
Using Hyperspectral Imagery to Characterize Rangeland Vegetation Composition at Process-Relevant Scales


Abstract
:1. Introduction
- Develop advanced plant community composition classification methods that could be used to map plant composition at our site and at other NEON sites or in areas with similar hyperspectral data;
- Determine the utility of plant community mapping for detecting the effects of soil type, weather, management, and ecological disturbances (e.g., burrowing mammals) on plant community composition and compositional change over time.
2. Materials and Methods
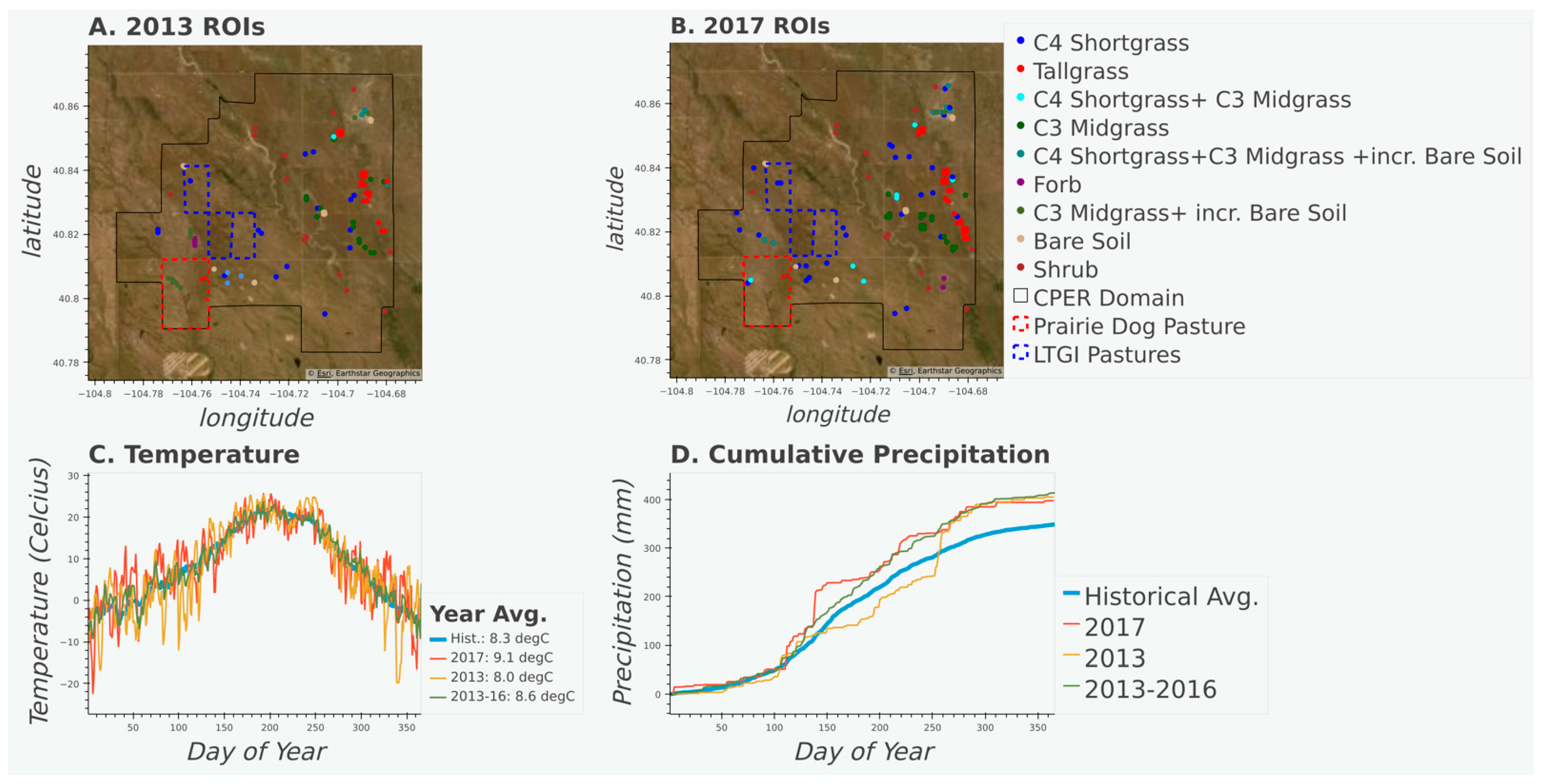
2.1. Study Area
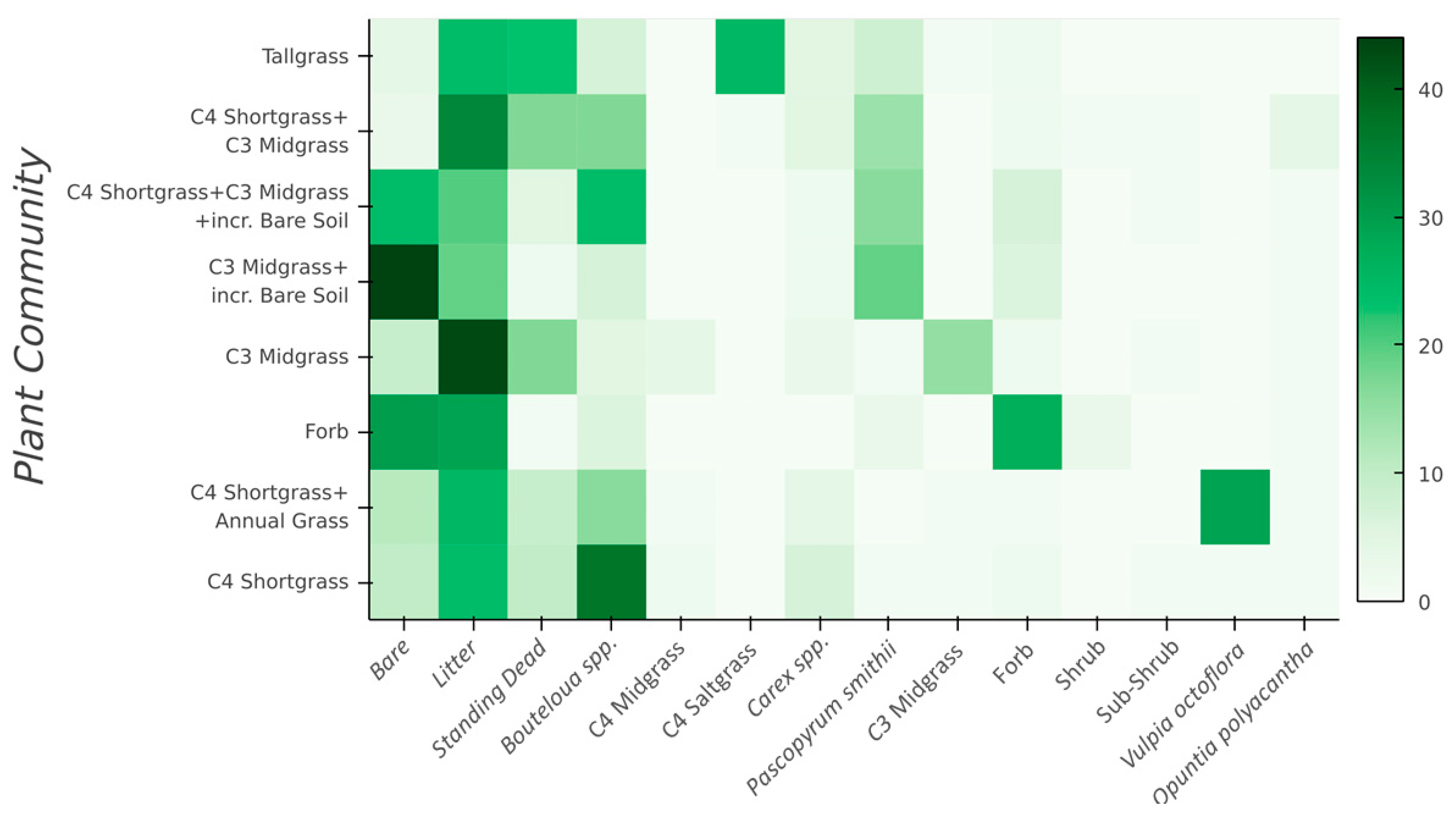
2.2. In situ Vegetation Data
2.3. Hyperspectral Data
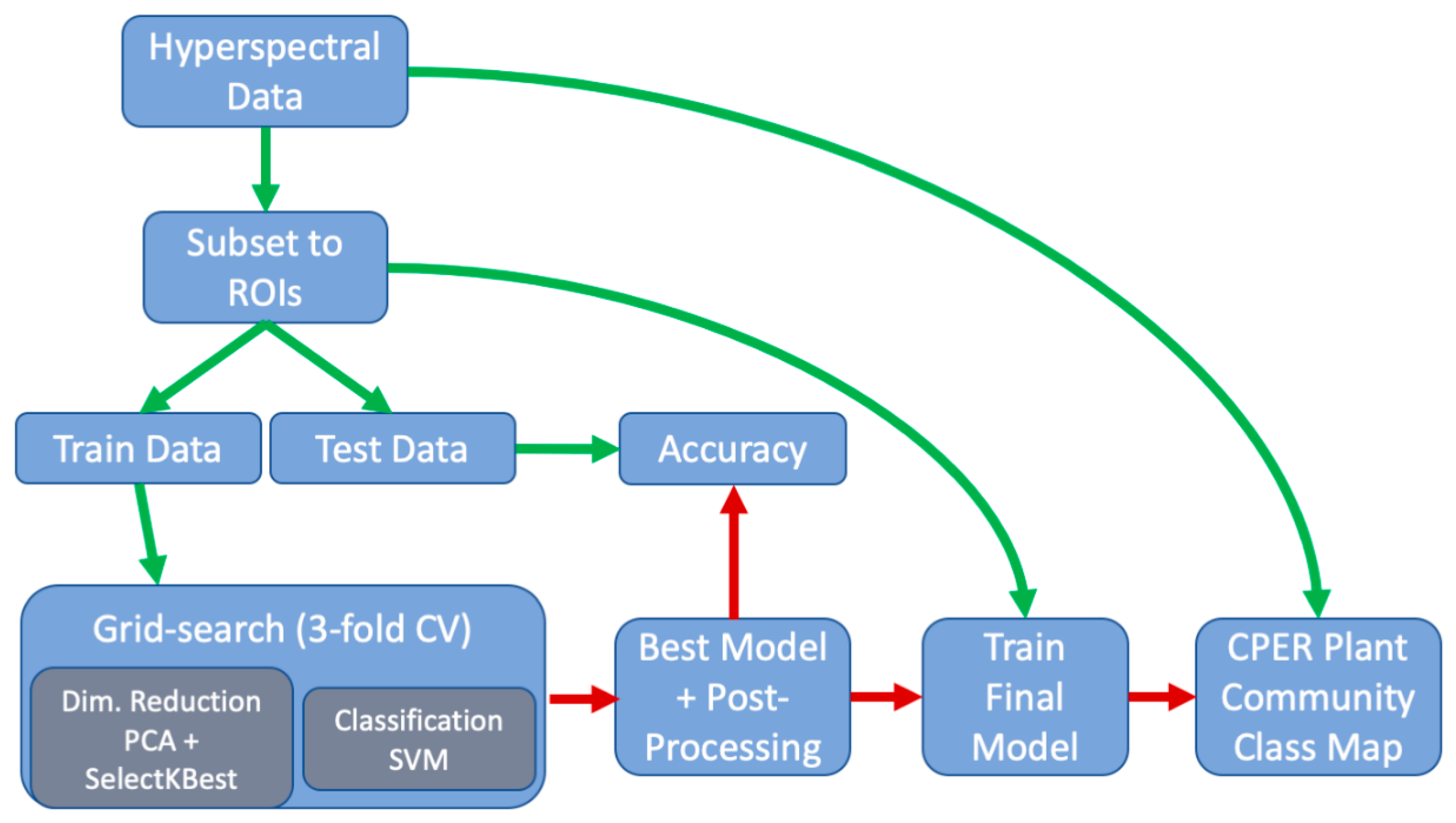
2.4. Vegetation Classification
2.5. Case Studies: Change Detection and Disturbance Impacts
2.6. Computational Environment
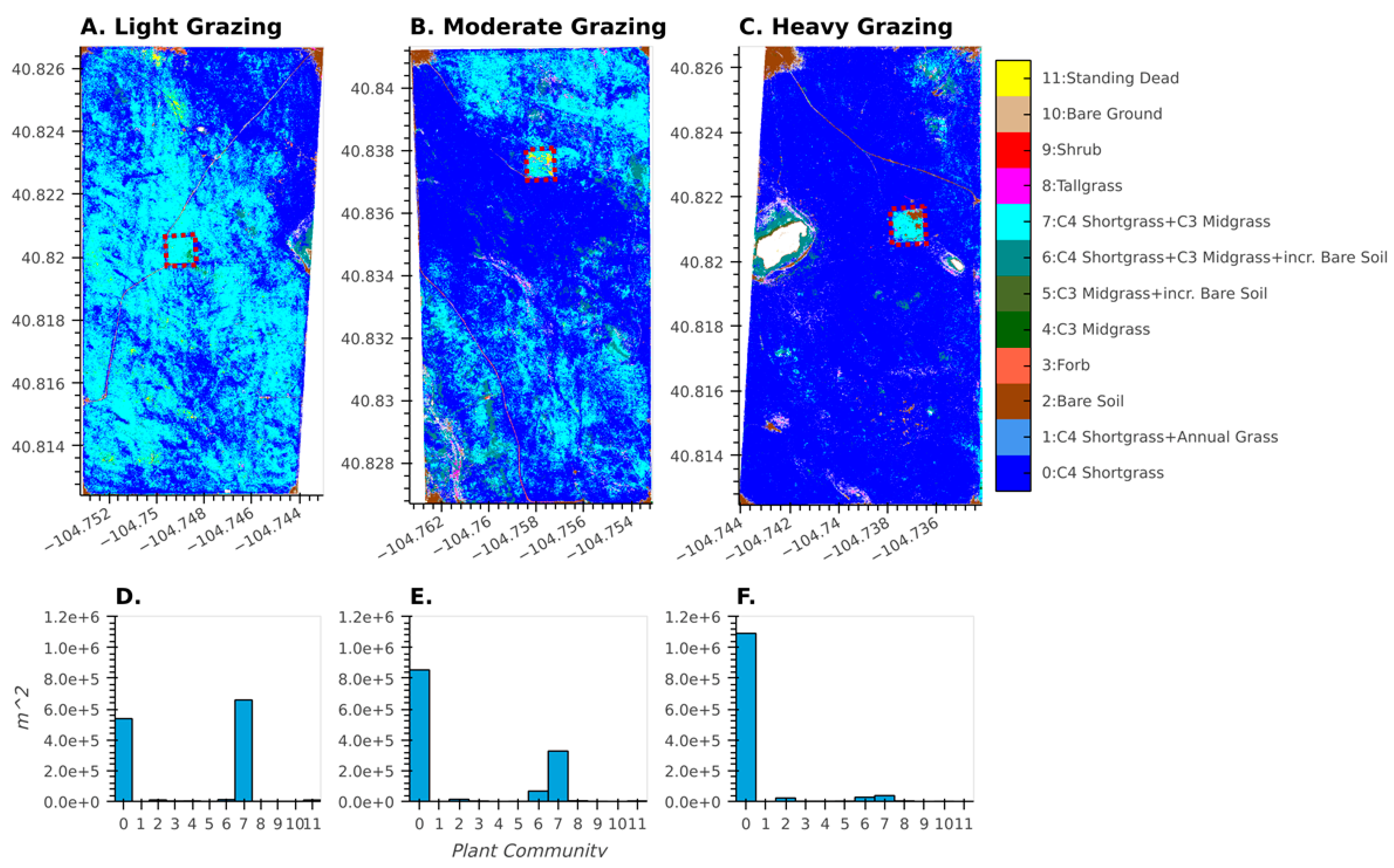
3. Results
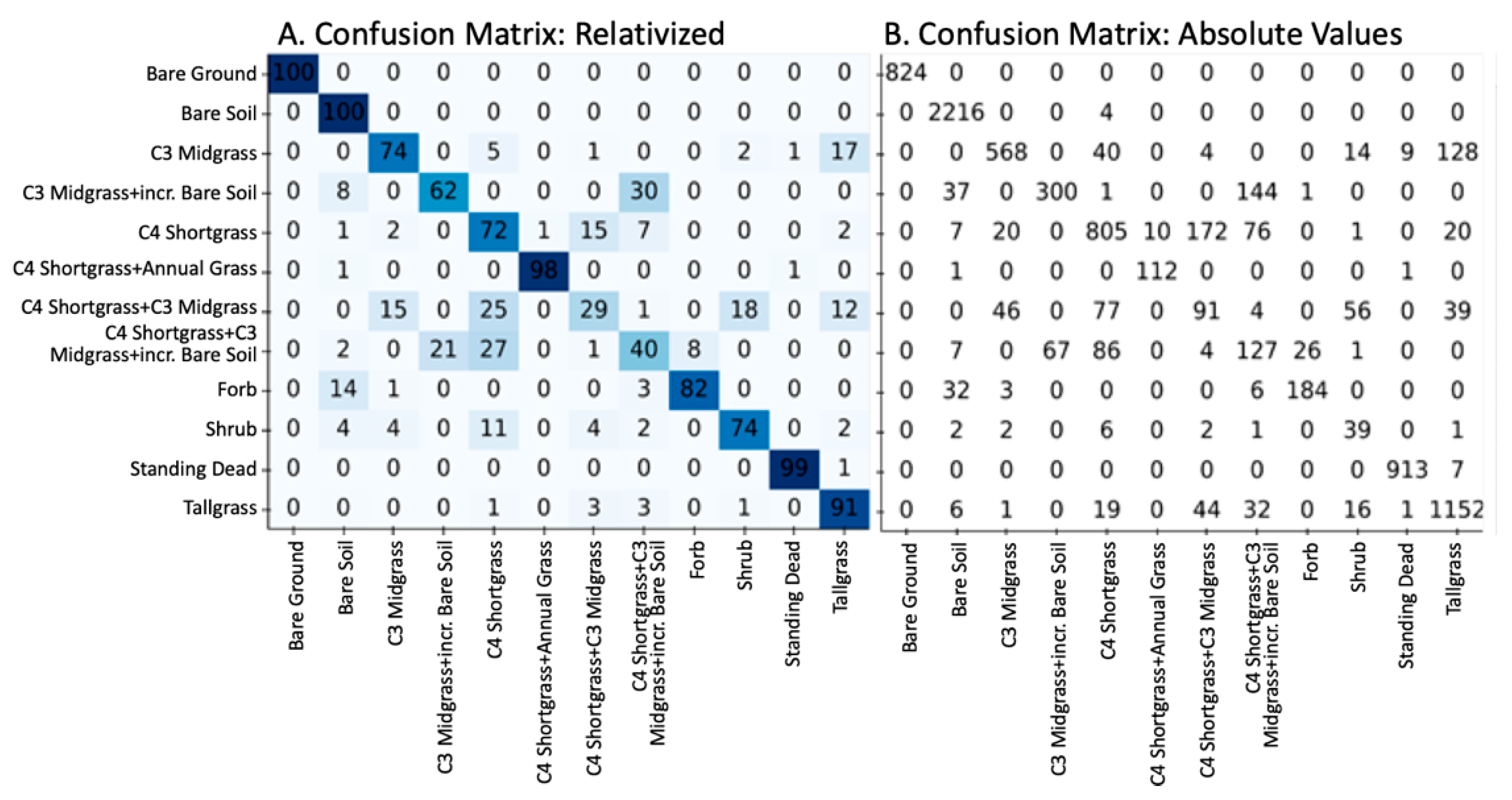
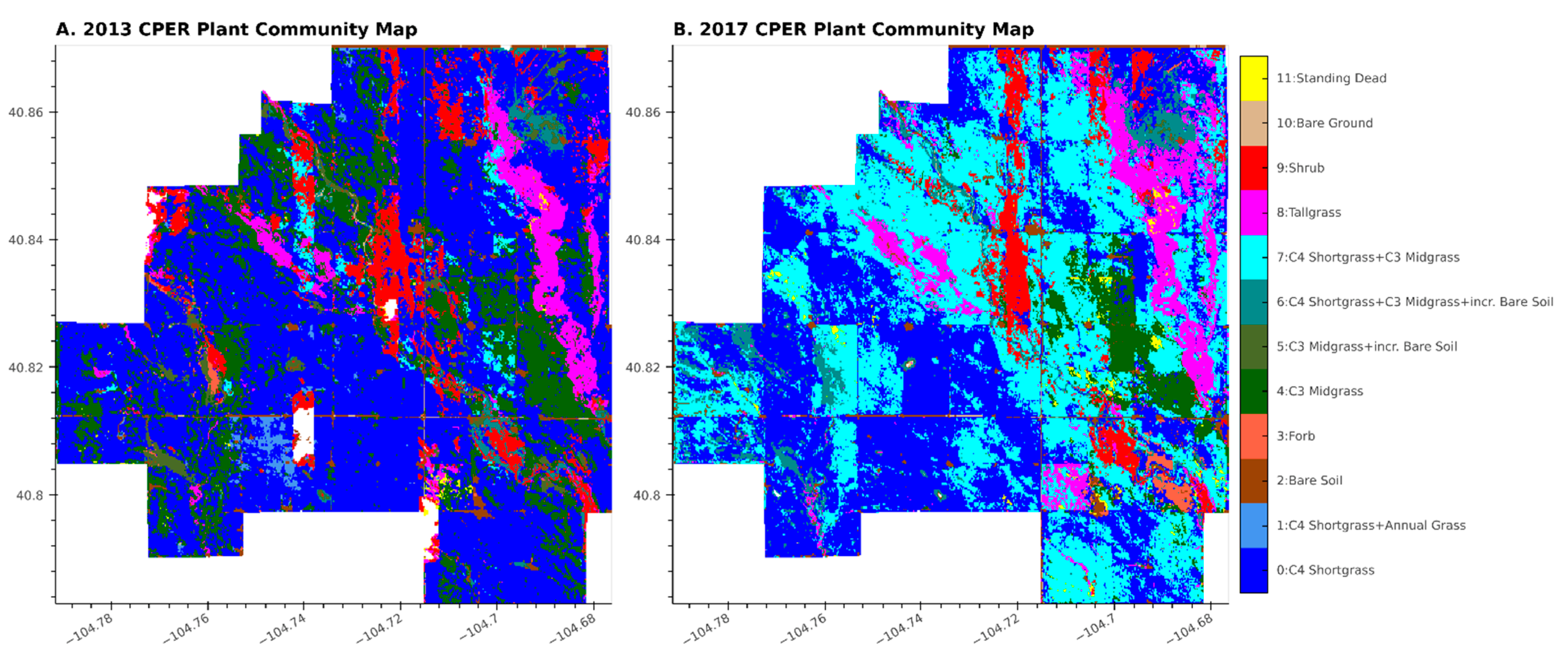
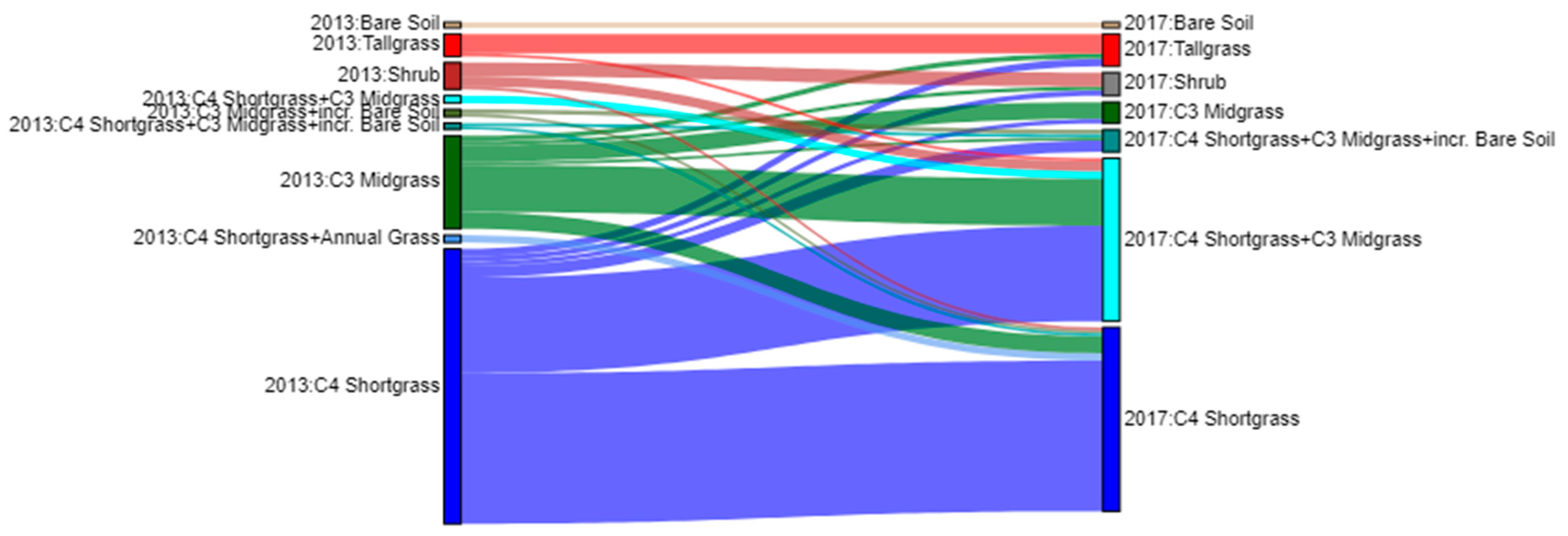
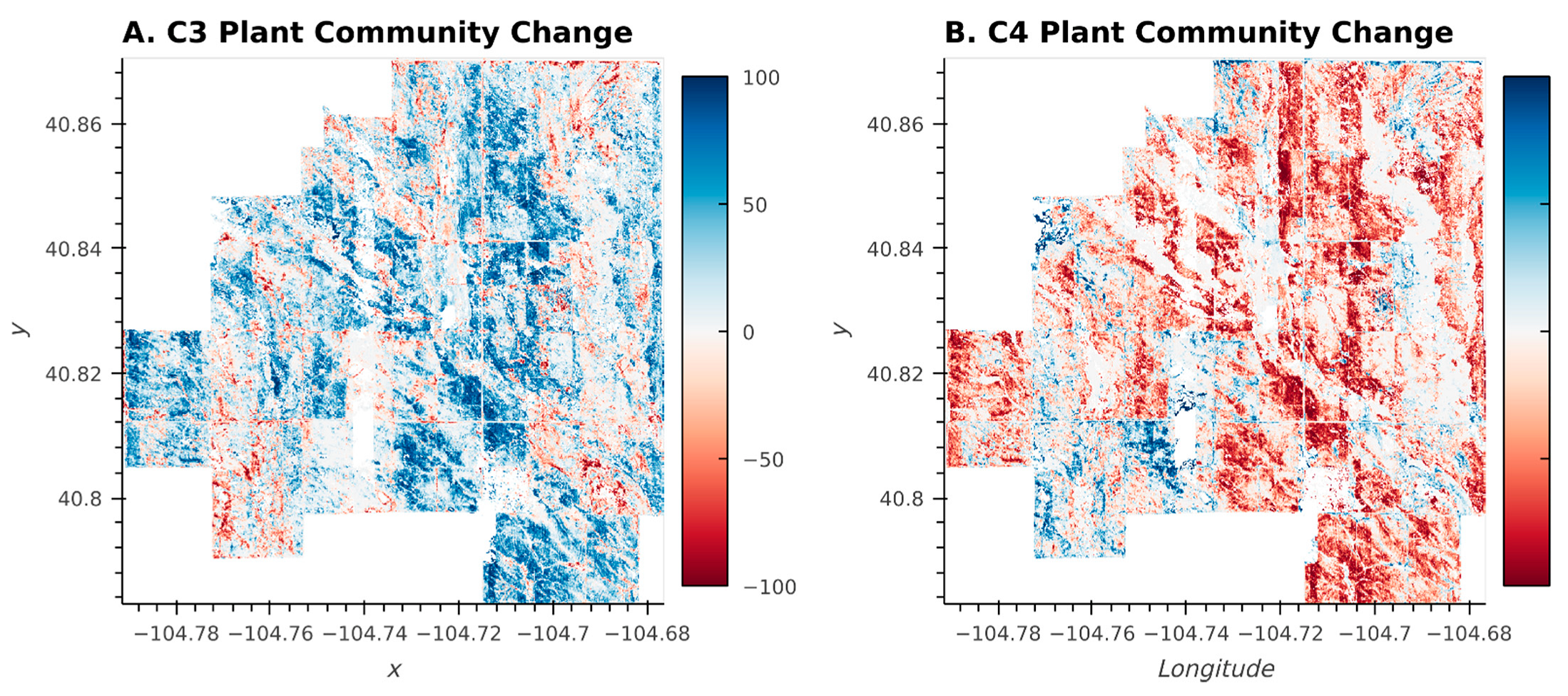
3.1. Accuracy and Plant Community Change
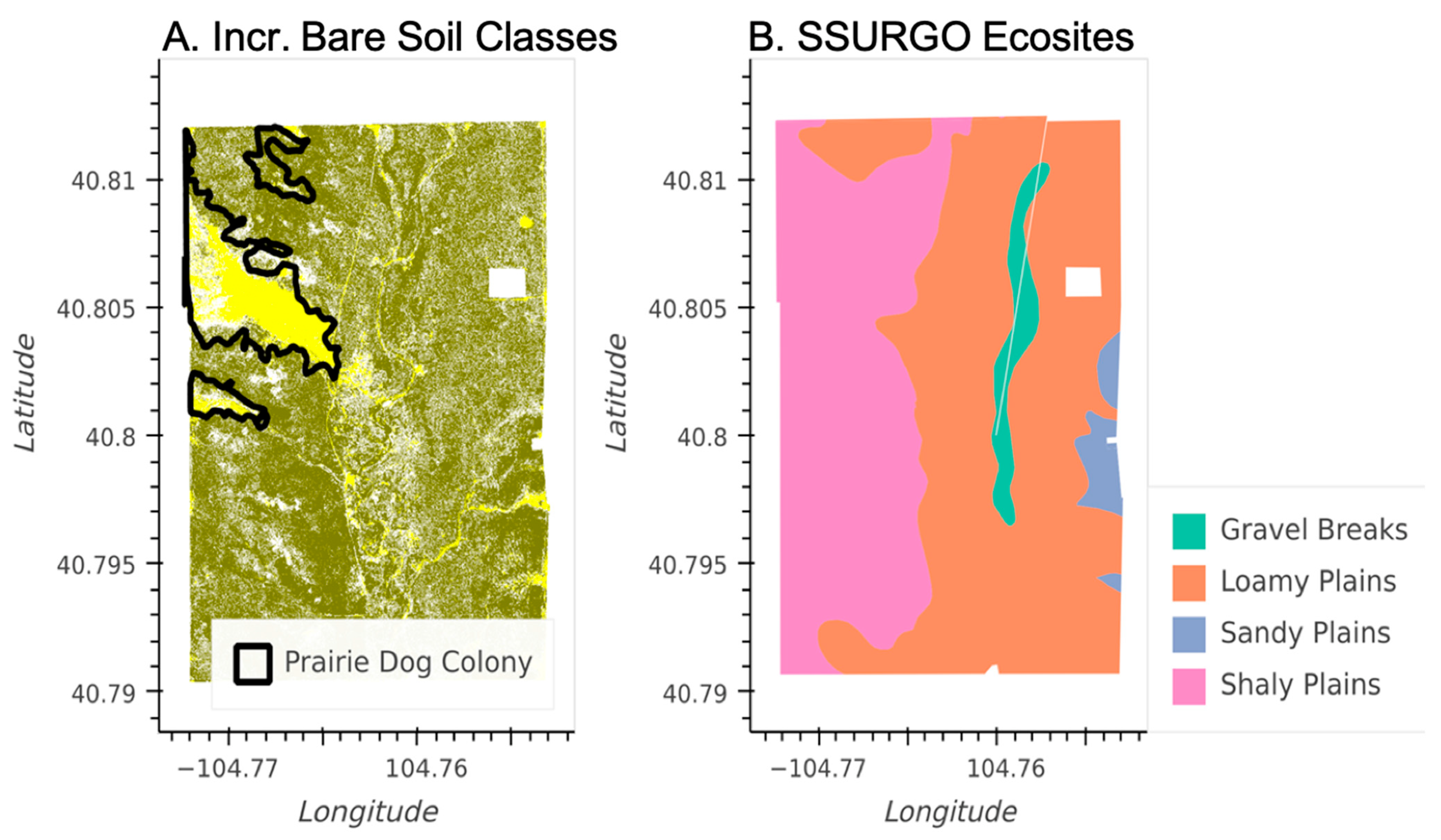
3.2. Characterizing Disturbances
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Irisarri, J.G.N.; Derner, J.D.; Porensky, L.M.; Augustine, D.J.; Reeves, J.L.; Mueller, K.E. Grazing Intensity Differentially Regulates ANPP Response to Precipitation in North American Semiarid Grasslands. Ecol. Appl. 2016, 26, 1370–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augustine, D.J.; McNaughton, S.J. Ungulate Effects on the Functional Species Composition of Plant Communities: Herbivore Selectivity and Plant Tolerance. J. Wildl. Manag. 1998, 62, 1165–1183. [Google Scholar] [CrossRef]
- Raynor, E.J.; Gersie, S.P.; Stephenson, M.B.; Clark, P.E.; Spiegal, S.A.; Boughton, R.K.; Bailey, D.W.; Cibils, A.; Smith, B.W.; Derner, J.D.; et al. Cattle Grazing Distribution Patterns Related to Topography Across Diverse Rangeland Ecosystems of North America. Rangel. Ecol. Manag. 2021, 75, 91–103. [Google Scholar] [CrossRef]
- Davis, S. Nest-Site Selection Patterns and the Influence of Vegetation on Nest Survival of Mixed-Grass Prairie Passerines. The Condor 2005, 107, 605–616. [Google Scholar] [CrossRef]
- Skagen, S.K.; Augustine, D.J.; Derner, J.D. Semi-arid Grassland Bird Responses to Patch-burn Grazing and Drought. J. Wildl. Manag. 2018, 82, 445–456. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.Z.; Giese, M.; Han, X.G.; Wang, D.L.; Zhou, Z.Y.; Brueck, H.; Lin, S.; Taube, F. Land Use and Drought Interactively Affect Interspecific Competition and Species Diversity at the Local Scale in a Semiarid Steppe Ecosystem. Ecol. Res. 2009, 24, 627–635. [Google Scholar] [CrossRef]
- Lauenroth, W. Vegetation of the Shortgrass Steppe. In Ecology of the Shortgrass Steppe: A Long-term Perspective; Lauenroth, W., Burke, I., Eds.; Oxford University Press: New York, NY, USA, 2008; pp. 70–83. [Google Scholar]
- McNaughton, S.J. Ecology of a Grazing Ecosystem: The Serengeti. Ecol. Monogr. 1985, 55, 259–294. [Google Scholar] [CrossRef]
- Milchunas, D.G.; Lauenroth, W.K.; Chapman, P.L.; Kazempour, M.K. Effects of Grazing, Topography, and Precipitation on the Structure of a Semiarid Grassland. Vegetatio 1989, 80, 11–23. [Google Scholar] [CrossRef]
- Peters, D.; Lauenroth, W.; Burke, I. The Role of Disturbances in Shortgrass Steppe Community and Ecosystem Dynamics. In Ecology of the Shortgrass Steppe: A Long-term Perspective; Lauenroth, W., Burke, I., Eds.; Oxford University Press: New York, NY, USA, 2008; pp. 84–118. [Google Scholar]
- Riginos, C.; Porensky, L.M.; Veblen, K.E.; Young, T.P. Herbivory and Drought Generate Short-Term Stochasticity and Long-Term Stability in a Savanna Understory Community. Ecol. Appl. Publ. Ecol. Soc. Am. 2018, 28, 323–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.; Homer, C.; Yang, L.; Danielson, P.; Dewitz, J.; Li, C.; Zhu, Z.; Xian, G.; Howard, D. Overall Methodology Design for the United States National Land Cover Database 2016 Products. Remote Sens. 2019, 11, 2971. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.O.; Allred, B.W.; Naugle, D.E.; Maestas, J.D.; Donnelly, P.; Metz, L.J.; Karl, J.; Smith, R.; Bestelmeyer, B.; Boyd, C.; et al. Innovation in Rangeland Monitoring: Annual, 30 m, Plant Functional Type Percent Cover Maps for U.S. Rangelands, 1984–2017. Ecosphere 2018, 9, e02430. [Google Scholar] [CrossRef]
- Lopatin, J.; Fassnacht, F.E.; Kattenborn, T.; Schmidtlein, S. Mapping Plant Species in Mixed Grassland Communities Using Close Range Imaging Spectroscopy. Remote Sens. Environ. 2017, 201, 12–23. [Google Scholar] [CrossRef]
- Porensky, L.; Derner, J.; Pellatz, D. Plant Community Responses to Historical Wildfire in a Shrubland-Grassland Ecotone Reveal Hybrid Disturbance Response. Ecosphere 2018, 8, e02363. [Google Scholar] [CrossRef] [Green Version]
- Duniway, M.; Bestelmeyer, B.; Tugel, A. Soil Processes and Properties That Distinguish Ecological Sites and States. Rangelands 2010, 32, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Hoover, D.L.; Lauenroth, W.K.; Milchunas, D.G.; Porensky, L.M.; Augustine, D.J.; Derner, J.D. Sensitivity of Productivity to Precipitation Amount and Pattern Varies by Topographic Position in a Semiarid Grassland. Ecosphere 2021, 12, e03376. [Google Scholar] [CrossRef]
- Theron, E.; van Aardt, A.; du Preez, P. Vegetation Distribution along a Granite Catena, Southern Kruger National Park, South Africa. Koedoe 2020, 62, 11. [Google Scholar] [CrossRef]
- Jones, M.O.; Robinson, N.P.; Naugle, D.E.; Maestas, J.D.; Reeves, M.C.; Lankston, R.W.; Allred, B.W. Annual and 16-Day Rangeland Production Estimates for the Western United States. Rangel. Ecol. Manag. 2021, 77, 112–117. [Google Scholar] [CrossRef]
- Möckel, T.; Löfgren, O.; Prentice, H.C.; Eklundh, L.; Hall, K. Airborne Hyperspectral Data Predict Ellenberg Indicator Values for Nutrient and Moisture Availability in Dry Grazed Grasslands within a Local Agricultural Landscape. Ecol. Indic. 2016, 66, 503–516. [Google Scholar] [CrossRef]
- Pullanagari, R.R.; Kereszturi, G.; Yule, I. Integrating Airborne Hyperspectral, Topographic, and Soil Data for Estimating Pasture Quality Using Recursive Feature Elimination with Random Forest Regression. Remote Sens. 2018, 10, 1117. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Townsend, P.A.; Schweiger, A.K.; Couture, J.J.; Singh, A.; Hobbie, S.E.; Cavender-Bares, J. Mapping Foliar Functional Traits and Their Uncertainties across Three Years in a Grassland Experiment. Remote Sens. Environ. 2019, 221, 405–416. [Google Scholar] [CrossRef]
- Marcinkowska-Ochtyra, A.; Zagajewski, B.; Raczko, E.; Ochtyra, A.; Jarocińska, A. Classification of High-Mountain Vegetation Communities within a Diverse Giant Mountains Ecosystem Using Airborne APEX Hyperspectral Imagery. Remote Sens. 2018, 10, 570. [Google Scholar] [CrossRef] [Green Version]
- Rapinel, S.; Rossignol, N.; Hubert-Moy, L.; Bouzillé, J.-B.; Bonis, A. Mapping Grassland Plant Communities Using a Fuzzy Approach to Address Floristic and Spectral Uncertainty. Appl. Veg. Sci. 2018, 21, 678–693. [Google Scholar] [CrossRef]
- Kopeć, D.; Sabat-Tomala, A.; Michalska-Hejduk, D.; Jarocińska, A.; Niedzielko, J. Application of Airborne Hyperspectral Data for Mapping of Invasive Alien Spiraea Tomentosa, L.: A Serious Threat to Peat Bog Plant Communities. Wetl. Ecol. Manag. 2020, 28, 357–373. [Google Scholar] [CrossRef] [Green Version]
- Bradley, B.; Curtis, C.; Fusco, E.; Abatzoglou, J.; Balch, J.; Dadashi, S.; Tuanmu, M.-N. Cheatgrass (Bromus Tectorum) Distribution in the Intermountain Western United States and Its Relationship to Fire Frequency, Seasonality, and Ignitions. Biol. Invasions 2018, 20, 1493–1506. [Google Scholar] [CrossRef] [Green Version]
- Augustine, D.J.; Derner, J.D.; Fernández-Giménez, M.E.; Porensky, L.M.; Wilmer, H.; Briske, D.D. Adaptive, Multipaddock Rotational Grazing Management: A Ranch-Scale Assessment of Effects on Vegetation and Livestock Performance in Semiarid Rangeland. Rangel. Ecol. Manag. 2020, 73, 796–810. [Google Scholar] [CrossRef]
- Herrick, J.; Zee, J.V.; Havstad, K.; Burkett, L.M.; Whitford, W. Monitoring Manual for Grassland, Shrubland and Savanna Ecosystems. Methods and Interpretation; Jornada Experimental Range: Las Cruces, NM, USA, 2005; Volume 1. [Google Scholar]
- Karpowicz, B.; Kampe, T. NEON Imaging Spectrometer Radiance to Reflectance Algorithm Theoretical Basis Document; The National Ecological Observatory Network: Boulder, CO, USA, 2015. [Google Scholar]
- Liu, N.; Chlus, A.; Townsend, P.A. HyToolsPro: An Open Source Package for Pre-Processing Airborne Hyperspectral Images. In Proceedings of the AGU Fall Meeting 2019, San Francisco, CA, USA, 9–13 December 2019. [Google Scholar]
- Daughtry, C.S.T.; Doraiswamy, P.C.; Hunt, E.R.; Stern, A.J.; McMurtrey, J.E.; Prueger, J.H. Remote Sensing of Crop Residue Cover and Soil Tillage Intensity. Soil Tillage Res. 2006, 91, 101–108. [Google Scholar] [CrossRef]
- Guerschman, J.P.; Hill, M.J.; Renzullo, L.J.; Barrett, D.J.; Marks, A.S.; Botha, E.J. Estimating Fractional Cover of Photosynthetic Vegetation, Non-Photosynthetic Vegetation and Bare Soil in the Australian Tropical Savanna Region Upscaling the EO-1 Hyperion and MODIS Sensors. Remote Sens. Environ. 2009, 113, 928–945. [Google Scholar] [CrossRef]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-Learn: Machine Learning in Python. Mach. Learn. Python 2011, 12, 2825–2830. [Google Scholar]
- Klipple, G.E.; Costello, D.F. Vegetation and Cattle Responses to Different Intensities of Grazing on Short-Grass Ranges on Tlie Central Great Plains; US Department of Agriculture: Washington, DC, USA, 1960. [Google Scholar]
- Porensky, L.M.; Derner, J.D.; Augustine, D.J.; Milchunas, D.G. Plant Community Composition After 75 Yr of Sustained Grazing Intensity Treatments in Shortgrass Steppe. Rangel. Ecol. Manag. 2017, 70, 456–464. [Google Scholar] [CrossRef]
- Augustine, D.J.; Derner, J.D.; Detling, J.K. Testing for Thresholds in a Semiarid Grassland: The Influence of Prairie Dogs and Plague. Rangel. Ecol. Manag. 2014, 67, 701–709. [Google Scholar] [CrossRef]
- Hart, R.H.; Mary, M. Ashby Grazing Intensities, Vegetation, and Heifer Gains: 55 Years on Shortgrass. J. Range Manag. 1998, 51, 392–398. [Google Scholar] [CrossRef] [Green Version]
- Merkel, D. Docker: Lightweight Linux Containers for Consistent Development and Deployment. Linux J. 2014, 239, 2. [Google Scholar]
- Kluyver, T.; Ragan-Kelley, B.; Pérez, F.; Bussonnier, M.; Frederic, J.; Hamrick, J.; Grout, J.; Corlay, S.; Ivanov, P.; Abdalla, S.; et al. Jupyter Notebooks—A Publishing Format for Reproducible Computational Workflows. In Proceedings of the 20th International Conference on Electronic Publishing, Toronto, ON, Canada, 1 January 2016. [Google Scholar]
- Gaffney, R. Rmg55/Container_stacks: Dsir_2021.04.1; CERN: Meyrin, Switzerland, 2021. [Google Scholar]
- Dask Development Team. Dask: Library for Dynamic Task Scheduling; VMware Inc.: Palo Alto, CA, USA, 2016. [Google Scholar]
- Rocklin, M. Dask: Parallel Computation with Blocked Algorithms and Task Scheduling. In Proceedings of the 14th Python in Science Conference, Austin, TX, USA, 6–12 July 2015; pp. 126–132. [Google Scholar]
- Hoyer, S.; Hamman, J.J. Xarray: N-D Labeled Arrays and Datasets in Python. J. Open Res. Softw. 2017, 5, 10. [Google Scholar] [CrossRef] [Green Version]
- Bednar, J.A.; Crail, J.; Crist-Harif, J.; Rudiger, P.; Brener, G.; B, C.; Mease, J.; Signell, J.; Stevens, J.-L.; Collins, B.; et al. Holoviz/Datashader: Version 0.12.1; Zenodo: Meyrin, Switzerland, 2021. [Google Scholar]
- Rudiger, P.; Signell, J.; Bednar, J.A.; Andrew; Stevens, J.-L.; B, C.; Kbowen; Fast, T.; Graser, A.; AurelienSciarra; et al. Holoviz/Hvplot: Version 0.7.1; Zenodo: Meyrin, Switzerland, 2021. [Google Scholar]
- Rudiger, P.; Stevens, J.-L.; Bednar, J.A.; Nijholt, B.; Mease, J.; Andrew; B, C.; Randelhoff, A.; Tenner, V.; Maxalbert; et al. Holoviz/Holoviews: Version 1.14.3; Zenodo: Meyrin, Switzerland, 2021. [Google Scholar]
- Augustine, D.J.; Derner, J.D.; Milchunas, D.; Blumenthal, D.; Porensky, L.M. Grazing Moderates Increases in C3 Grass Abundance over Seven Decades across a Soil Texture Gradient in Shortgrass Steppe. J. Veg. Sci. 2017, 28, 562–572. [Google Scholar] [CrossRef]
- Gaffney, R.; Porensky, L.; Gao, F.; Irisarri, J.; Durante, M.; Derner, J.; Augustine, D. Using APAR to Predict Aboveground Plant Productivity in Semi-Aid Rangelands: Spatial and Temporal Relationships Differ. Remote Sens. 2018, 10, 1474. [Google Scholar] [CrossRef] [Green Version]
- Milchunas, D.G.; Mosier, A.R.; Morgan, J.A.; LeCain, D.R.; King, J.Y.; Nelson, J.A. Elevated CO2 and Defoliation Effects on a Shortgrass Steppe: Forage Quality versus Quantity for Ruminants. Agric. Ecosyst. Environ. 2005, 111, 166–184. [Google Scholar] [CrossRef]
- Augustine, D.J.; Derner, J.D. Long-Term Effects of Black-Tailed Prairie Dogs on Livestock Grazing Distribution and Mass Gain. J. Wildl. Manag. 2021, 85, 1332–1343. [Google Scholar] [CrossRef]
- Gersie, S.P.; Augustine, D.J.; Derner, J.D. Cattle Grazing Distribution in Shortgrass Steppe: Influences of Topography and Saline Soils. Rangel. Ecol. Manag. 2019, 72, 602–614. [Google Scholar] [CrossRef]
- Senft, R.L.; Rittenhouse, L.R.; Woodmansee, R.G. Factors Influencing Patterns of Cattle Grazing Behavior on Shortgrass Steppe. J. Range Manag. 1985, 38, 82. [Google Scholar] [CrossRef]
- Swain, D.L.; Charters, S.M. Back to Nature With Fenceless Farms—Technology Opportunities to Reconnect People and Food. Front. Sustain. Food Syst. 2021, 5, 222. [Google Scholar] [CrossRef]
- Magiera, A.; Feilhauer, H.; Waldhardt, R.; Wiesmair, M.; Otte, A. Modelling Biomass of Mountainous Grasslands by Including a Species Composition Map. Ecol. Indic. 2017, 78, 8–18. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Group | Species Included, or Cover Characteristics |
|---|---|
| C4 Shortgrasses | Bouteloua gracilis, Bouteloua dactyloides |
| C4 Saltgrasses | Distichlis dactyloides, Sporobolus airoides |
| C4 Midgrasses | Aristida purpurea, Sporobolus cryptandrus |
| C3 Sedges | Carex duriscula, Carex filifolia |
| C3 Western wheatgrass | Pascopyrum smithii |
| C3 Needle and thread | Hesperostipa comata |
| C3 Squirreltail | Elymus elymoides |
| C3 Annual grass | Vulpia octoflora |
| Forbs | >20 species, including annuals and perennials |
| Subshrubs | Artemisia frigida, Eriogonum effusum, Gutierrezia sarothrae |
| Shrubs | Atriplex canescens, Ericameria nauseosa |
| Standing dead | Residual standing vegetation produced in prior year |
| Bare soil | Exposed mineral soil |
| Litter | Dead vegetation, dung, or lichen covering the soil surface |
| Model Component | Parameter | Search Grid |
|---|---|---|
| SVM hyperparameter | C (Regularization) | (1, 10, 1 × 102, 5 × 102, 1 × 104, 5 × 104, 1 × 105) |
| SVM hyperparameter | γ (Kernel Coef.) | (1 × 10−6, 1 × 10−5, 5 × 10−5, 1 × 10−4, 5 × 10−3, 1 × 10−3, 0.01, 0.1) |
| PCA | N Components | (60, 80, 100, 120, 140, 160) |
| SelectKBest | N Components | (1, 5, 9, 13, 17) |
| Plant Community Class | Precision | Recall | F1-Score |
|---|---|---|---|
| Bare Ground | 1.0 | 1.0 | 1.0 |
| Bare Soil | 0.96 | 1.0 | 0.98 |
| C3 Midgrass | 0.89 | 0.74 | 0.81 |
| C3 Midgrass-incr Bare Soil | 0.82 | 0.62 | 0.71 |
| C4 Shortgrass | 0.78 | 0.72 | 0.75 |
| C4 Shortgrass + Annual Grass | 0.92 | 0.98 | 0.95 |
| C4 Shortgrass + C3 Midgrass | 0.29 | 0.29 | 0.29 |
| C4 Shortgrass + C3 Midgrass+incr. Bare Soil | 0.33 | 0.40 | 0.36 |
| Forb | 0.87 | 0.82 | 0.84 |
| Shrub | 0.31 | 0.74 | 0.43 |
| Standing Dead | 0.99 | 0.99 | 0.99 |
| Tallgrass | 0.86 | 0.91 | 0.88 |
| Accuracy | 0.85 | 0.85 | 0.85 |
| Macro Average | 0.75 | 0.77 | 0.75 |
| Weighted Average | 0.86 | 0.85 | 0.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaffney, R.; Augustine, D.J.; Kearney, S.P.; Porensky, L.M. Using Hyperspectral Imagery to Characterize Rangeland Vegetation Composition at Process-Relevant Scales. Remote Sens. 2021, 13, 4603. https://doi.org/10.3390/rs13224603
Gaffney R, Augustine DJ, Kearney SP, Porensky LM. Using Hyperspectral Imagery to Characterize Rangeland Vegetation Composition at Process-Relevant Scales. Remote Sensing. 2021; 13(22):4603. https://doi.org/10.3390/rs13224603
Chicago/Turabian StyleGaffney, Rowan, David J. Augustine, Sean P. Kearney, and Lauren M. Porensky. 2021. "Using Hyperspectral Imagery to Characterize Rangeland Vegetation Composition at Process-Relevant Scales" Remote Sensing 13, no. 22: 4603. https://doi.org/10.3390/rs13224603
APA StyleGaffney, R., Augustine, D. J., Kearney, S. P., & Porensky, L. M. (2021). Using Hyperspectral Imagery to Characterize Rangeland Vegetation Composition at Process-Relevant Scales. Remote Sensing, 13(22), 4603. https://doi.org/10.3390/rs13224603