Potential of High-Resolution Pléiades Imagery to Monitor Salt Marsh Evolution After Spartina Invasion

 , , , ,
, , , ,  ,
, 
Abstract
:
1. Introduction
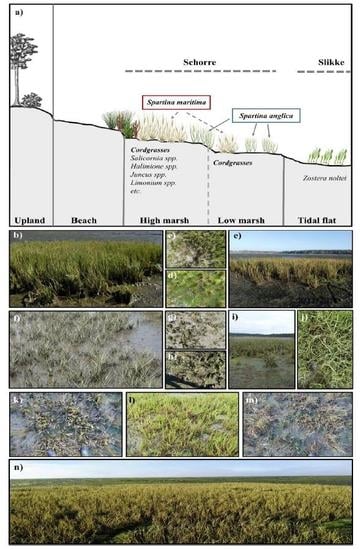
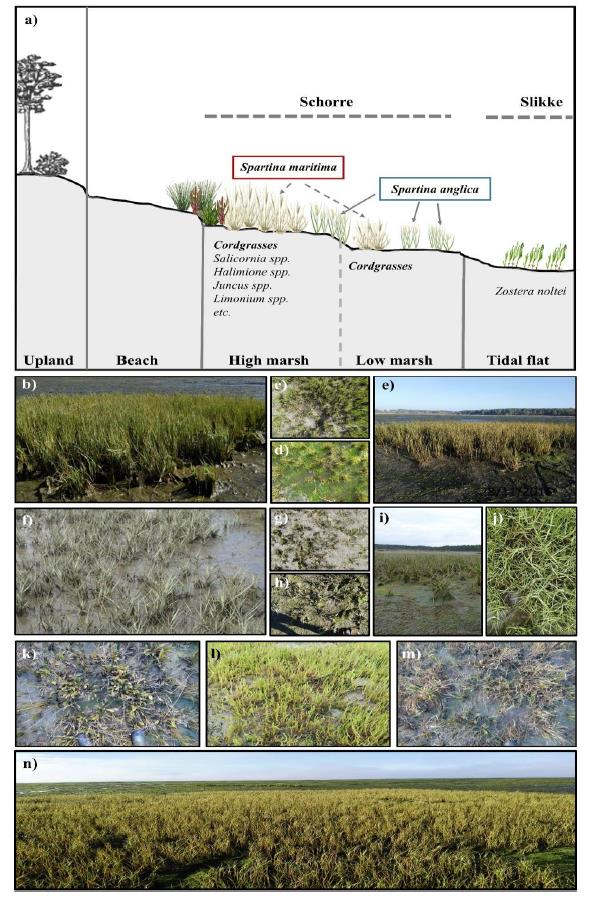
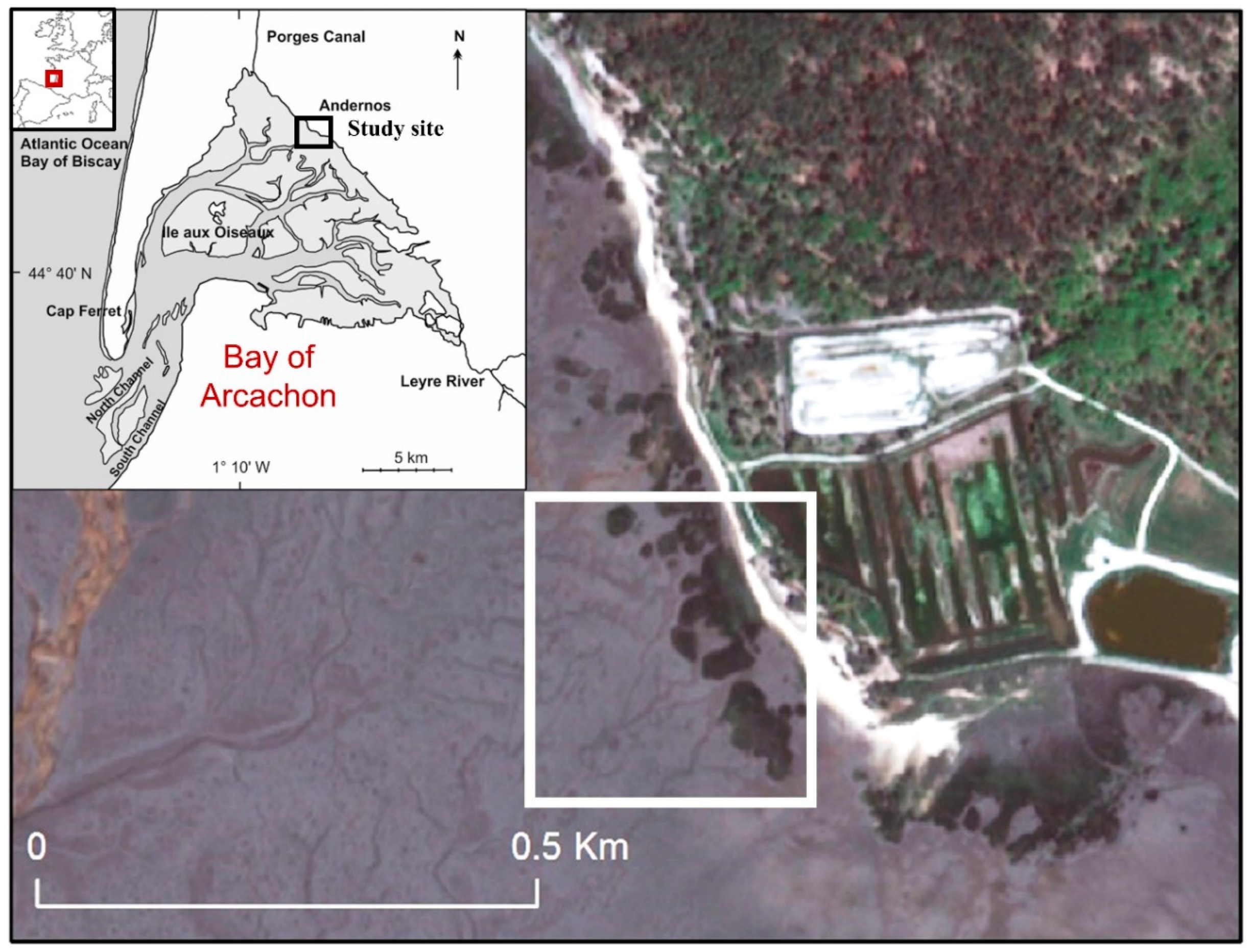
2. Study Site
Vegetation in the Bay of Arcachon
3. Datasets and Methods
3.1. Aerial Photographs
3.2. GNSS Data
3.3. Radiometric Measurements
3.4. High Resolution Pléiades Images
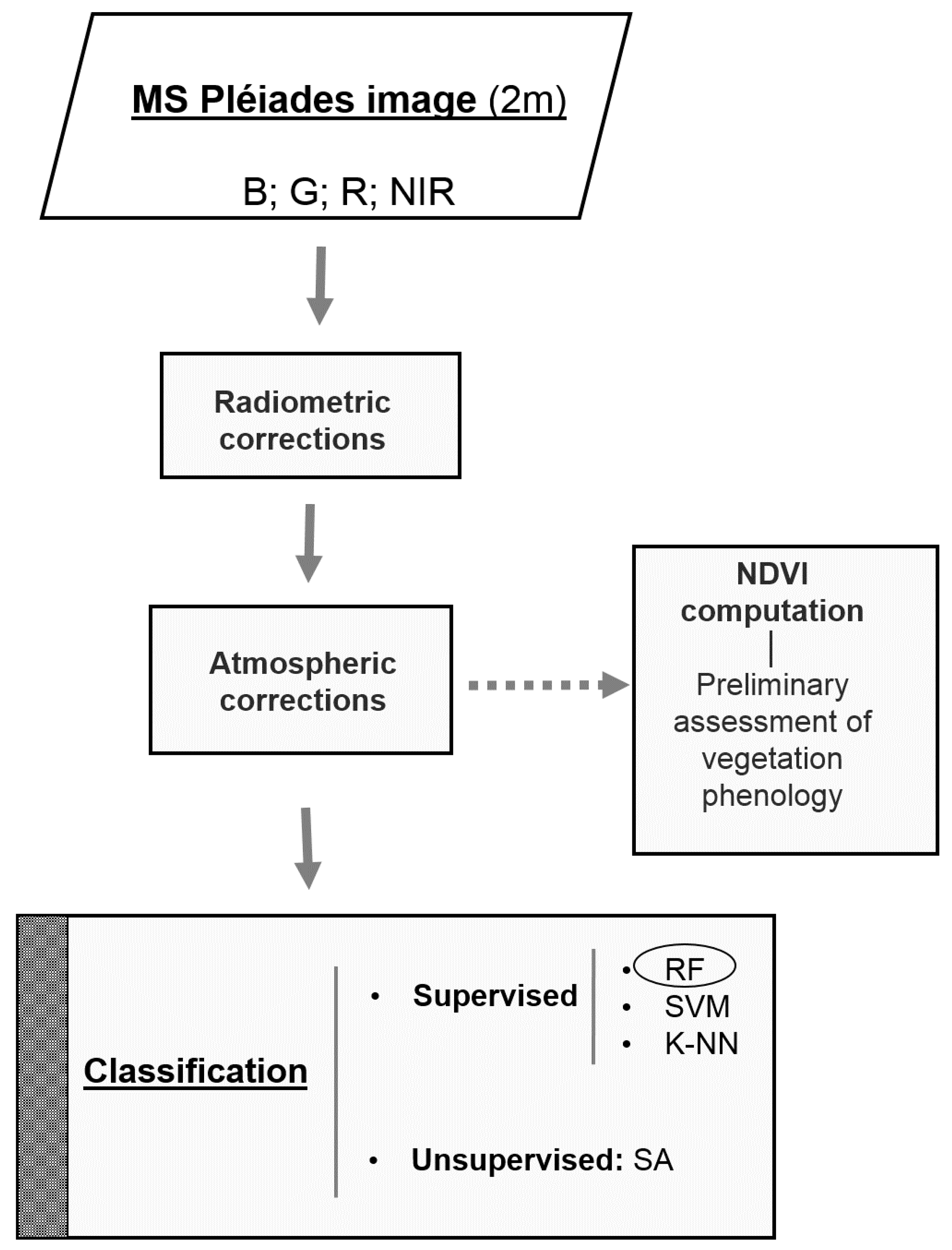
3.5. Pre-processing of the Pléiades Images
3.6. Pixel-based Classification
4. Results and Discussion
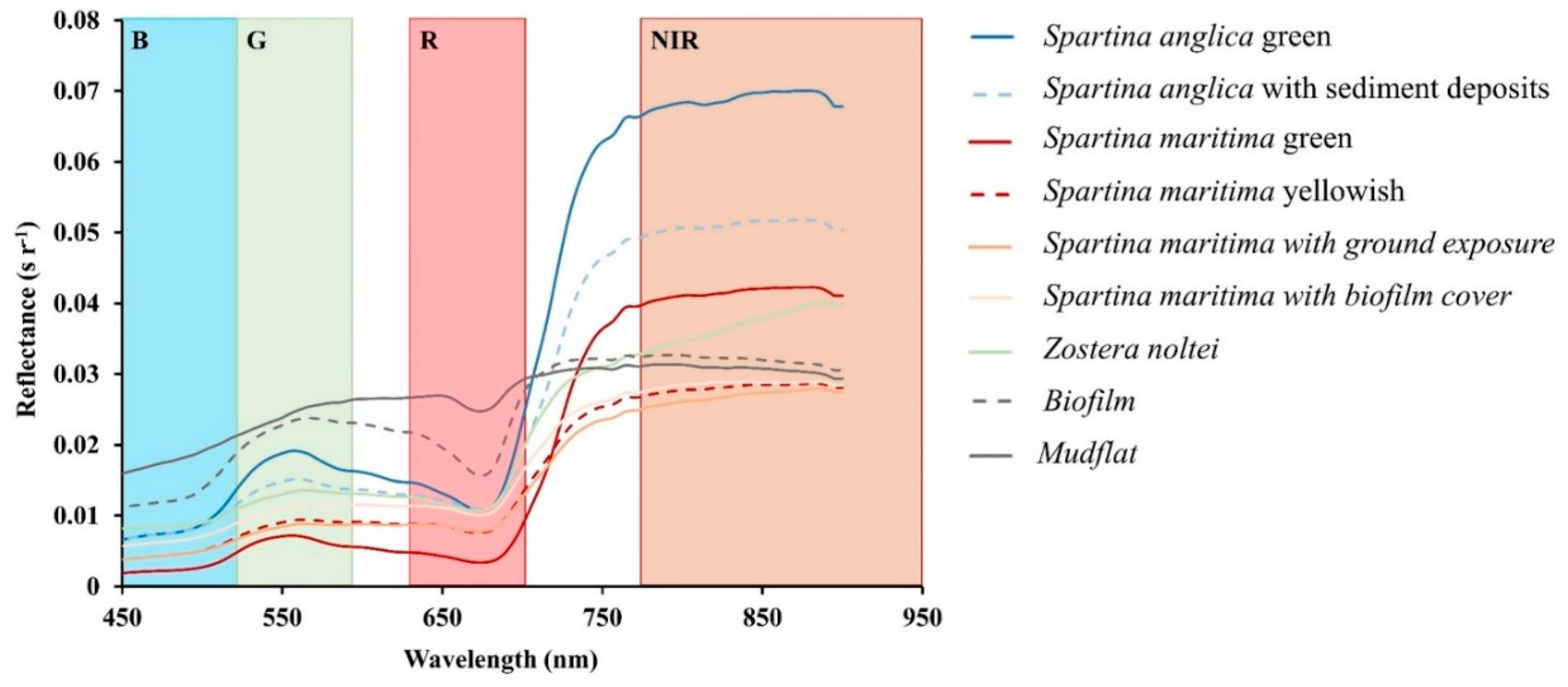
4.1. Spectral Signature of Vegetated Structures
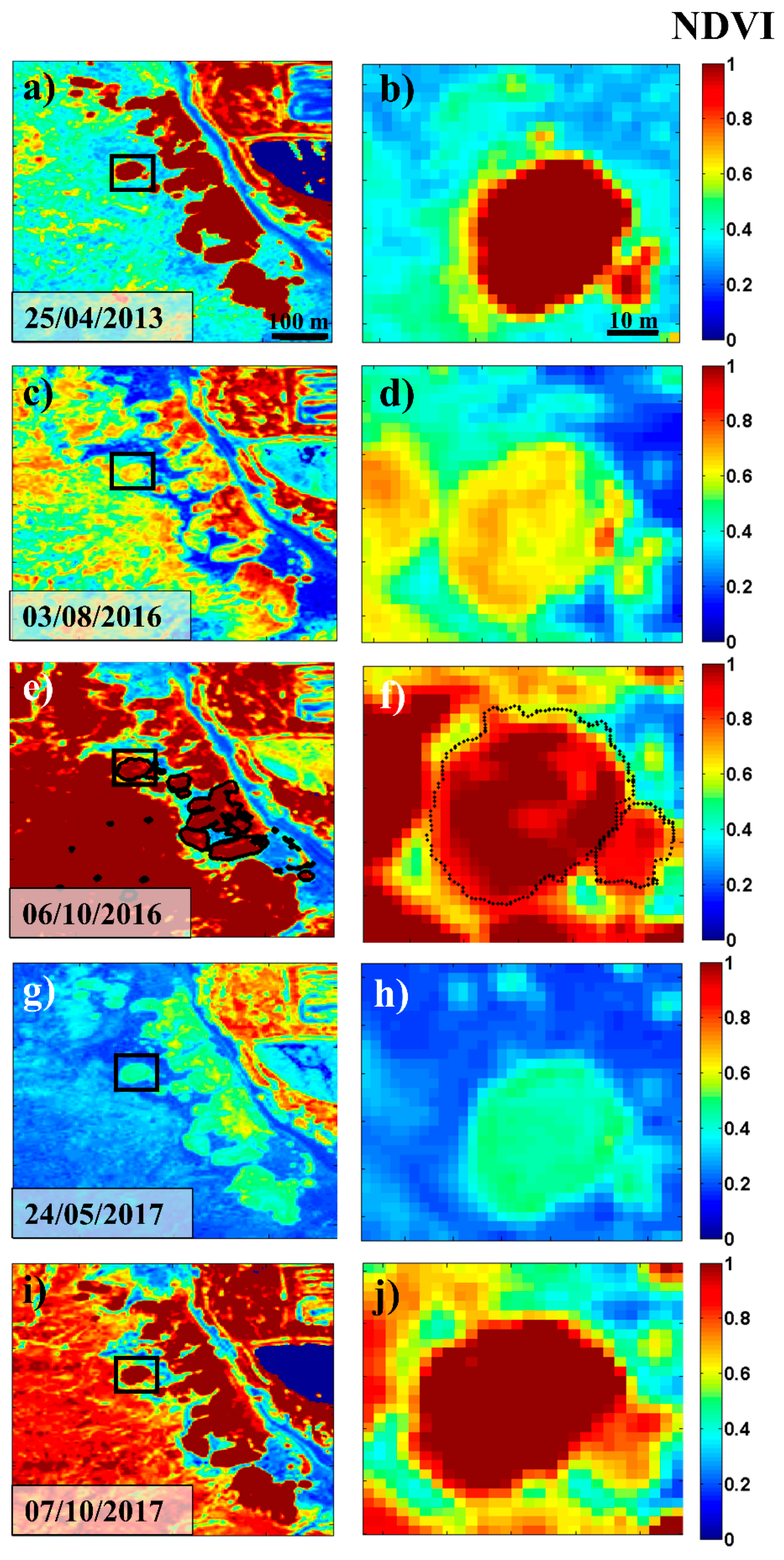
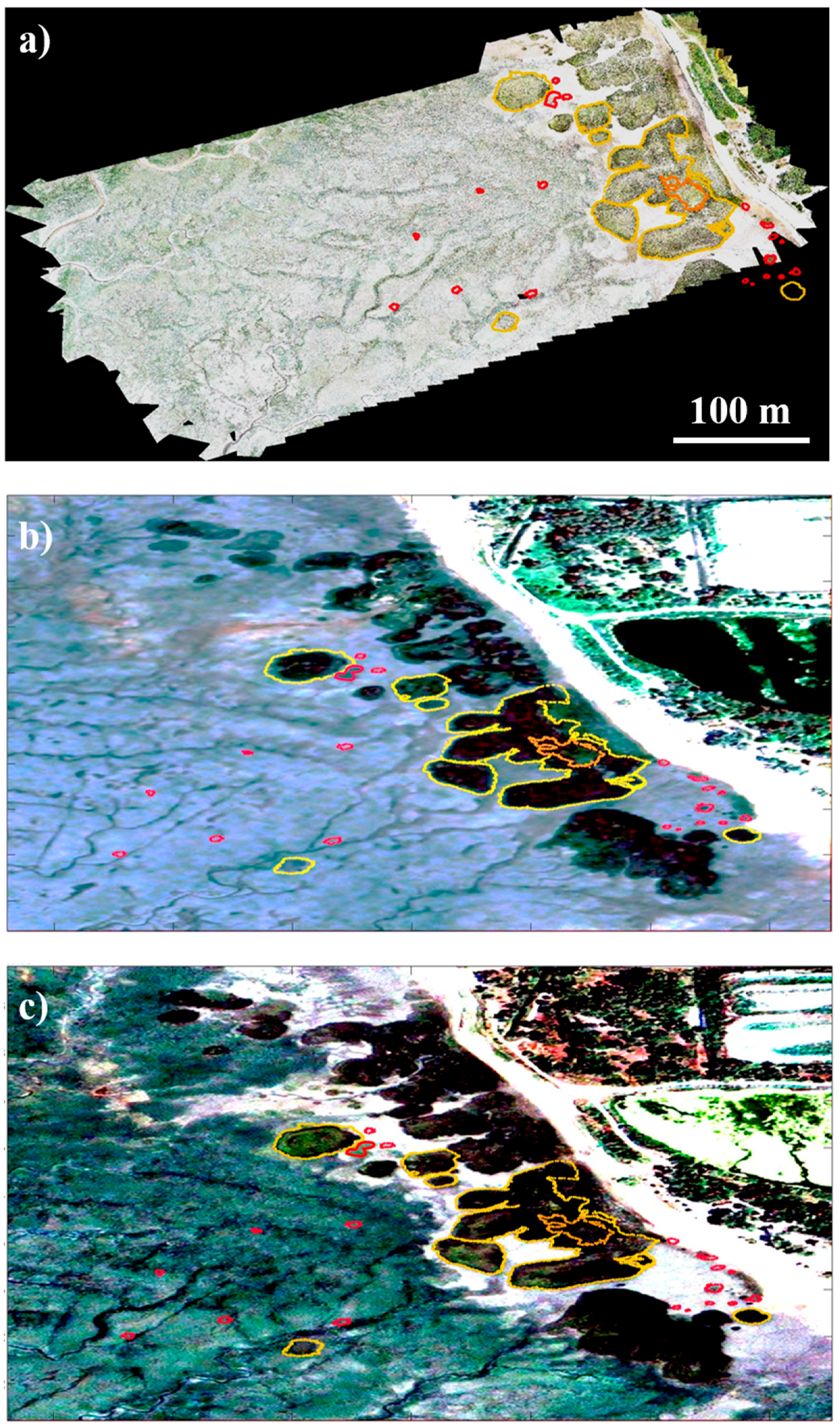
4.2. Long-Term Evolution of the High Marsh Zone and Ground Truth Data Validation
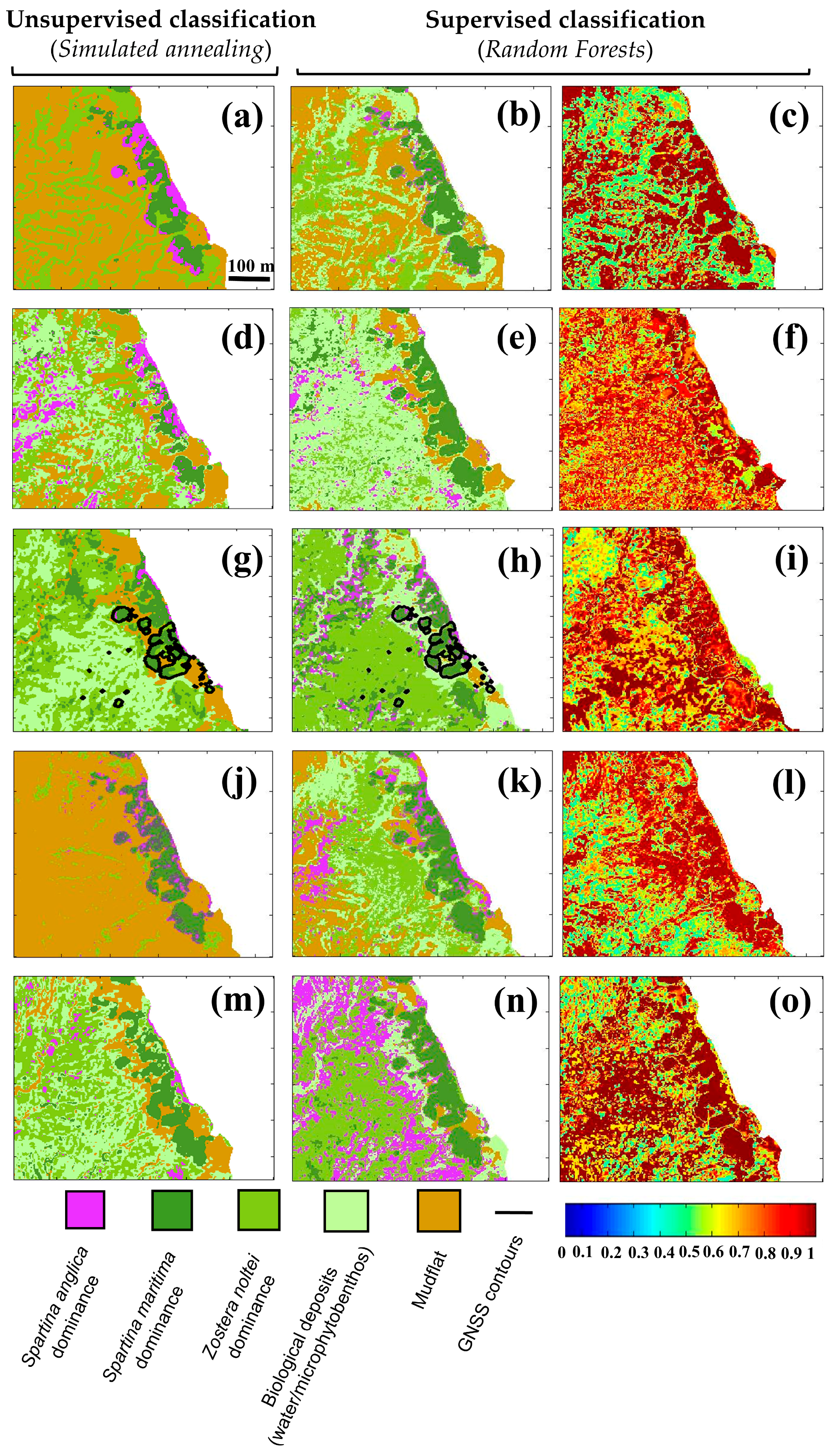
4.3. Pixel Classification Using Unsupervised and Supervised Methods
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
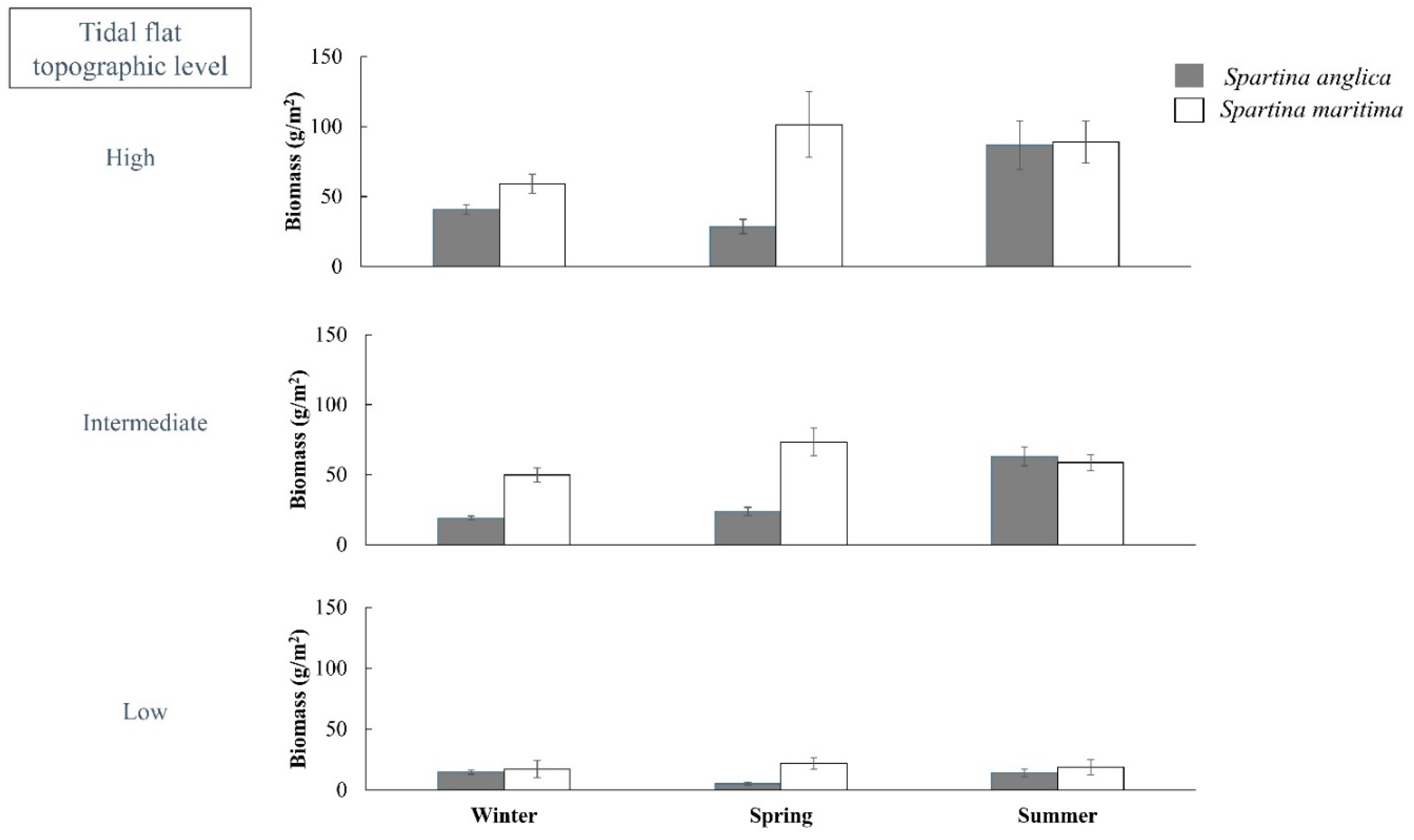
Appendix A—Quarterly Monitoring of Spartina anglica and Spartina maritima Biomass in the Bay of Arcachon between 2014 and 2015.

References
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human Domination of Earth’s Ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef]
- Davis, M.A. Biotic Globalization: Does Competition from Introduced Species Threaten Biodiversity? BioScience 2003, 53, 481. [Google Scholar] [CrossRef] [Green Version]
- Seabloom, E.W.; Ruggiero, P.; Hacker, S.D.; Mull, J.; Zarnetske, P. Invasive grasses, climate change, and exposure to storm-wave overtopping in coastal dune ecosystems. Glob. Chang. Biol. 2013, 19, 824–832. [Google Scholar] [CrossRef]
- Simberloff, D.; Martin, J.-L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M.; et al. Impacts of biological invasions: what’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Crooks, J.A. Characterizing ecosystem-level consequences of biological invasions: The role of ecosystem engineers. Oikos 2002, 97, 153–166. [Google Scholar] [CrossRef]
- Zarnetske, P.L.; Seabloom, E.W.; Hacker, S.D. Non-target effects of invasive species management: Beachgrass, birds, and bulldozers in coastal dunes. Ecosphere 2010, 1, art13. [Google Scholar] [CrossRef]
- Schlaepfer, M.A.; Sax, D.F.; Olden, J.D. The potential conservation value of non-native species. Conserv. Biol. J. Soc. Conserv. Biol. 2011, 25, 428–437. [Google Scholar] [CrossRef]
- Strayer, D.L.; Eviner, V.T.; Jeschke, J.M.; Pace, M.L. Understanding the long-term effects of species invasions. Trends Ecol. Evol. 2006, 21, 645–651. [Google Scholar] [CrossRef]
- Bustamante, J.; Aragonés, D.; Afán, I.; Luque, C.J.; Pérez-Vázquez, A.; Castellanos, E.M.; Díaz-Delgado, R. Hyperspectral Sensors as a Management Tool to Prevent the Invasion of the Exotic Cordgrass Spartina densiflora in the Doñana Wetlands. Remote Sens. 2016, 8, 1001. [Google Scholar] [CrossRef]
- Correll, M.D.; Hantson, W.; Hodgman, T.P.; Cline, B.B.; Elphick, C.S.; Shriver, W.G.; Tymkiw, E.L.; Olsen, B.J. Fine-Scale Mapping of Coastal Plant Communities in the Northeastern USA. Wetlands 2018, 1–12. [Google Scholar] [CrossRef]
- Hacker, S.D.; Heimer, D.; Hellquist, C.E.; Reeder, T.G.; Reeves, B.; Riordan, T.J.; Dethier, M.N. A Marine Plant (Spartina Anglica) Invades Widely Varying Habitats: Potential Mechanisms of Invasion and Control. Biol. Invasions 2001, 3, 211–217. [Google Scholar] [CrossRef]
- Early, R.; Bradley, B.A.; Dukes, J.S.; Lawler, J.J.; Olden, J.D.; Blumenthal, D.M.; Gonzalez, P.; Grosholz, E.D.; Ibañez, I.; Miller, L.P.; et al. Global threats from invasive alien species in the twenty-first century and national response capacities. Nat. Commun. 2016, 7, 12485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marani, M.; Lanzoni, S.; Silvestri, S.; Rinaldo, A. Tidal landforms, patterns of halophytic vegetation and the fate of the lagoon of Venice. J. Mar. Syst. 2004, 51, 191–210. [Google Scholar] [CrossRef]
- Silvestri, S.; Marani, M.; Marani, A. Hyperspectral remote sensing of salt marsh vegetation, morphology and soil topography. Phys. Chem. Earth Parts ABC 2003, 28, 15–25. [Google Scholar] [CrossRef]
- Belluco, E.; Camuffo, M.; Ferrari, S.; Modenese, L.; Silvestri, S.; Marani, A.; Marani, M. Mapping salt-marsh vegetation by multispectral and hyperspectral remote sensing. Remote Sens. Environ. 2006, 105, 54–67. [Google Scholar] [CrossRef]
- Klemas, V. Remote Sensing of Wetlands: Case Studies Comparing Practical Techniques. J. Coast. Res. 2011, 418–427. [Google Scholar]
- Thomson, A.G.; Huiskes, A.; Cox, R.; Wadsworth, R.A.; Boorman, L.A. Short-term vegetation succession and erosion identified by airborne remote sensing of Westerschelde salt marshes, The Netherlands. Int. J. Remote Sens. 2004, 25, 4151–4176. [Google Scholar] [CrossRef]
- Barillé, L.; Robin, M.; Harin, N.; Bargain, A.; Launeau, P. Increase in seagrass distribution at Bourgneuf Bay (France) detected by spatial remote sensing. Aquat. Bot. 2010, 92, 185–194. [Google Scholar] [CrossRef]
- Zuo, P.; Zhao, S.; Liu, C.; Wang, C.; Liang, Y. Distribution of Spartina spp. along China’s coast. Ecol. Eng. 2012, 40, 160–166. [Google Scholar] [CrossRef]
- Zhang, R.S.; Shen, Y.M.; Lu, L.Y.; Yan, S.G.; Wang, Y.H.; Li, J.L.; Zhang, Z.L. Formation of Spartina alterniflora salt marshes on the coast of Jiangsu Province, China. Ecol. Eng. 2004, 23, 95–105. [Google Scholar] [CrossRef]
- Li, J.; Gao, S.; Wang, Y. Invading cord grass vegetation changes analyzed from Landsat-TM imageries: A case study from the Wanggang area, Jiangsu coast, eastern China. Acta Oceanol. Sin. 2010, 29, 26–37. [Google Scholar] [CrossRef]
- Ouyang, Z.-T.; Gao, Y.; Xie, X.; Guo, H.-Q.; Zhang, T.-T.; Zhao, B. Spectral Discrimination of the Invasive Plant Spartina alterniflora at Multiple Phenological Stages in a Saltmarsh Wetland. PLoS ONE 2013, 8, e67315. [Google Scholar] [CrossRef] [PubMed]
- Chapman, V.J. Coastal Vegetation; Pergamon Press: Oxford, UK; Macmillan: New York, NY, USA, 1964. [Google Scholar]
- Pennings, S.C.; Callaway, R.M. Salt Marsh Plant Zonation: The Relative Importance of Competition and Physical Factors. Ecology 1992, 73, 681–690. [Google Scholar] [CrossRef]
- Adam, E.; Mutanga, O.; Rugege, D. Multispectral and hyperspectral remote sensing for identification and mapping of wetland vegetation: A review. Wetl. Ecol. Manag. 2010, 18, 281–296. [Google Scholar] [CrossRef]
- Silvestri, S.; Marani, M.; Settle, J.; Benvenuto, F.; Marani, A. Salt marsh vegetation radiometry: Data analysis and scaling. Remote Sens. Environ. 2002, 80, 473–482. [Google Scholar] [CrossRef]
- Clint Slatton, K.; Crawford, M.M.; Chang, L.-D. Modeling temporal variations in multipolarized radar scattering from intertidal coastal wetlands. ISPRS J. Photogramm. Remote Sens. 2008, 63, 559–577. [Google Scholar] [CrossRef]
- Sun, C.; Liu, Y.; Zhao, S.; Zhou, M.; Yang, Y.; Li, F. Classification mapping and species identification of salt marshes based on a short-time interval NDVI time-series from HJ-1 optical imagery. Int. J. Appl. Earth Obs. Geoinf. 2016, 45, 27–41. [Google Scholar] [CrossRef]
- Mouw, C.B.; Greb, S.; Aurin, D.; DiGiacomo, P.M.; Lee, Z.; Twardowski, M.; Binding, C.; Hu, C.; Ma, R.; Moore, T.; et al. Aquatic color radiometry remote sensing of coastal and inland waters: Challenges and recommendations for future satellite missions. Remote Sens. Environ. 2015, 160, 15–30. [Google Scholar] [CrossRef]
- Hill, V.J.; Zimmerman, R.C.; Bissett, W.P.; Dierssen, H.; Kohler, D.D.R. Evaluating Light Availability, Seagrass Biomass, and Productivity Using Hyperspectral Airborne Remote Sensing in Saint Joseph’s Bay, Florida. Estuaries Coasts 2014, 37, 1467–1489. [Google Scholar] [CrossRef]
- Garzelli, A.; Nencini, F.; Alparone, L.; Aiazzi, B.; Baronti, S. Pan-sharpening of multispectral images: A critical review and comparison. In Proceedings of the 2004 IEEE International Geoscience and Remote Sensing Symposium, Anchorage, AK, USA, 20–24 September 2004; Volume 1, p. 84. [Google Scholar]
- Akula, R.; Gupta, R.; Devi, M.R.V. An efficient PAN sharpening technique by merging two hybrid approaches. Procedia Eng. 2012, 30, 535–541. [Google Scholar] [CrossRef] [Green Version]
- Carter, G.A.; Lucas, K.L.; Blossom, G.A.; Lassitter, C.L.; Holiday, D.M.; Mooneyhan, D.S.; Fastring, D.R.; Holcombe, T.R.; Griffith, J.A. Remote Sensing and Mapping of Tamarisk along the Colorado River, USA: A Comparative Use of Summer-Acquired Hyperion, Thematic Mapper and QuickBird Data. Remote Sens. 2009, 1, 318–329. [Google Scholar] [CrossRef] [Green Version]
- Zak, M.R.; Cabido, M. Spatial patterns of the Chaco vegetation of central Argentina: Integration of remote sensing and phytosociology. Appl. Veg. Sci. 2002, 5, 213–226. [Google Scholar]
- Allard, J.; Chaumillon, E.; Féniès, H. A synthesis of morphological evolutions and Holocene stratigraphy of a wave-dominated estuary: The Arcachon lagoon, SW France. Cont. Shelf Res. 2009, 29, 957–969. [Google Scholar] [CrossRef]
- Cayocca, F. Long-term morphological modeling of a tidal inlet: the Arcachon Basin, France. Coast. Eng. 2001, 42, 115–142. [Google Scholar] [CrossRef]
- Plus, M.; Dumas, F.; Stanisiere, J.-Y.; Maurer, D. Hydrodynamic characterization of the Arcachon Bay, using model-derived descriptors. Cont. Shelf Res. 2009, 29, 1008–1013. [Google Scholar] [CrossRef]
- Baumel, A.; Ainouche, M.L.; Levasseur, J.E. Molecular investigations in populations of Spartina anglica C.E. Hubbard (Poaceae) invading coastal Brittany (France). Mol. Ecol. 2001, 10, 1689–1701. [Google Scholar] [CrossRef]
- Soriano-Sierra, E.J. Etude Ecologique des Marais Sales du Bassin d’Arcachon: Structure et Evolution de Schorres, Production et Dégradation de leur Végétation et Echanges de Matières Particulaires entre les Schorres et le Bassin 1992 (Doctoral dissertation). Available online: http://www.theses.fr/1992BOR10565 (accessed on 14 March 2019).
- Labourg, P.J.; Soriano-Sierra, E.-J.; Auby, I. Evolution récente de la végétation intertidale du Delta de l’Eyre. In Proceedings of the Travaux et coloques Scientifiques, Le Delta de La Leyre, Le Teich, France, 21–23 October 1993. [Google Scholar]
- Pierce, S.M. Estimation of the non-seasonal production of Spartina maritima (Curtis) Fernald in a South African Estuary. Estuar. Coast. Shelf Sci. 1983, 16, 241–254. [Google Scholar] [CrossRef]
- Proença, B.; Nez, T.; Poli, A.; Ciutat, A.; Devaux, L.; Sottolichio, A.; de Montaudouin, X.; Michalet, R. Intraspecific facilitation explains the spread of the invasive engineer Spartina anglica in Atlantic salt marshes. J. Veg. Sci. 2019, 30, 212–223. [Google Scholar] [CrossRef]
- Auby, I.; Labourg, P.-J. Seasonal dynamics of Zostera noltii hornem. In the Bay of Arcachon (France). J. Sea Res. 1996, 35, 269–277. [Google Scholar] [CrossRef]
- Castel, J.; Caumette, P.; Herbert, R. Eutrophication gradients in coastal lagoons as exemplified by the Bassin d’Arcachon and the Etang du Prévost. Hydrobiologia 1996, 329, ix–xxviii. [Google Scholar] [CrossRef]
- Takasu, T. RTKLIB: Open source program package for RTK-GPS. In Proceedings of the FOSS4G, Tokyo, Japan, 2 November 2009. [Google Scholar]
- Takasu, T.; Yasuda, A. Development of the low-cost RTK-GPS receiver with an open source program package RTKLIB. In Proceedings of the International Symposium on GPS/GNSS, Jeju, Korea, 4–6 November 2009; pp. 4–6. [Google Scholar]
- Frappart, F.; Roussel, N.; Darrozes, J.; Bonneton, P.; Bonneton, N.; Detandt, G.; Perosanz, F.; Loyer, S. High rate GNSS measurements for detecting non-hydrostatic surface wave. Application to tidal bore in the Garonne River. Eur. J. Remote Sens. 2016, 49, 917–932. [Google Scholar] [CrossRef] [Green Version]
- Duggin, M.J.; Philipson, W.R. Field measurement of reflectance: Some major considerations. Appl. Opt. 1982, 21, 2833–2840. [Google Scholar] [CrossRef]
- Inglada, J.; Christophe, E. The Orfeo Toolbox remote sensing image processing software. In Proceedings of the 2009 IEEE International Geoscience and Remote Sensing Symposium, Cape Town, South Africa, 12–17 July 2009; Volume 4, pp. IV-733–IV-736. [Google Scholar]
- Bru, D.; Lubac, B.; Normandin, C.; Robinet, A.; Leconte, M.; Hagolle, O.; Martiny, N.; Jamet, C. Atmospheric correction of multi-spectral littoral images using a PHOTONS/AERONET-based regional aerosol model. Remote Sens. 2017, 9, 814. [Google Scholar] [CrossRef]
- Rouse, J.W., Jr.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring Vegetation Systems in the Great Plains with ERTS; NASA: Washington, DC, USA, 1974; pp. 309–317.
- Hutchinson, C.F. Land remote sensing technology and methodology. Clim. Chang. 1986, 9, 149–166. [Google Scholar] [CrossRef]
- Bertsimas, D.; Tsitsiklis, J. Simulated Annealing. Stat. Sci. 1993, 8, 10–15. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Liu, Y.; Zhao, S.; Li, H.; Sun, J. Saltmarshes Response to Human Activities on a Prograding Coast Revealed by a Dual-Scale Time-Series Strategy. Estuaries Coasts 2017, 40, 522–539. [Google Scholar] [CrossRef]
- Sun, C.; Fagherazzi, S.; Liu, Y. Classification mapping of salt marsh vegetation by flexible monthly NDVI time-series using Landsat imagery. Estuar. Coast. Shelf Sci. 2018, 213, 61–80. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Satellite | Acquisition Date | Season | Acquisition Time (UTC) | Time of Low Tide (UTC) |
|---|---|---|---|---|
| Pléiades-1A | 25/04/2013 | Spring | 11h15 | 12h04 |
| Pléiades-1A | 03/08/2016 | Summer | 11h15 | 12h45 |
| Pléiades-1A | 06/10/2016 | Autumn | 11h23 | 14h59 |
| Pléiades-1B | 24/05/2017 | Spring | 11h04 | 11h06 |
| Pléiades-1A | 07/10/2017 | Autumn | 11h08 | 13h27 |
| Spectral Band (nm) | Field Rrs (sr−1) | Field Rrs_SRF (sr−1) | Pléiades Rrs (sr−1) | ||||
|---|---|---|---|---|---|---|---|
| S. Maritima. | S. Anglica | S. Maritima | S. Anglica | S. Maritima. | S. Anglica | ||
| Green | 560 500–620 | 0.007 ± 0.002 - | 0.019 ± 0.003 - | - 0.006 ± 0.004 | - 0.017 ± 0.004 | - 0.015 ± 0.001 | - 0.018 ± 0.001 |
| Red | 650 590–710 | 0.004 ± 0.002 - | 0.013 ± 0.004 - | - 0.005 ± 0.006 | - 0.014 ± 0.005 | - 0.007 ± 0.001 | - 0.009 ± 0.001 |
| NIR | 840 740–940 | 0.042 ± 0.008 - | 0.071 ± 0.007 - | - 0.039 ± 0.009 | - 0.064 ± 0.010 | - 0.029 ± 0.003 | - 0.034 ± 0.003 |
| NDVI | 0.83 ± 0.03 | 0.69 ± 0.02 | 0.77 ± 0.05 | 0.64 ± 0.04 | 0.60 ± 0.074 | 0.57 ± 0.061 | |
| April 2013 | August 2016 | October 2016 | May 2016 | October 2017 | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Overall accuracy | 0.87 | 0.66 | 0.79 | 0.85 | 0.56 | ||||||||||||||||||||
| Kappa index | 0.81 | 0.56 | 0.72 | 0.80 | 0.46 | ||||||||||||||||||||
| Average accuracy | 0.69 | 0.72 | 0.79 | 0.83 | 0.60 | ||||||||||||||||||||
| Class accuracy | C1: 0.64 C2: 0.93 C3: 0.97 C4: 0.88 C5: 0.037 | C1: 0.9 C2: 0.96 C3: 0 C4: 1 C5: 0.76 | C1: 0.72 C2: 0.88 C3: 0.96 C4: 0.39 C5: 0.98 | C1: 0.85 C2: 0.74 C3: 0.98 C4: 0.96 C5: 0.64 | C1: 0.53 C2: 0.78 C3: 0.39 C4: 0.35 C5: 0.92 | ||||||||||||||||||||
| Confusion matrix (in %) | 64 | 0 | 36 | 0 | 0 | 90 | 0 | 0 | 3 | 7 | 72 | 8 | 0 | 0 | 20 | 85 | 2.5 | 10 | 2.5 | 0 | 53 | 0 | 47 | 0 | 0 |
| 2 | 93 | 0 | 0 | 5 | 0 | 96 | 0.4 | 0 | 3.6 | 10 | 88 | 0 | 0 | 1 | 14 | 74 | 11 | 0 | 1 | 6 | 78 | 15 | 0 | 1 | |
| 1 | 0 | 97 | 0 | 2 | 0 | 26 | 0 | 0 | 74 | 0.2 | 3.8 | 96 | 0 | 0 | 0.4 | 0 | 98 | 0 | 1.6 | 60 | 0 | 39 | 0 | 1 | |
| 0 | 0 | 0 | 88 | 12 | 0 | 0 | 0 | 100 | 0 | 0 | 0 | 0 | 39 | 61 | 0.4 | 0 | 2.4 | 96 | 1.2 | 5 | 0 | 0 | 36 | 59 | |
| 0 | 0 | 18 | 78 | 4 | 0 | 22 | 2 | 0 | 76 | 1 | 0 | 0 | 1 | 98 | 0.5 | 7 | 28 | 0.5 | 64 | 0 | 0 | 0 | 8 | 92 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Proença, B.; Frappart, F.; Lubac, B.; Marieu, V.; Ygorra, B.; Bombrun, L.; Michalet, R.; Sottolichio, A. Potential of High-Resolution Pléiades Imagery to Monitor Salt Marsh Evolution After Spartina Invasion. Remote Sens. 2019, 11, 968. https://doi.org/10.3390/rs11080968
Proença B, Frappart F, Lubac B, Marieu V, Ygorra B, Bombrun L, Michalet R, Sottolichio A. Potential of High-Resolution Pléiades Imagery to Monitor Salt Marsh Evolution After Spartina Invasion. Remote Sensing. 2019; 11(8):968. https://doi.org/10.3390/rs11080968
Chicago/Turabian StyleProença, Bárbara, Frédéric Frappart, Bertrand Lubac, Vincent Marieu, Bertrand Ygorra, Lionel Bombrun, Richard Michalet, and Aldo Sottolichio. 2019. "Potential of High-Resolution Pléiades Imagery to Monitor Salt Marsh Evolution After Spartina Invasion" Remote Sensing 11, no. 8: 968. https://doi.org/10.3390/rs11080968
APA StyleProença, B., Frappart, F., Lubac, B., Marieu, V., Ygorra, B., Bombrun, L., Michalet, R., & Sottolichio, A. (2019). Potential of High-Resolution Pléiades Imagery to Monitor Salt Marsh Evolution After Spartina Invasion. Remote Sensing, 11(8), 968. https://doi.org/10.3390/rs11080968