
Bio-Optical Measurements Indicative of Biogeochemical Transformations of Ocean Waters by Coral Reefs

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Variability in the Optical Properties of Coral Waters
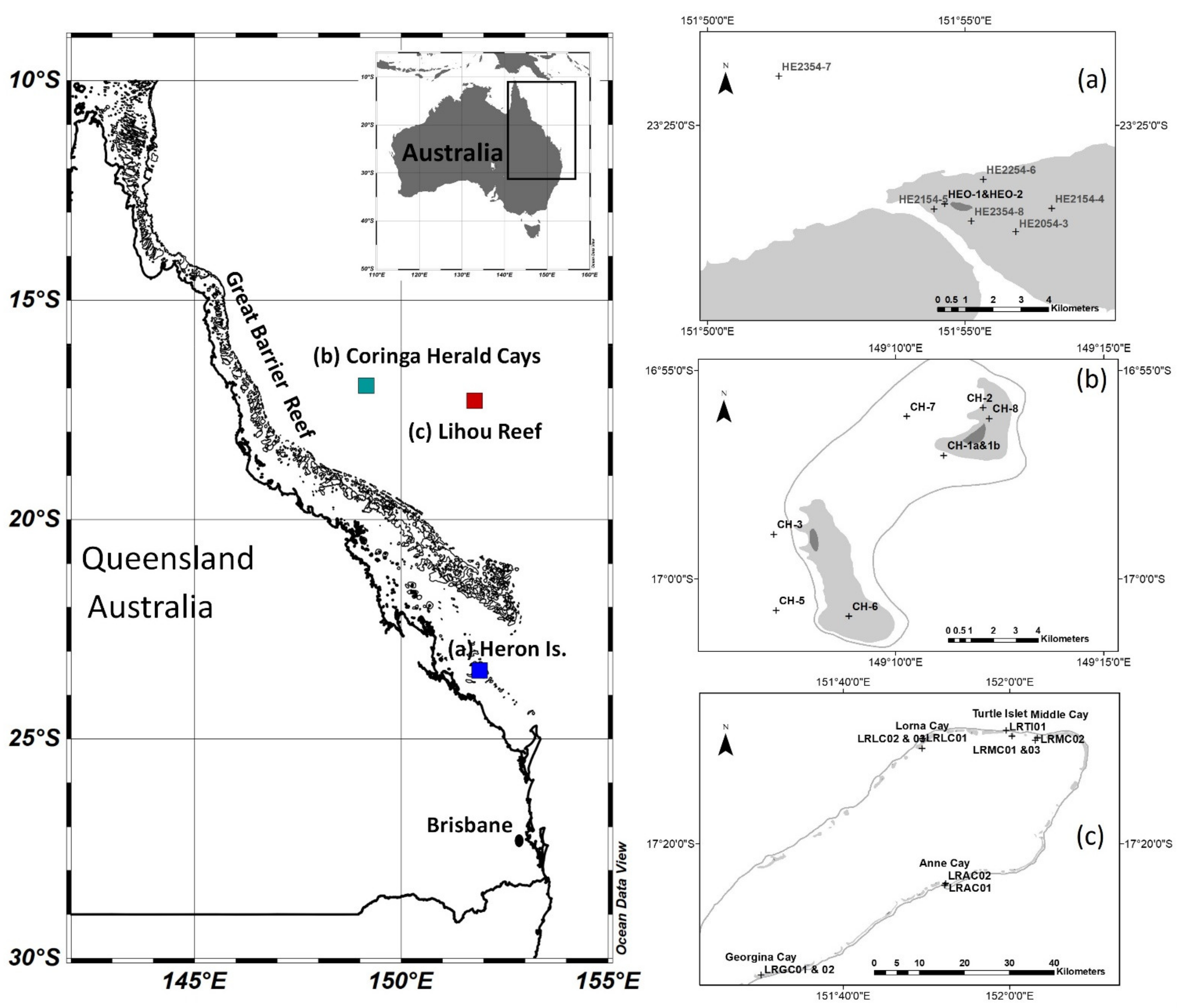
3. Study Areas and Measurements
3.1. Heron Island
3.2. Coringa-Herald
3.3. Lihou Reef
3.4. Sampling and Laboratory Measurements
3.5. Pigment Analysis
3.6. Particulate and Non-Algal Absorption
3.7. CDOM Absorption
3.8. Total Suspended Matter (TSM) Analysis
4. Results
5. Conclusions
5.1. Variability in Biogeochemical and Optical Properties in Coral Reef vs. Open Ocean Waters
5.2. Implications for Underwater Light Climate Models for Remote Sensing of Coral Reefs
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hoegh-Guldberg, O.; Poloczanska, E.S.; Skirving, W.; Dove, S. Coral Reef Ecosystems under Climate Change and Ocean Acidification. Front. Mar. Sci. 2017, 4, 158. [Google Scholar] [CrossRef] [Green Version]
- Albright, R.; Benthuysen, J.; Cantin, N.; Caldeira, K.; Anthony, K. Coral reef metabolism and carbon chemistry dynamics of a coral reef flat, Geophys. Res. Lett. 2015, 42, 3980–3988. [Google Scholar] [CrossRef]
- Pawlik, J.R.; McMurray, S.E. The Emerging Ecological and Biogeochemical Importance of Sponges on Coral Reefs. Annu. Rev. Mar. Sci. 2020, 12, 315–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polónia, A.R.M.; Cleary, D.F.R.; de Voogd, N.J.; Renema, W.; Hoeksema, B.W.; Martins, A.; Gomes, N.C.M. Habitat and water quality variables as predictors of community composition in an Indonesian coral reef: A multi-taxon study in the Spermonde Archipelago. Sci. Total Environ. 2015, 537, 139–151. [Google Scholar] [CrossRef]
- Varunan, T.; Shanmugam, P. Modelling the optical properties of coral reefs and its implication for optical remote sensing. Estuar. Coast. Shelf Sci. 2021, 249, 107054. [Google Scholar] [CrossRef]
- Weeks, S.; Werdell, P.J.; Schaffelke, B.; Canto, M.; Lee, Z.; Wilding, J.G.; Feldman, G.C. Satellite-derived photic depth on the great barrier reef: Spatio-temporal patterns of water clarity. Remote Sens. 2012, 4, 3781–3795. [Google Scholar] [CrossRef] [Green Version]
- Reichstetter, M.; Fearns, P.R.; Weeks, S.J.; McKinna, L.I.; Roelfsema, C.; Furnas, M. Bottom reflectance in ocean color satellite remote sensing for coral reef environments. Remote Sens. 2015, 7, 16756–16777. [Google Scholar] [CrossRef] [Green Version]
- McKinna, L.I.W.; Fearns, P.R.C.; Weeks, S.J.; Werdell, P.J.; Reichstetter, M.; Franz, B.A.; Shea, D.M.; Feldman, G.C. A semi-analytical ocean color inversion algorithm with explicit water column depth and substrate reflectance parameterization. J. Geophys. Res.-Ocean. 2015, 120, 1741–1770. [Google Scholar] [CrossRef] [Green Version]
- Yamano, H.; Tamura, M. Detection limits of coral reef bleaching by satellite remote sensing: Simulation and data analysis. Remote Sens. Environ. 2004, 90, 86–103. [Google Scholar] [CrossRef]
- Goodman, J.; Ustin, S. Airborne hyperspectral analysis of coral reef ecosystems in the Hawaiian Islands. In International Symposium on Remote Sensing of Environment; ISRSE: Honolulu, HI, USA, 2003. [Google Scholar]
- Dekker, A.G.; Phinn, S.R.; Anstee, J.M.; Bissett, P.; Brando, V.E.; Casey, B.; Fearns, P.; Hedley, J.; Klonowski, W.; Lee, Z.P.; et al. Intercomparison of shallow water bathymetry, hydro-optics and benthos mapping techniques in Australian and Caribbean coastal environments. Limnol. Oceanogr. Methods 2011, 9, 396–425. [Google Scholar] [CrossRef] [Green Version]
- Zoffoli, M.L.; Frouin, R.; Kampel, M. Water column correction for coral reef studies by remote sensing. Sensors 2014, 14, 16881–16931. [Google Scholar] [CrossRef] [PubMed]
- Russell, B.J.; Dierssen, M.H.; Hochberg, E.J. Water Column Optical Properties of Pacific Coral Reefs Across Geomorphic Zones and in Comparison to Offshore Waters. Remote Sens. 2019, 757, 1757. [Google Scholar] [CrossRef] [Green Version]
- Maritorena, S.; Guillocheau, N. Optical properties of water and spectral light absorption by living and non-living particles and by yellow substances in coral reef waters of French Polynesia. Mar. Ecol. Prog. Ser. 1996, 131, 245–255. [Google Scholar] [CrossRef]
- Su, H.; Liu, H.; Heyman, W.D. Automated Derivation of Bathymetric Information from Multi-Spectral Satellite Imagery Using a Non-Linear Inversion Model. Mar. Geod. 2008, 4, 281–298. [Google Scholar] [CrossRef]
- Goodman, J.A.; Lee, Z.P.; Ustin, S.L. Influence of atmospheric and sea-surface corrections on retrieval of bottom depth and reflectance using a semi-analytical model: A case study in Kaneohe Bay, Hawaii. Appl. Opt. 2008, 28, F1–F12. [Google Scholar] [CrossRef] [Green Version]
- Kerr, J.M.; Purkis, S. An algorithm for optically-deriving water depth from multispectral imagery in coral reef landscapes in the absence of ground-truth data. Remote Sens. Environ. 2018, 210, 307–324. [Google Scholar] [CrossRef]
- Wei, J.; Lee, Z.; Ondrusek, M.; Mannino, A.; Tzortziou, M.; Armstrong, R. Spectral slopes of the absorption coefficient of colored dissolved and detrital material inverted from UV-visible remote sensing reflectance. J. Geophys. Res. Ocean. 2016, 121, 1953–1969. [Google Scholar] [CrossRef]
- Kutser, T.; Dekker, A.G.; Skirving, W. Modeling spectral discrimination of Great Barrier Reef benthic communities by remote sensing instruments. Limnol. Oceanogr. 2003, 48, 497–510. [Google Scholar] [CrossRef] [Green Version]
- Hochberg, E.J.; Atkinson, M.J.; Andréfouët, S. Spectral reflectance of coral reef bottom-types worldwide and implications for coral reef remote sensing. Remote Sens. Environ. 2003, 85, 159–173. [Google Scholar] [CrossRef]
- Boss, E.; Zaneveld, J.R.V. The effect of bottom substrate on inherent optical properties: Evidence of biogeochemical processes. Limnol. Oceanogr. 2003, 48, 346–354. [Google Scholar] [CrossRef] [Green Version]
- Hedley, J.; Roelfsema, C.; Koetz, B.; Phinn, S. Capability of the Sentinel-2 mission for tropical coral reef mapping and coral beaching detection. Remote Sens. Environ. 2012, 120, 145–155. [Google Scholar] [CrossRef]
- Lesser, M.; Mobley, C. Bathymetry, water optical properties, and benthic classification of coral reefs using hyperspectral remote sensing imagery. Coral Reefs 2007, 26, 819–829. [Google Scholar] [CrossRef]
- Klonowski, W.M.; Fearns, P.R.C.S.; Lynch, M.J. Retrieving key benthic cover types and bathymetry from hyperspectral imagery. J. Appl. Rem. Sens. 2007, 1, 011505. [Google Scholar] [CrossRef]
- Petit, T.; Bajjouk, T.; Mouquet, P.; Rochette, S.; Vozel, B.; Delacourt, C. Hyperspectral remote sensing of coral reefs by semi-analytical model inversion-Comparison of different inversion setups. Remote Sens. Environ. 2017, 190, 348–365. [Google Scholar] [CrossRef]
- Hedley, J.; Mirhakak, E.M.; Wentworth, A.; Dierssen, H. Influence of Three-Dimensional Coral Structures on Hyperspectral Benthic Reflectance and Water-Leaving Reflectance. Appl. Sci. 2018, 8, 2688. [Google Scholar] [CrossRef] [Green Version]
- Dekker, A.G.; Botha, E.J.; Park, Y.J.; Anstee, J.M.; Cherukuru, N.; Clementson, L. Remote sensing for habitat mapping and change detection in tropical Commonwealth marine protected areas—Phase 2. In Report Prepared for: The Department of Environment, Water, Heritage and the Arts, National Research Flagships, Wealth from Oceans; CSIRO: Canberra, Australia, 2010; p. 135. [Google Scholar]
- Clementson, L.A. The CSIRO Method. In The Fifth SeaWiFS HPLC Analysis Round-Robin Experiment (SeaHARRE-5), NASA Technical Memorandum 2012–217503; Hooker, S.B., Clementson, L.A., Thomas, C.S., Schlüter, L., Allerup, M., Ras, J., Claustre, H., Normandeau, C., Cullen, J., Kienast, M., et al., Eds.; NASA Goddard Space Flight Center: Greenbelt, MD, USA, 2013. [Google Scholar]
- Kishino, M.; Takahashi, M.; Okami, N.; Ichimura, S. Estimation of the spectral absorption-coefficients of phytoplankton in the Sea. Bull. Mar. Sci. 1985, 37, 634–642. [Google Scholar]
- Mitchell, B.G. Algorithms for determining the absorption coefficient for aquatic particulates using the quantitative filter technique. Proc. Ocean Opt. X 1990, 1302, 137–148. [Google Scholar]
- Tilstone, G.H.; Moore, G.F.; Sørensen, K.; Doerffer, R.; Røttgers, R.; Ruddick, K.G.; Pasterkamp, R.; Jørgensen, V. REVAMP Regional Validation of MERIS Chlorophyll Products in North Sea Coastal Waters Report; EU funded ENVISAT-1 Project. Proposal: EVG2–2001–00009; Contract: EVG1–CT–2001–00049 Protocols Document; Plymouth Marine Laboratories: Plymouth, UK, 2002. [Google Scholar]
- Buiteveld, H. A model for calculation of diffuse light attenuation (PAR) and Secchi depth. Neth. J. Aquat. Ecol. 1995, 29, 55–65. [Google Scholar] [CrossRef]
- Silva, S.C.; Ferreira, I.; Dias, M.M.; Barreiro, M.F. Microalgae-Derived Pigments: A 10-Year Bibliometric Review and Industry and Market Trend Analysis. Molecules 2020, 25, 3406. [Google Scholar] [CrossRef]
- Blondeau-Patissier, D.; Brando, V.E.; Oubelkheir, K.; Dekker, A.G.; Clementson, L.A.; Daniel, P. Bio-optical variability of the absorption and scattering properties of the Queensland inshore and reef waters, Australia. J. Geophys. Res. 2009, 114, C05003. [Google Scholar] [CrossRef]
- Vidussi, F.; Claustre, H.; Manca, B.; Luchetta, A.; Marty, J.-C. Phytoplankton pigment distribution in relation to upper thermocline circulation in the Eastern Mediterranean Sea during winter. J. Geophys. Res. 2001, 106, 19939–19956. [Google Scholar] [CrossRef]
- Uitz, J.; Claustre, H.; Morel, A.; Hooker, S.B. Vertical distribution of phytoplankton communities in open ocean: An assessment based on surface chlorophyll. J. Geophys. Res. Ocean. 2006, 111, C8. [Google Scholar] [CrossRef]
- Brando, V.E.; Dekker, A.G.; Park, Y.J.; Schroeder, T. An adaptive semi-analytical inversion of ocean colour radiometry in optically complex waters. Appl. Opt. 2012, 51, 2808–2833. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reef | Ocean | Ratio | |
|---|---|---|---|
| Mean ± STD (Min–Max) | Mean ± STD (Min–Max) | Mean R:O | |
| Chlorophyll a (mg m−3) | |||
| Heron Island | 0.113 ± 0.090 (0.030–0.217) | 0.386 ± 0.083 (0.285–0.502) | 0.294 |
| Coringa-Herald | 0.039 ± 0.022 (0.022–0.078) | 0.101 ± 0.023 (0.087–0.128) | 0.389 |
| Lihou Reef | 0.053 ± 0.015 (0.031–0.074) | 0.106 ± 0.023 (0.079–0.137) | 0.498 |
| Non-Algal Particulate (mg L−1) | |||
| Heron Island | 1.232 ± 0.555 (0.686–1.995) | 1.106 ± 1.345 (0.177–3.765) | 1.114 |
| Coringa-Herald | 0.276 ± 0.241 (0.020–0.560) | 0.233 ± 0.303 (0.020–0.580) | 1.183 |
| Lihou Reef | 1.470 ± 0.413 (0.982–2.077) | 1.152 ± 0.338 (0.657–1.655) | 1.276 |
| (a) | |||
| Reef | Ocean | Ratio | |
| Heron Island | Mean ± STD (Min–Max) N = 5 | Mean ± STD (Min–Max) N = 6 | Mean R:O |
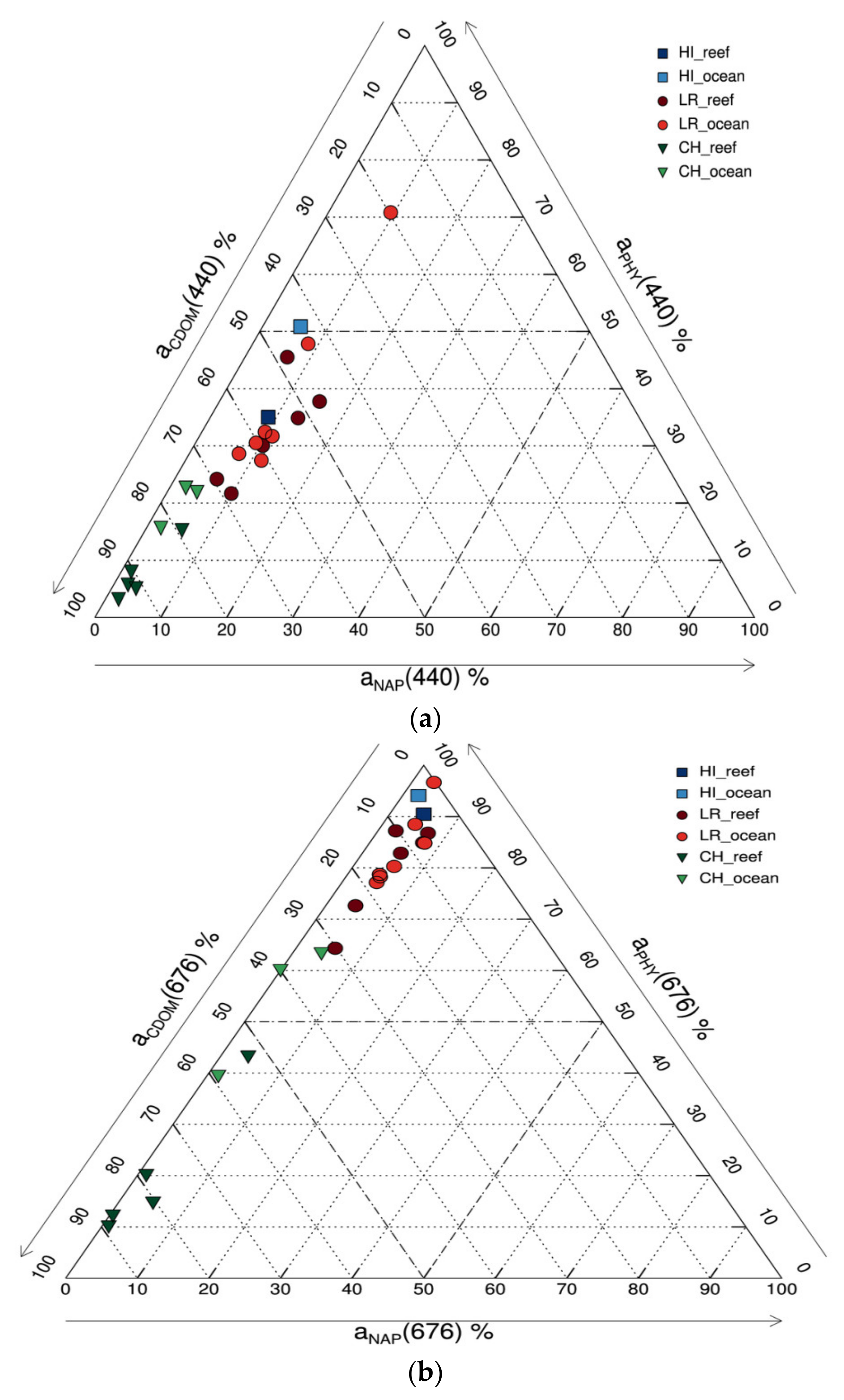
| a(NAP)440 | 0.0054 ± 0.0015 (0.0036–0.0072) | 0.0043 ± 0.0010 (0.0032–0.0055) | 1.274 |
| S(NAP)440 | −0.0099 ± 0.0008 (−0.0108–−0.0087) | −0.0096 ± 0.0012 (−0.0112–−0.0080) | 1.031 |
| a(NAP)*440 | 0.0047 ± 0.0026 (0.0027–0.0091) | 0.0103 ± 0.0104 (0.0015–0.0282) | 0.458 |
| a(CDOM)440 | 0.0231 * | 0.0249 * | 0.925 |
| S(CDOM)440 | −0.0183 | −0.0168 | 1.089 |
| a(PHY)440 | 0.0107 ± 0.0065 (0.0042–0.0200) | 0.0313 ± 0.0046 (0.0275–0.0374) | 0.342 |
| a(PHY)*440 | 0.1108 ± 0.0304 (0.0678–0.1419) | 0.0824 ± 0.0109 (0.0678–0.0999) | 1.345 |
| (b) | |||
| Reef | Ocean | Ratio | |
| Coringa Herald Cays | Mean ± STD (Min–Max) N = 5 | Mean ± STD (Min–Max) N = 3 | Mean R:O |
| a(NAP)440 | 0.0026 ± 0.0008 (0.0018–0.0036) | 0.0016 ± 0.0007 (0.0012–0.0024) | 1.589 |
| S(NAP)440 | −0.0117 ± 0.0014 (−0.0130–−0.0094) | −0.0122 ± 0.0021 (−0.0138–−0.0099) | 0.959 |
| a(NAP)*440 | 0.0291 ± 0.0358 (0.0042–0.0888) | 0.0291 ± 0.0299 (0.0022–0.0613) | 1.001 |
| a(CDOM)440 | 0.0853 ± 0.0215 (0.0510–0.1066) | 0.0415 ± 0.0065 (0.0367–0.0489) | 2.055 |
| S(CDOM)440 | −0.0105 ± 0.0017 (−0.0125–−0.0079) | −0.0125 ± 0.0013 (−0.0136–−0.0111) | 0.840 |
| a(PHY)440 | 0.0065 ± 0.0030 (0.0033–0.0098) | 0.0107 ± 0.0012 (0.0093–0.0116) | 0.608 |
| a(PHY)*440 | 0.1784 ± 0.0802 (0.1218–0.3197 | 0.1081 ± 0.0220 (0.0871–0.1310) | 1.650 |
| (c) | |||
| Reef | Ocean | Ratio | |
| Lihou Reefs | Mean ± STD (Min–Max) N = 6 | Mean ± STD (Min–Max) N = 7 | Mean R:O |
| a(NAP)440 | 0.0247 ± 0.0122 (0.0055–0.0353) | 0.0222 ± 0.0164 (0.0044–0.0522) | 1.113 |
| S(NAP)440 | −0.0099 ± 0.0004 (−0.0107–−0.0095) | −0.0098 ± 0.0005 (−0.0105–−0.0091) | 1.010 |
| a(NAP)*440 | 0.0172 ± 0.0096 (0.0036–0.0321) | 0.0208 ± 0.0144 (0.0032–0.0479) | 0.824 |
| a(CDOM)440 | 0.1354 ± 0.0591 (0.0413–0.2190) | 0.1287 ± 0.0892 (0.0092–0.2767) | 1.052 |
| S(CDOM)440 | −0.0135 ± 0.0016 (−0.0152–−0.0109) | −0.0152 ± 0.0043 (−0.0243–−0.0123) | 0.888 |
| a(PHY)440 | 0.0697 ± 0.0203 (0.0391–0.0880) | 0.0706 ± 0.0327(0.0318–0.1245) | 0.988 |
| a(PHY)*440 | 0.0282 ± 0.0088 (0.0154–0.0422) | 0.0401 ± 0.0173 (0.0138–0.0686) | 0.703 |
| (d) | |||
| Reef | Ocean | Ratio | |
| Lihou Reefs | Mean ± STD (Min–Max) N = 5 | Mean ± STD (Min–Max) N = 6 | Mean R:O |
| a(NAP)440 | 0.0233 ± 0.0131 (0.0055–0.0353) | 0.0172 ± 0.0105 (0.0044–0.0300) | 1.358 |
| S(NAP)440 | −0.0099 ± 0.0005 (−0.0107–−0.0095) | −0.0098 ± 0.0006 (−0.0105–−0.0091) | 1.010 |
| a(NAP)*440 | 0.0142 ± 0.0069 (0.0036–0.0215) | 0.0163 ± 0.0089 (0.0032–0.0263) | 0.868 |
| a(CDOM)440 | 0.1187 ± 0.0477 (0.0413–0.1623) | 0.1040 ± 0.0666 (0.0092–0.1554) | 1.141 |
| S(CDOM)440 | −0.0141 ± 0.0011 (−0.0152–−0.0127) | −0.0149 ± 0.0047 (−0.0243–−0.0123) | 0.946 |
| a(PHY)440 | 0.0698 ± 0.0226 (0.0391–0.0880) | 0.0616 ± 0.0246 (0.0318–0.0864) | 1.134 |
| a(PHY)*440 | 0.0254 ± 0.0062 (0.0154–0.0318) | 0.0354 ± 0.0130 (0.0138–0.0508) | 0.718 |
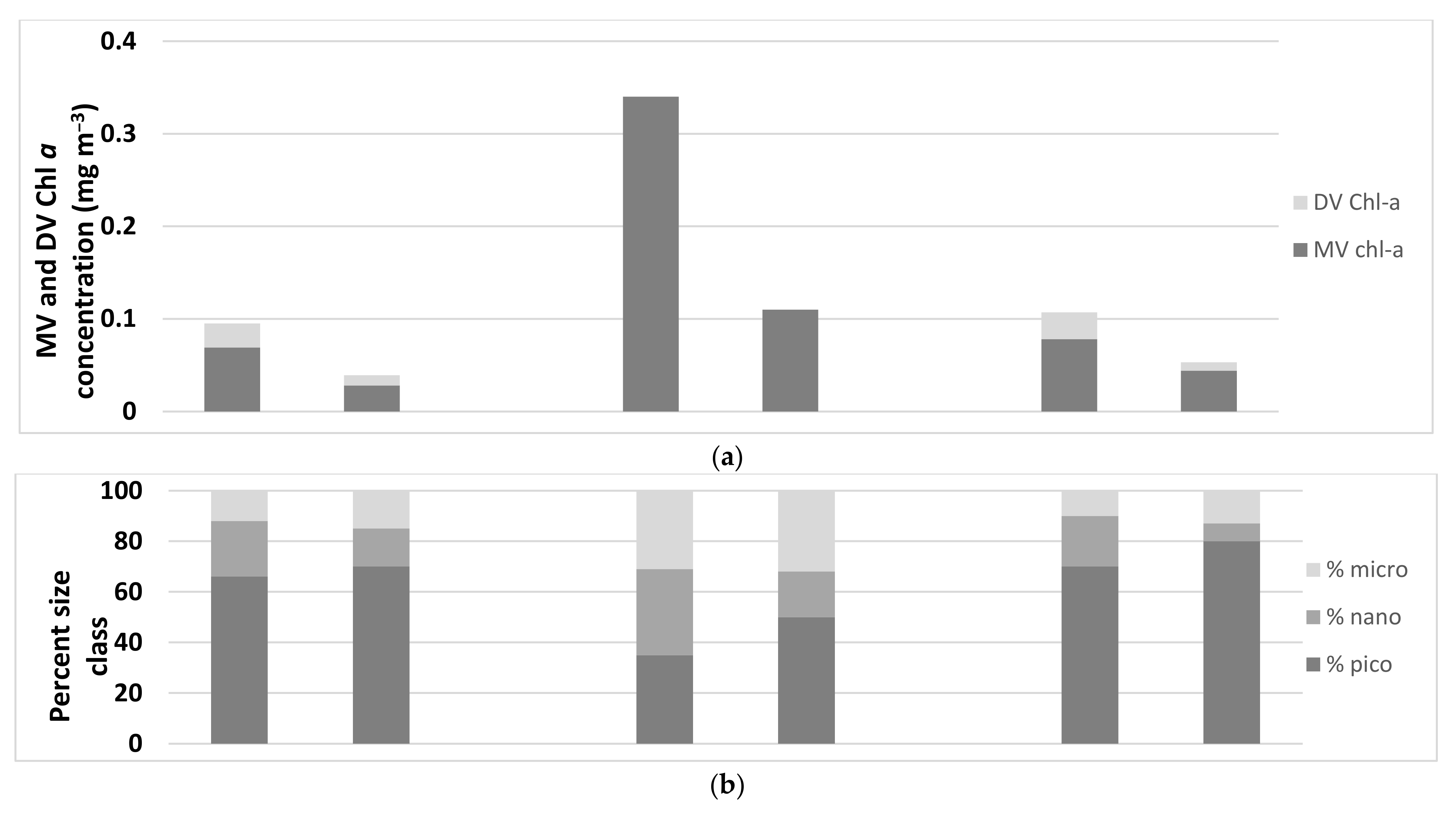
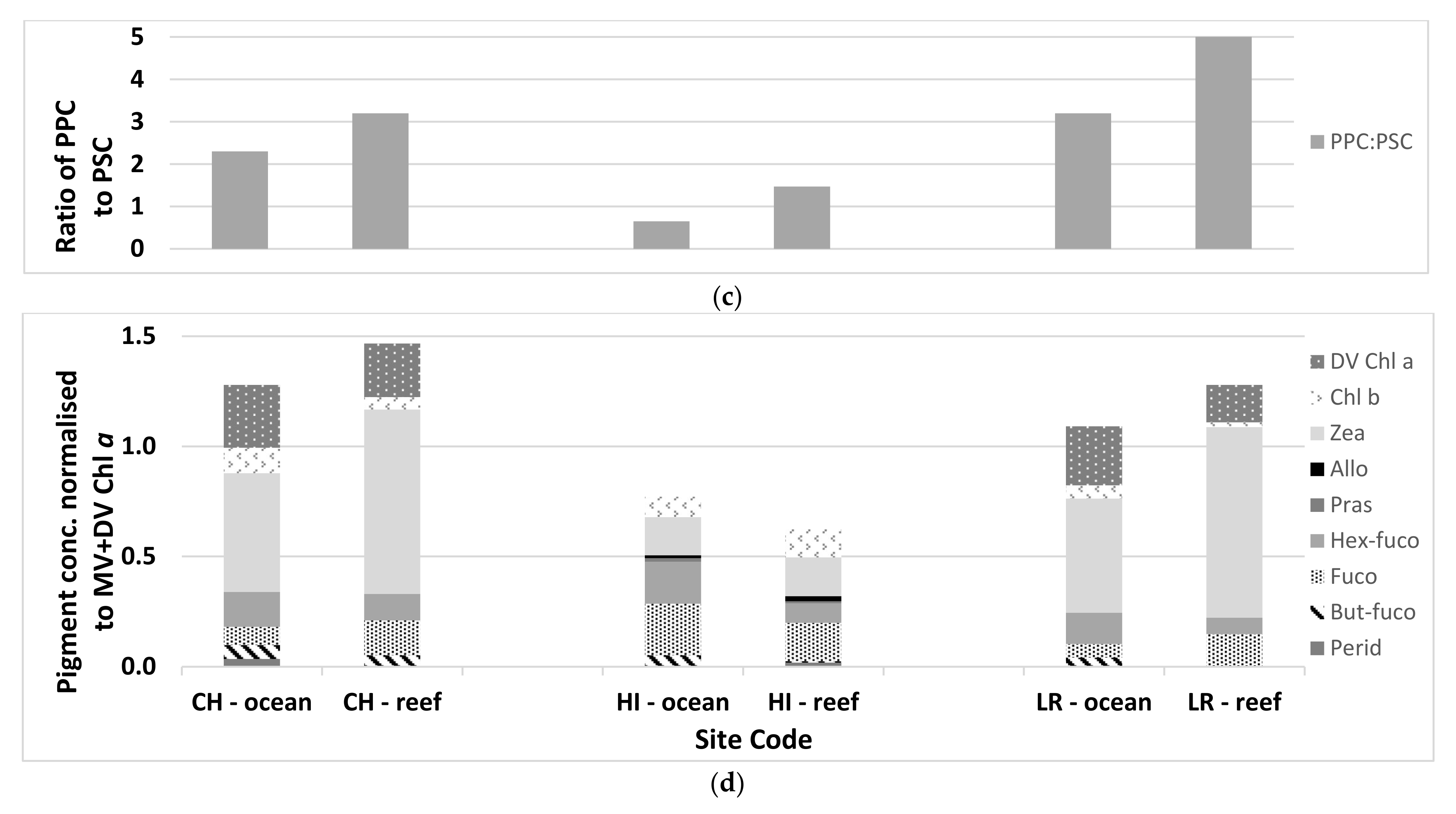
| Total Chl a (mg m−3) | Dominant Phytoplankton | Picoplankton | Size Class (%) Nanoplankton | Microplankton | PPC:PSC | |
|---|---|---|---|---|---|---|
| Heron Island | ||||||
| Ocean waters | 0.39 ± 0.08 | Cyanobacteria, haptophytes, diatoms | 35 ± 3 | 34 ± 4 | 31 ± 6 | 0.7 ± 0.1 |
| Reef waters | 0.11 ± 0.09 | Cyanobacteria, haptophytes, diatoms | 50 ± 9 | 18 ± 11 | 32 ± 4 | 1.5 ± 0.6 |
| Coringa Herald Reef | ||||||
| Ocean waters | 0.101 ± 0.023 | Cyanobacteria | 66 ± 3 | 22 ± 5 | 12 ± 4 | 2.3 ± 0.6 |
| Reef waters | 0.039 ± 0.022 | Cyanobacteria | 70 ± 12 | 15 ± 8 | 15 ± 10 | 3.2 ± 1.5 |
| Lihou Reef | ||||||
| Ocean waters | 0.106 ± 0.023 | Cyanobacteria | 70 ± 4 | 23 ± 4 | 10 ± 9 | 3.2 ± 0.7 |
| Reef waters | 0.053 ± 0.015 | Cyanobacteria | 80 ± 4 | 7 ± 6 | 13 ± 3 | 5.3 ± 1.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dekker, A.G.; Clementson, L.A.; Wettle, M.; Cherukuru, N.; Botha, H.; Oubelkheir, K. Bio-Optical Measurements Indicative of Biogeochemical Transformations of Ocean Waters by Coral Reefs. Remote Sens. 2022, 14, 2892. https://doi.org/10.3390/rs14122892
Dekker AG, Clementson LA, Wettle M, Cherukuru N, Botha H, Oubelkheir K. Bio-Optical Measurements Indicative of Biogeochemical Transformations of Ocean Waters by Coral Reefs. Remote Sensing. 2022; 14(12):2892. https://doi.org/10.3390/rs14122892
Chicago/Turabian StyleDekker, Arnold G., Lesley A. Clementson, Magnus Wettle, Nagur Cherukuru, Hannelie Botha, and Kadija Oubelkheir. 2022. "Bio-Optical Measurements Indicative of Biogeochemical Transformations of Ocean Waters by Coral Reefs" Remote Sensing 14, no. 12: 2892. https://doi.org/10.3390/rs14122892
APA StyleDekker, A. G., Clementson, L. A., Wettle, M., Cherukuru, N., Botha, H., & Oubelkheir, K. (2022). Bio-Optical Measurements Indicative of Biogeochemical Transformations of Ocean Waters by Coral Reefs. Remote Sensing, 14(12), 2892. https://doi.org/10.3390/rs14122892