
Assessing Vegetation Response to Multi-Scalar Drought across the Mojave, Sonoran, Chihuahuan Deserts and Apache Highlands in the Southwest United States

 , ,
, ,  ,
,  and
and
Abstract
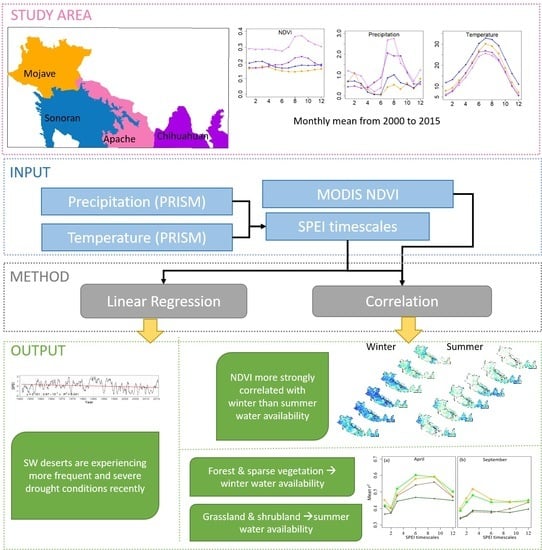
:
1. Introduction
- How do the trends in water availability (SPEI) at different timescales vary by ecoregion?
- How does vegetation productivity (NDVI) respond to changes in water availability (SPEI)?
- For each ecoregion, what was the dominant timescale for the relationship?
- How uniform was the response of NDVI to SPEI across these ecoregions?
- Does this relationship change by dominant vegetation type?
- What do these results suggest about plant physiology responses to drought stress in these ecoregions?
2. Materials and Methods
2.1. Study Area
2.2. Data
2.2.1. Vegetation Productivity (MODIS NDVI)
2.2.2. Climate Data (Temperature and Precipitation)
2.2.3. Drought Index (SPEI)
2.2.4. Landcover Data
2.3. Methods and Analysis
2.3.1. Long Term Trends and Variation of Different SPEI Timescales by Ecoregions
2.3.2. Correlation Analysis for the Relationship of NDVI to SPEI
3. Results
3.1. Long Term Trend and Variation of Different SPEI Timescales by Ecoregions
3.2. Seasonal NDVI Response to Different SPEI Timescales
3.2.1. April NDVI Response to Different SPEI Timescales
3.2.2. September NDVI Response to SPEI Timescales
3.3. Vegetation Types Response by Ecoregions
4. Discussion
4.1. Trend and Interannual Variations of Different SPEI Timescales
4.2. Ecoregion Response to Different SPEI Timescales
4.3. Vegetation Type Responses to SPEI Timescales
5. Conclusions
- There was a weak downward interannual trend of SPEI from 1950 to 2015, and the frequency and severity of dry periods are increasing in the 21st century.
- Vegetation productivity depends on seasonal water availability and drought conditions. The impact of water stress (SPEI) was greater on vegetation productivity during the winter than during the summer.
- The vegetation productivity response to SPEI timescales also depends on the vegetation types and was different between winter and summer. Grassland and shrubland productivity were more dependent on summer water availability whereas forest and sparse vegetation productivity was more dependent on winter water availability.
- We identified the dominant drought timescale that has the strongest influence on vegetation productivity on each ecoregion and vegetation types. This information can be useful for land managers to understand and mitigate drought impacts and help in enhancing our understanding of vegetation vulnerability to climate change.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A




References
- Archer, S.R.; Predick, K.I. Climate change and ecosystems of the Southwestern United States. Rangelands 2008, 30, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Gremer, J.R.; Bradford, J.B.; Munson, S.M.; Duniway, M.C. Desert grassland responses to climate and soil moisture suggest divergent vulnerabilities across the southwestern United States. Glob. Chang. Biol. 2015, 21, 4049–4062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garfin, G.; Jardine, A.; Merideth, R.; Black, M.; LeRoy, S. Assessment of Climate Change in the Southwest United States; Island Press: Washington, DC, USA, 2013; 531p. [Google Scholar] [CrossRef]
- Zhang, X.; Goldberg, M.; Tarpley, D.; Friedl, M.A.; Morisette, J.; Kogan, F.; Yu, Y. Drought-induced vegetation stress in southwestern North America. Environ. Res. Lett. Environ. Res. Lett. 2010, 5, 24008–24011. [Google Scholar] [CrossRef]
- Munson, S.M.; Muldavin, E.H.; Belnap, J.; Peters, D.P.C.; Anderson, J.P.; Reiser, M.H.; Gallo, K.; Melgoza-Castillo, A.; Herrick, J.E.; Christiansen, T.A. Regional signatures of plant response to drought and elevated temperature across a desert ecosystem. Ecology 2013, 94, 2030–2041. [Google Scholar] [CrossRef] [Green Version]
- Cañón, J.; Domínguez, F.; Valdes, J.B. Vegetation responses to precipitation and temperature: A spatiotemporal analysis of ecoregions in the Colorado River Basin. Int. J. Remote Sens. 2011, 1161. [Google Scholar] [CrossRef]
- Barnes, M.L.; Moran, M.S.; Scott, R.L.; Kolb, T.E.; Ponce-campos, G.E.; Moore, D.J.P.; Ross, M.A.; Mitra, B.; Dore, S. Vegetation productivity responds to sub—Annual climate conditions across semiarid biomes. Ecosphere 2016, 7, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Ichii, K.; Kawabata, A.; Yamaguchi, Y. Global correlation analysis for NDVI and climatic variables and NDVI trends: 1982–1990. Int. J. Remote Sens. 2002, 23, 3873–3878. [Google Scholar] [CrossRef]
- Kawabata, A.; Ichii, K.; Yamaguchi, Y. Global monitoring of interannual changes in vegetation activities using NDVI and its relationships to temperature and precipitation. Int. J. Remote Sens. 2001, 22, 1377–1382. [Google Scholar] [CrossRef]
- Wang, J.; Rich, P.M.; Price, K.P. Temporal responses of NDVI to precipitation and temperature in the central Great Plains, USA. Int. J. Remote Sens. 2003, 24, 2345–2364. [Google Scholar] [CrossRef]
- Vicente-serrano, S.M.; Gouveia, C.; Julio, J.; Beguería, S.; Trigo, R. Response of vegetation to drought time-scales across global land biomes. Proc. Natl. Acad. Sci. USA 2012, 110, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKee, T.B.; Doesken, N.J.; Kleist, J. The relationship of drought frequency and duration to time scales. In Proceedings of the 8th Conference of Applied Climatology, Anaheim, CA, USA, 17–22 January 1993; pp. 17–22. [Google Scholar]
- Ji, L.; Peters, A.J. Assessing vegetation response to drought in the northern Great Plains using vegetation and drought indices. Remote Sens. Environ. 2003, 87, 85–98. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A multiscalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef] [Green Version]
- McClaran, M.P.; Wei, H. Recent drought phase in a 73-year record at two spatial scales: Implications for livestock production on rangelands in the Southwestern United States. Agric. For. Meteorol. 2014, 197, 40–51. [Google Scholar] [CrossRef]
- Rouse, J.W. Monitoring the Vernal Advancement and Retrogradation of Natural Vegetation. NASA/GSFCT 1973. Available online: https://scholar.google.com/scholar?hl=en&as_sdt=0%2C48&q=Monitoring+the+vernal+advancement+and+retrogradation+of+natural+vegetation.+&btnG= (accessed on 23 February 2021).
- Tucker, C.J. Red and Photographic Infrared, linear Combinations for Monitoring Vegetation. Remote Sens. Environ. 1979, 150, 127–150. [Google Scholar] [CrossRef] [Green Version]
- Geng, L.; Ma, M.; Yu, W.; Wang, X.; Jia, S. Validation of the MODIS NDVI Products in Different Land-Use Types Using In Situ Measurements in the Heihe River Basin. IEEE Geosci. Remote Sens. Lett. 2014, 11, 1649–1653. [Google Scholar] [CrossRef]
- Anees, A.; Aryal, J. A Statistical Framework for Near-Real Time Detection of Beetle Infestation in Pine Forests Using MODIS Data. IEEE Geosci. Remote Sens. Lett. 2014, 11, 1717–1721. [Google Scholar] [CrossRef]
- Anees, A.; Aryal, J. Near-Real Time Detection of Beetle Infestation in Pine Forests Using MODIS Data. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2014, 7, 3713–3723. [Google Scholar] [CrossRef]
- Ardakani, A.S.; Zoej, M.J.V.; Mohammadzadeh, A.; Mansourian, A. Spatial and Temporal Analysis of Fires Detected by MODIS Data in Northern Iran from 2001 to 2008. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2011, 4, 216–225. [Google Scholar] [CrossRef]
- Shahabfar, A.; Ghulam, A.; Conrad, C. Understanding Hydrological Repartitioning and Shifts in Drought Regimes in Central and South-West Asia Using MODIS Derived Perpendicular Drought Index and TRMM Data. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2014, 7, 983–993. [Google Scholar] [CrossRef]
- Gouveia, C.M.; Trigo, R.M.; Beguería, S.; Vicente-Serrano, S.M. Drought impacts on vegetation activity in the Mediterranean region: An assessment using remote sensing data and multi-scale drought indicators. Glob. Planet. Chang. 2017, 151, 15–27. [Google Scholar] [CrossRef] [Green Version]
- The Nature Conservancy. TNC Terrestrial Ecoregions; The Nature Conservancy: Arlington, VA, USA, 2012. [Google Scholar]
- Shepard, C.; Schaap, M.G.; Crimmins, M.A.; Van Leeuwen, W.J.D.; Rasmussen, C. Geoderma Regional Subsurface soil textural control of aboveground productivity in the US Desert Southwest. GEODRS 2015, 4, 44–54. [Google Scholar] [CrossRef]
- Reynolds, J.F.; Kemp, P.R.; Ogle, K.; Fernández, R.J. Modifying the “pulse-reserve” paradigm for deserts of North America: Precipitation pulses, soil water, and plant responses. Oecologia 2004, 141, 194–210. [Google Scholar] [CrossRef]
- Lee, J.; Dimmitt, M.; Lee, C.; Arroyo, R. Deserts of North America. 2016. Available online: http://editors.eol.org/eoearth/wiki/Deserts_of_North_America (accessed on 14 March 2021).
- NASA LP DAAC. MODIS NDVI Data Version 6; NASA LP DAAC: Sioux Falls, SD, USA, 2016. [Google Scholar]
- PRISM Climate Group. PRISM Gridded Climate Data; PRISM Climate Group: Corvallis, OR, USA, 2016. [Google Scholar]
- Thornthwaite, C.W. An Approach toward a Rational Classification of Climate. Geogr. Rev. 1948, 38, 55. [Google Scholar] [CrossRef]
- Beguería, S.; Vicente-Serrano, S.M.; Reig, F.; Latorre, B. Standardized precipitation evapotranspiration index (SPEI) revisited: Parameter fitting, evapotranspiration models, tools, datasets and drought monitoring. Int. J. Climatol. 2014, 34, 3001–3023. [Google Scholar] [CrossRef] [Green Version]
- Begueria, S.; Vicente-Serrano, S.M. SPEI: Calculation of the Standardised Precipitation-Evapotranspiration Index. R Package Version 1.3. 2013. Available online: https://cran.r-project.org/web/packages/SPEI/index.html (accessed on 14 March 2021).
- Cayan, D.R.; Das, T.; Pierce, D.W.; Barnett, T.P.; Tyree, M.; Gershunova, A. Future dryness in the Southwest US and the hydrology of the early 21st century drought. Proc. Natl. Acad. Sci. USA 2010, 107, 21271–21276. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhou, T. Responses of vegetation growth to climate change in china. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. ISPRS Arch. 2015, 40, 225–229. [Google Scholar] [CrossRef] [Green Version]
- El-Vilaly, M.A.S.; Didan, K.; Marsh, S.E.; Crimmins, M.A.; Munoz, A.B. Characterizing Drought Effects on Vegetation Productivity in the Four Corners Region of the US Southwest. Sustainability 2018, 10, 1643. [Google Scholar] [CrossRef] [Green Version]
- Seager, R.; Ting, M.; Held, I.; Kushnir, Y.; Lu, J.; Vecchi, G.; Huang, H.P.; Harnik, N.; Leetmaa, A.; Lau, N.C.; et al. Model projections of an imminent transition to a more arid climate in southwestern North America. Science 2007, 316, 1181–1184. [Google Scholar] [CrossRef] [PubMed]
- Breshears, D.D.; Cobb, N.S.; Rich, P.M.; Price, K.P.; Allen, C.D.; Balice, R.G.; Romme, W.H.; Kastens, J.H.; Floyd, M.L.; Belnap, J.; et al. Regional vegetation die-off in response to global-change-type drought. Proc. Natl. Acad. Sci. USA 2005, 102, 15144–15148. [Google Scholar] [CrossRef] [Green Version]
- Westerling, A.L.; Hidalgo, H.G.; Cayan, D.R.; Swetnam, T.W. Warming and earlier spring increase western U.S. forest wildfire activity. Science 2006, 313, 940–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, H. Natural savannahs as a transition to the arid zone. Ecol. Trop. Subtrop. Veg. 1971, 238–265. [Google Scholar]
- Munson, S.M.; Webb, R.H.; Belnap, J.; Hubbard, J.A.; Swann, D.E.; Rutman, S. Forecasting climate change impacts to plant community composition in the Sonoran Desert region. Glob. Chang. Biol. 2012, 18, 1083–1095. [Google Scholar] [CrossRef]
- Smith, S.D.; Monson, R.K.; Anderson, J.E. Physiological Ecology of North. American Desert Plants; Springer: Berlin/Heidelberg, Germany, 1997. [Google Scholar]
- Munson, S.M.; Webb, R.H.; Housman, D.C.; Veblen, K.E.; Nussear, K.E.; Beever, E.A.; Hartney, K.B.; Miriti, M.N.; Phillips, S.L.; Fulton, R.E.; et al. Long-term plant responses to climate are moderated by biophysical attributes in a North American desert. J. Ecol. 2015, 103, 657–668. [Google Scholar] [CrossRef]
- Kemp, P.R. Phenological Patterns of Chihuahuan Desert Plants in Relation to the Timing of Water Availability. J. Ecol. 1983, 71, 427. [Google Scholar] [CrossRef]
- Reynolds, J.F.; Virginia, R.A.; Kemp, P.R.; De Soyza, A.G.; Tremmel, D.C. Impact of drought on desert shrubs: Effects of seasonality and degree of resource island development. Ecol. Monogr. 1999, 69, 69–106. [Google Scholar] [CrossRef]
- Shreve, F.; Wiggins, I. Vegetation and Flora of the Sonoran Desert; Stanford University Press: Stanford, CA, USA, 1964. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SPEI Timescale | Ecoregions | Slope | r2 |
|---|---|---|---|
| 1 month—SPEI | Mojave | −1.77 × 10−5 | 0.021 |
| 3 month—SPEI | Mojave | −2.6 × 10−5 | 0.044 |
| 6 month—SPEI | Mojave | −3.26 × 10−5 | 0.071 |
| 12 month—SPEI | Mojave | −3.97 × 10−5 | 0.102 |
| 1 month—SPEI | Sonoran | −3.18 × 10−5 | 0.063 |
| 3 month—SPEI | Sonoran | −4.69 × 10−5 | 0.140 |
| 6 month—SPEI | Sonoran | −5.98 × 10−5 | 0.200 |
| 12 month—SPEI | Sonoran | −7.29 × 10−5 | 0.300 |
| 1 month—SPEI | Apache | −1.46 × 10−5 | 0.013 |
| 3 month—SPEI | Apache | −1.74 × 10−5 | 0.018 |
| 6 month—SPEI | Apache | −2.1 × 10−5 | 0.025 |
| 12 month—SPEI | Apache | −2.67 × 10−5 | 0.041 |
| 1 month—SPEI | Chihuahuan | −7.62 × 10−6 | 0.005 |
| 3 month—SPEI | Chihuahuan | −9.5 × 10−6 | 0.007 |
| 6 month—SPEI | Chihuahuan | −1.02 × 10−5 | 0.008 |
| 12 month—SPEI | Chihuahuan | −1.02 × 10−5 | 0.007 |
| Ecoregion | Historic Mean SPEI (1950–1999) | 21st Century Mean SPEI (2000–2015) |
|---|---|---|
| Mojave | 0.190 | −0.658 |
| Sonoran | 0.303 | −1.038 |
| Apache Highlands | 0.218 | −0.592 |
| Chihuahuan | 0.184 | −0.493 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khatri-Chhetri, P.; Hendryx, S.M.; Hartfield, K.A.; Crimmins, M.A.; Leeuwen, W.J.D.v.; Kane, V.R. Assessing Vegetation Response to Multi-Scalar Drought across the Mojave, Sonoran, Chihuahuan Deserts and Apache Highlands in the Southwest United States. Remote Sens. 2021, 13, 1103. https://doi.org/10.3390/rs13061103
Khatri-Chhetri P, Hendryx SM, Hartfield KA, Crimmins MA, Leeuwen WJDv, Kane VR. Assessing Vegetation Response to Multi-Scalar Drought across the Mojave, Sonoran, Chihuahuan Deserts and Apache Highlands in the Southwest United States. Remote Sensing. 2021; 13(6):1103. https://doi.org/10.3390/rs13061103
Chicago/Turabian StyleKhatri-Chhetri, Pratima, Sean M. Hendryx, Kyle A. Hartfield, Michael A. Crimmins, Willem J. D. van Leeuwen, and Van R. Kane. 2021. "Assessing Vegetation Response to Multi-Scalar Drought across the Mojave, Sonoran, Chihuahuan Deserts and Apache Highlands in the Southwest United States" Remote Sensing 13, no. 6: 1103. https://doi.org/10.3390/rs13061103
APA StyleKhatri-Chhetri, P., Hendryx, S. M., Hartfield, K. A., Crimmins, M. A., Leeuwen, W. J. D. v., & Kane, V. R. (2021). Assessing Vegetation Response to Multi-Scalar Drought across the Mojave, Sonoran, Chihuahuan Deserts and Apache Highlands in the Southwest United States. Remote Sensing, 13(6), 1103. https://doi.org/10.3390/rs13061103