equal to 51 has been set by EN standards. The CN value depends on the number of unsaturated fatty acids, the number of double bonds, and the molecular weight of the FAME. Therefore, it correlates with the IV and the SN. The CN should not be very high as, as it increases, the IV and subsequently the degree of unsaturation decrease, and this may lead to poor cold flow properties of the biodiesel. Therefore, a maximum value for CN has been set to 65 by the ASTEM standard.
Table 4 shows the SN, IV, CN, and HHV values of the FAME, obtained from
Table 3 of the bio-oil extracted from the biomass of
Chlorella sorokiniana cultivated with 0%, 15%, 30%, and 50% AD, using Equations (1)–(4). Values of FAME properties in
Table 4 can be explained in terms of the FA distribution with respect to the chain length (
Figure 4) and the degree of saturation (
Figure 5). Increasing the degree of saturation leads to a decrease in the iodine number (IV). Additionally, increasing the average chain length decreases both the iodine number (IV) and the saponification number (SN) and increases the cetane number (CN). However, according to Equation (3), a decrease in the SN has more profound effect on the CN than a decrease in IV. The combined results showed that the SN, HHV, and CN values of the FAME by the addition of 15%, 30%, and 50% of AD in the growth medium did not show statistically significant differences compared with the control (crude glycerol + 0% AD), while increasing the percentage of AD in the growth medium the CN values show an increase trend from 52.68 to 56.12. The IV values of the FAME decreased by the addition of 30% and 50% of AD in the growth medium compared with the control while, by the addition of 15% AD in the growth medium, the IV values of the FAME did not show statistically significant differences compared with the control (
Table 4).
4. Conclusions
Chlorella sorokiniana was cultivated heterotrophically using glycerol and varying amounts of AD such as 0%, 15%, 30%, and 50%, respectively. Glycerin, a biodiesel by-product, seems to be a potential substrate for the heterotrophic cultivation of Chlorella sorokiniana, while the AD can be an additional source of carbon for heterotrophic or mixotrophic growth.
These results indicate that different addition rates of AD in the growth medium affect the rate of carbon uptake up by the Chlorella sorokiniana cells, as well as its protein and lipid content. The maximum carbon uptake rate occurs at about 30% AD. Increasing the percentage of AD from 0% to 50% in the growth medium, the lipid content gradually decreases by up to 5%. This decrease can be ameliorated or reversed by other modes of cultivation, based of biochemical engineering principles, such as fed-batch mode rather than batch cultivation.
Additionally, the different addition rates of AD in the growth medium affect the FA distribution in the oil extracted and the FAME properties. FA distribution ranged from C10 to C26. Increasing the percentage of AD from 0% to 50% increased the percentage of the FA with C < 16 from 7.6 to 26.4% of total FA and the percentage of the FA with C > 18 from 3.8 to 19.7% of total FA. On the contrary, the percentage of the FA with C16–C18 decreased from 88.6% to 53.9% of total FA. In addition, increasing the percentage of AD from 0% to 50% increased the percentage of SFA by 33% and decreased the percentage of PUFA by 57%. In all AD percentages, the percentage of MUFA did not vary significantly and ranged from 48% to 56% of the total FA. The changes in the FA distribution as the percentage of AD increases from 0% to 50% in the growth medium, lead to increase cetane numbers by 6.5% and decrease iodine values by 20%. AD is a useful raw material for the formulation of growth media for the cultivation of Chlorella sorokiniana.
These results could stimulate the scientific community in order to use anaerobic digestate, enriched with carbon sources other than glycerol, into the formulation of growth media for the cultivation of microalgae and investigate growth kinetics and the biodiesel properties. Further research is needed in order to determine the effect of other modes of cultivation using glycerol and AD in microalgal cultivation for producing lipids for biodiesel production. As different strains of microalgae can utilize glycerol in varying degrees, glycerol and AD can be tried on different strains, especially genetically modified strains.



 and
and 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}