Species Richness (of Insects) Drives the Use of Acoustic Space in the Tropics


 and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Sites and Acoustic Data Collection
2.2. Regional Species Richness
2.3. Acoustic Morphospecies Richness and Composition in Recordings
2.4. Acoustic Space Use (ASU)
3. Results
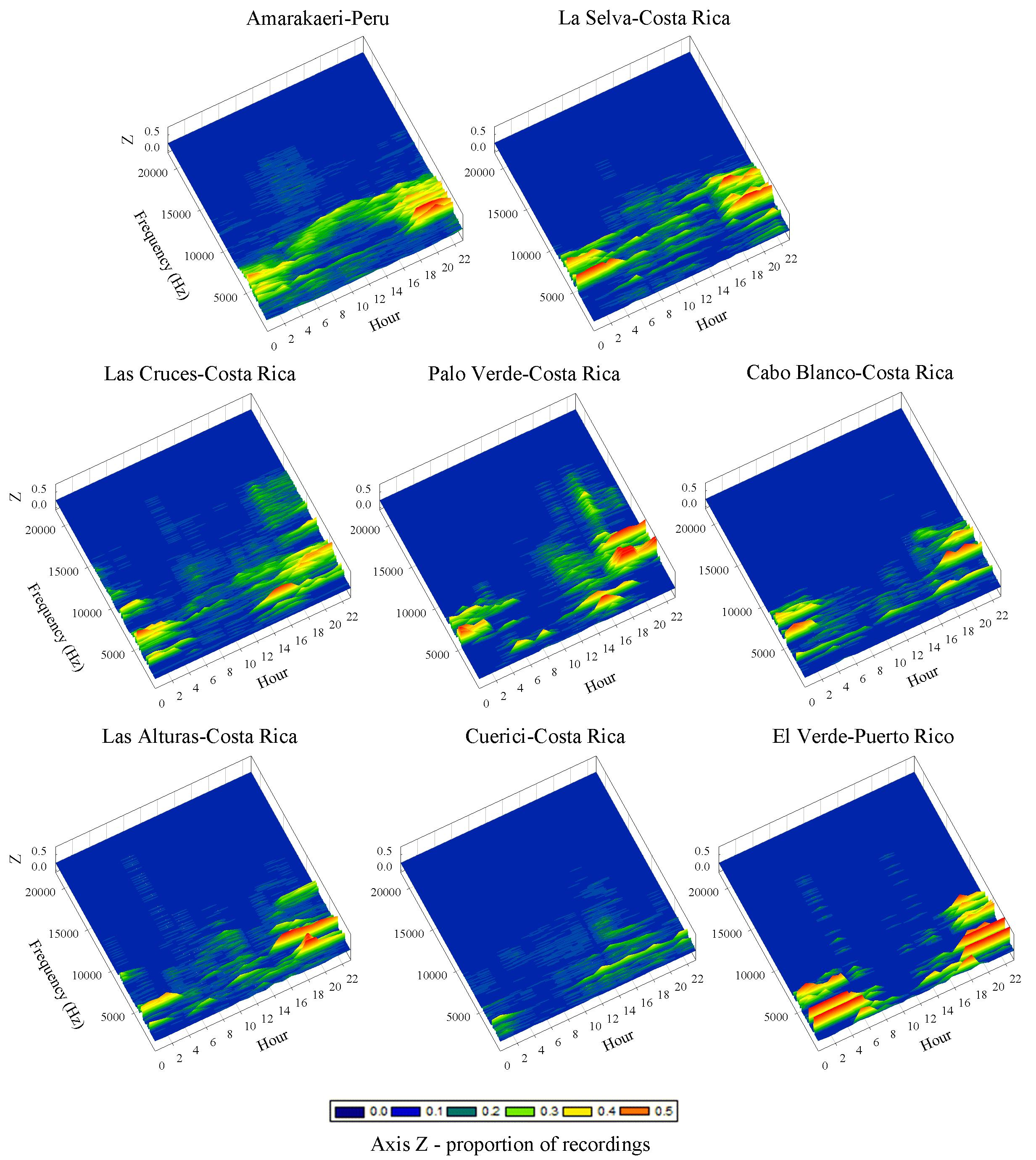
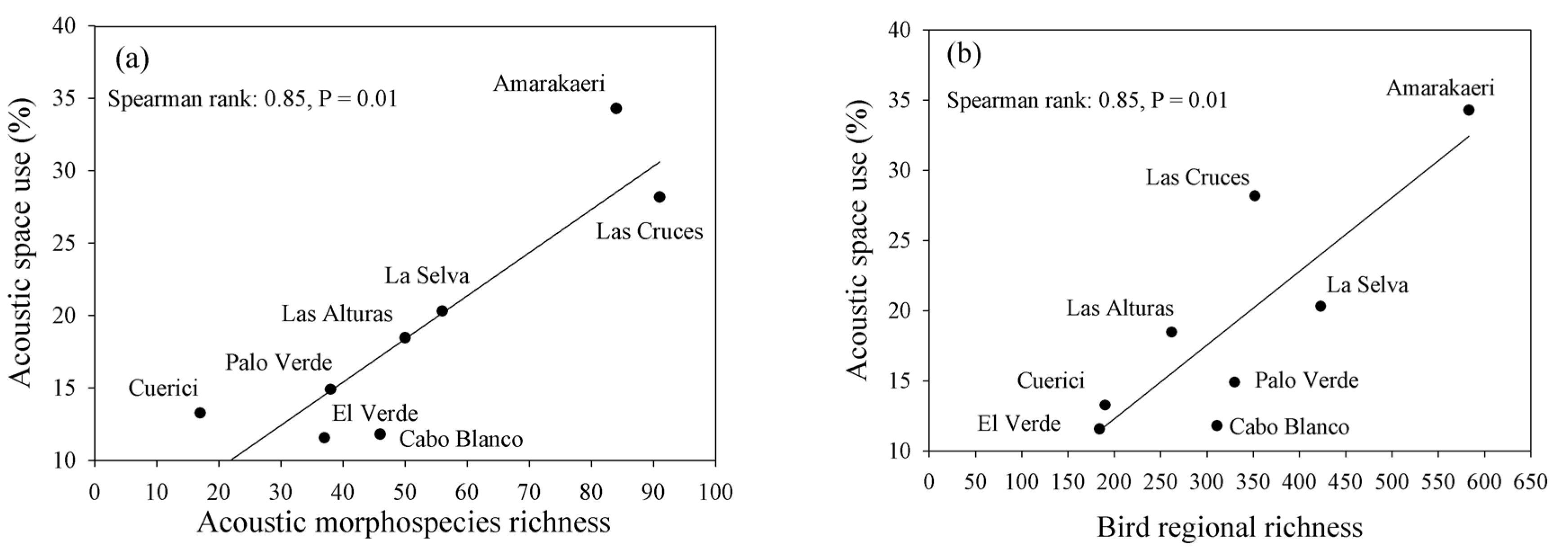
3.1. Species Richness and Acoustic Space Use (ASU)
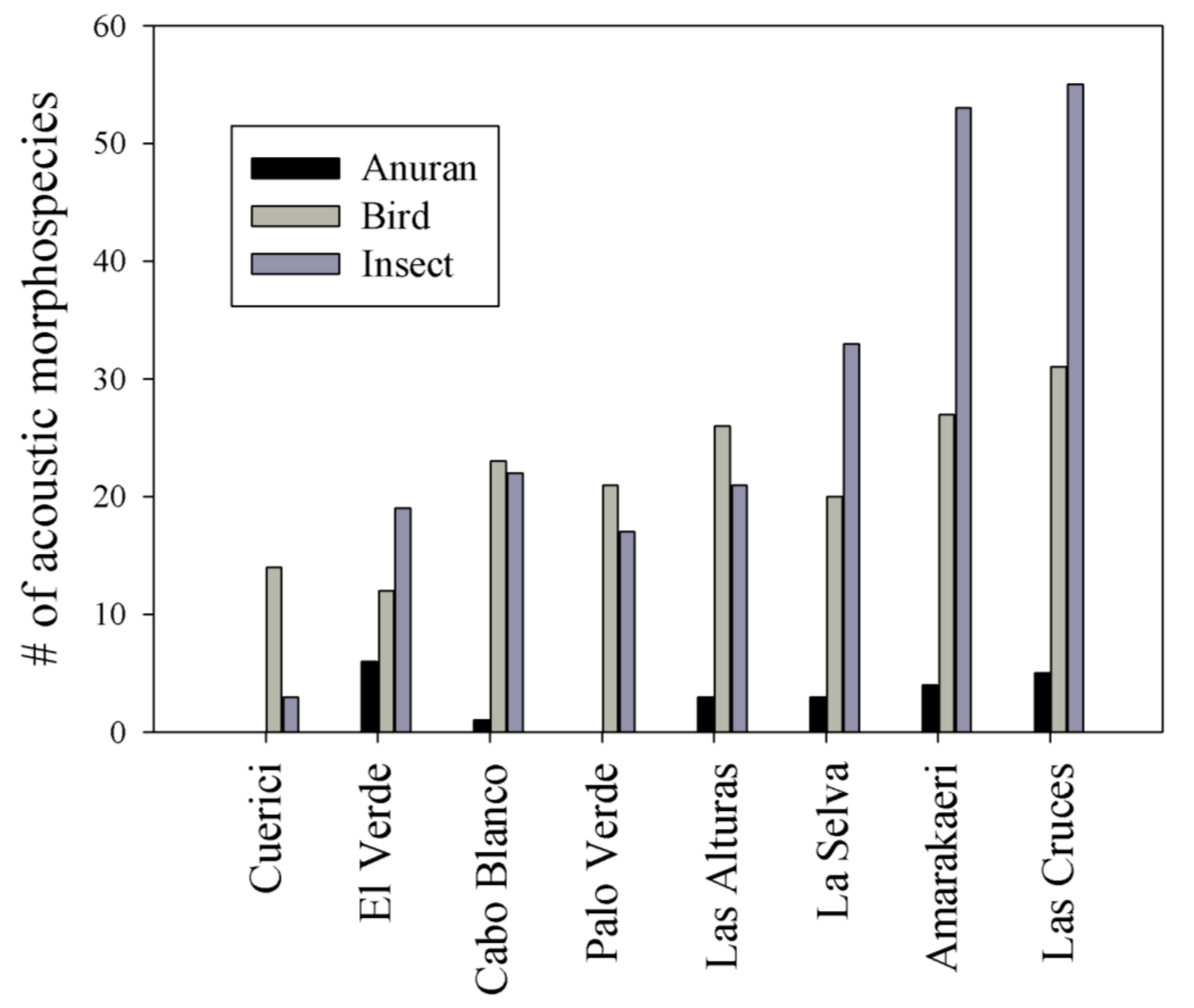
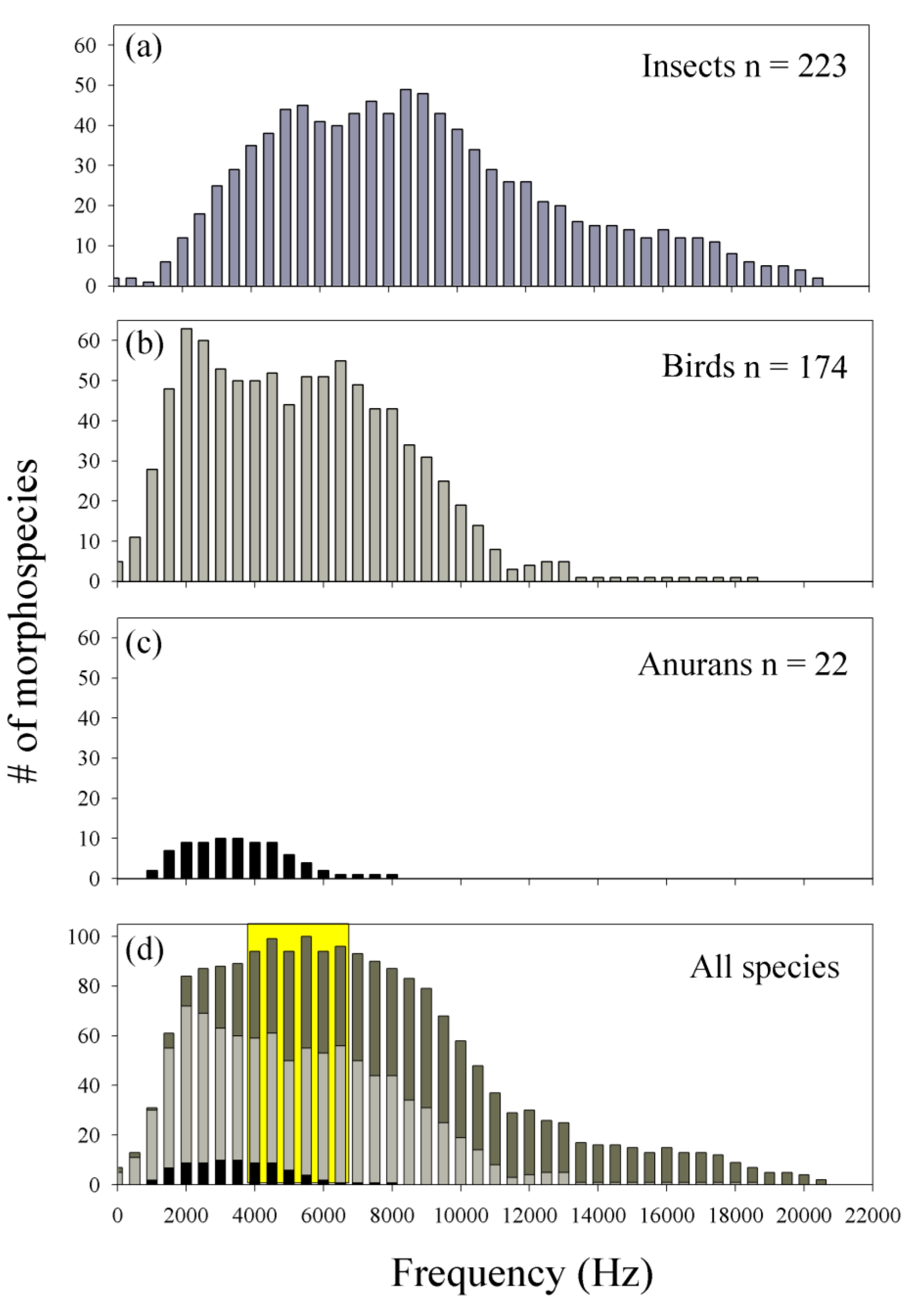
3.2. Acoustic Morphospecies Composition and Acoustic Space Use (ASU)
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

References
- Krause, B. The niche hypothesis: How animals taught us to dance and sing. Whole Earth Rev. 1987, 57, 1–6. [Google Scholar]
- Krause, B. The niche hypothesis: A virtual symphony of animal sounds, the origins of musical expression and the health of habitats. Soundscape Newsl. 1993, 6, 1–5. [Google Scholar]
- Mendelson, T.C.; Shaw, K.L. The (mis) concept of species recognition. Trends Ecol. Evol. 2012, 27, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Irwin, D.E.; Bensch, S.; Price, T.D. Speciation in a ring. Nature 2001, 409, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Tobias, J.A.; Seddon, N. Signal design and perception in Hypocnemis antbirds: Evidence for convergent evolution via social selection. Evolution 2009, 63, 3168–3189. [Google Scholar] [CrossRef] [PubMed]
- Morton, E.S. Ecological sources of selection on avian sounds. Am. Nat. 1975, 109, 17–34. [Google Scholar] [CrossRef]
- Ryan, M.J.; Brenowitz, E.A. The role of body size, phylogeny, and ambient noise in the evolution of bird song. Am. Nat. 1985, 126, 87. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Seddon, N.; Safran, R.J. Evolutionary divergence in acoustic signals: Causes and consequences. Trends Ecol. Evol. 2013, 28, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Sueur, J. Cicada acoustic communication: Potential sound partitioning in a multispecies community from Mexico (Hemiptera: Cicadomorpha: Cicadidae). Biol. J. Linn. Soc. 2002, 75, 379–394. [Google Scholar] [CrossRef]
- Chek, A.A.; Bogart, J.P.; Lougheed, S.C. Mating signal partitioning in multi-species assemblages: A null model test using frogs. Ecol. Lett. 2003, 6, 235–247. [Google Scholar] [CrossRef]
- Schmidt, A.K.D.; Römer, H.; Riede, K. Spectral niche segregation and community organization in a tropical cricket assemblage. Behav. Ecol. 2013, 24, 470–480. [Google Scholar] [CrossRef]
- Villanueva-Rivera, L.J. Eleutherodactylus frogs show frequency but no temporal partitioning: Implications for the acoustic niche hypothesis. PeerJ 2014, 2, e496. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.K.D.; Balakrishnan, R. Ecology of acoustic signalling and the problem of masking interference in insects. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2014, 201, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Ruppé, L.; Clément, G.; Herrel, A.; Ballesta, L.; Décamps, T.; Kéver, L.; Parmentier, E. Environmental constraints drive the partitioning of the soundscape in fishes. Proc. Natl. Acad. Sci. USA 2015, 112, 6092–6097. [Google Scholar] [CrossRef] [PubMed]
- Amézquita, A.; Flechas, S.V.S.; Lima, A.P.; Gasser, H.; Hödl, W. Acoustic interference and recognition space within a complex assemblage of dendrobatid frogs. Proc. Natl. Acad. Sci. USA 2011, 108, 17058–17063. [Google Scholar] [CrossRef] [PubMed]
- Tobias, J.A.; Planqué, R.; Cram, D.L.; Seddon, N. Species interactions and the structure of complex communication networks. Proc. Natl. Acad. Sci. USA 2014, 111, 1020–1025. [Google Scholar] [CrossRef] [PubMed]
- Luther, D. The influence of the acoustic community on songs of birds in a neotropical rain forest. Behav. Ecol. 2009, 20, 864–871. [Google Scholar] [CrossRef]
- Roca, I.T.; Proulx, R. Acoustic assessment of species richness and assembly rules in ensiferan communities from temperate ecosystems. Ecology 2016, 97, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Sueur, J.; Pavoine, S.; Hamerlynck, O.; Duvail, S. Rapid acoustic survey for biodiversity appraisal. PLoS ONE 2008, 3, e4065. [Google Scholar] [CrossRef] [PubMed]
- Depraetere, M.; Pavoine, S.; Jiguet, F.; Gasc, A.; Duvail, S.; Sueur, J. Monitoring animal diversity using acoustic indices: Implementation in a temperate woodland. Ecol. Indic. 2012, 13, 46–54. [Google Scholar] [CrossRef]
- Deichmann, J.L.; Hernández-Serna, A.; Delgado-Cornejo, J.A.; Campos-Cerqueira, M.; Aide, T.M. Soundscape analysis and acoustic monitoring document impacts of natural gas exploration on biodiversity in a tropical forest. Ecol. Indic. 2017, 74, 39–48. [Google Scholar] [CrossRef]
- Campos-Cerqueira, M.; Arendt, W.J.; Wunderle, J.M., Jr.; Aide, T.M. Have bird distributions shifted along an elevational gradient on a tropical mountain? Ecol. Evol. 2017. [Google Scholar] [CrossRef]
- Rowcliffe, M.J.; Carbone, C.; Jansen, P.A.; Kays, R.; Kranstauber, B. Quantifying the sensitivity of camera traps: An adapted distance sampling approach. Methods Ecol. Evol. 2011, 2, 464–476. [Google Scholar] [CrossRef]
- Pekin, B.K.; Jung, J.; Villanueva-Rivera, L.J.; Pijanowski, B.C.; Ahumada, J.A. Modeling acoustic diversity using soundscape recordings and LIDAR-derived metrics of vertical forest structure in a neotropical rainforest. Landsc. Ecol. 2012, 27, 1513–1522. [Google Scholar] [CrossRef]
- Rodriguez, A.; Gasc, A.; Pavoine, S.; Grandcolas, P.; Gaucher, P.; Sueur, J. Temporal and spatial variability of animal sound within a neotropical forest. Ecol. Inform. 2014, 21, 133–143. [Google Scholar] [CrossRef]
- Llusia, D.; Márquez, R.; Bowker, R. Terrestrial sound monitoring systems, a methodology for quantitative calibration. Bioacoustics 2011, 20, 277–286. [Google Scholar] [CrossRef]
- Aide, T.M.; Corrada-Bravo, C.; Campos-Cerqueira, M.; Milan, C.; Vega, G.; Alvarez, R. Real-time bioacoustics monitoring and automated species identification. PeerJ 2013, 1, e103. [Google Scholar] [CrossRef] [PubMed]
- Celis-Murillo, A.; Deppe, J.L.; Allen, M.F. Using soundscape recordings to estimate bird species abundance, richness, and composition. J. Field Ornithol. 2009, 80, 64–78. [Google Scholar] [CrossRef]
- Celis-Murillo, A.; Deppe, J.L.; Ward, M.P. Effectiveness and utility of acoustic recordings for surveying tropical birds. J. Field Ornithol. 2012, 83, 166–179. [Google Scholar] [CrossRef]
- Campos-Cerqueira, M.; Aide, T.M. Improving distribution data of threatened species by combining acoustic monitoring and occupancy modelling. Methods Ecol. Evol. 2016, 7, 1340–1348. [Google Scholar] [CrossRef]
- Sueur, J.; Farina, A.; Gasc, A.; Pieretti, N.; Pavoine, S. Acoustic indices for biodiversity assessment and landscape investigation. Acta Acust. United Acust. 2014, 100, 772–781. [Google Scholar] [CrossRef]
- Ospina, O.E.; Villanueva-Rivera, L.J.; Corrada-Bravo, C.J.; Aide, T.M. Variable response of anuran calling activity to daily precipitation and temperature: Implications for climate change. Ecosphere 2013, 4, 1–12. [Google Scholar] [CrossRef]
- Gasc, A.; Pavoine, S.; Lellouch, L.; Grandcolas, P.; Sueur, J. Acoustic indices for biodiversity assessments: Analyses of bias based on simulated bird assemblages and recommendations for field surveys. Biol. Conserv. 2015, 191, 306–312. [Google Scholar] [CrossRef]
- Jenkins, C.N.; Pimm, S.L.; Joppa, L.N. Global patterns of terrestrial vertebrate diversity and conservation. Proc. Natl. Acad. Sci. USA 2013, 110, E2602–E2610. [Google Scholar] [CrossRef] [PubMed]
- Sueur, J.J.; Aubin, T.; Simonis, C. Seewave, a free modular tool for sound analysis and synthesis. Bioacustics 2008, 18, 213–226. [Google Scholar] [CrossRef]
- Burnham, K.P. Multimodel inference: Understanding AIC and BIC in model selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- Prestwich, K. The energetics of acoustic signalling in anurans and insects. Am. Zool. 1994, 34, 625–643. [Google Scholar] [CrossRef]
- Ophir, A.G.; Schrader, S.B.; Gillooly, J.F. Energetic cost of calling: General constraints and species-specific differences. J. Evol. Biol. 2010, 23, 1564–1569. [Google Scholar] [CrossRef] [PubMed]
- Fay, R.R. Hearing in Vertebrates: A Psychophysics Databook; Hill-Fay: Winnetka, IL, USA, 1988; ISBN 0961855908. [Google Scholar]
- Robinson, D.J.; Hall, M.J. Sound signalling in Orthoptera. In Advances in Insect Physiology; Evans, P., Ed.; Elsevier: Kidlington, UK, 2002; pp. 151–278. ISBN 0065-2806. [Google Scholar]
- Warfield, D. The study of hearing in animals. In Methods of Animal Experimentation, IV; Gay, W., Ed.; Academic Press: London, UK, 2012; pp. 43–143. [Google Scholar]
- Hamilton, A.J.; Basset, Y.; Benke, K.K.; Grimbacher, P.S.; Miller, S.E.; Novotný, V.; Samuelson, G.A.; Stork, N.E.; Weiblen, G.D.; Yen, J.D.L. Quantifying uncertainty in estimation of tropical arthropod species richness. Am. Nat. 2010, 176, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Mora, C.; Tittensor, D.P.; Adl, S.; Simpson, A.G.B.; Worm, B. How many species are there on earth and in the ocean? PLoS Biol. 2011, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Paez, V.P.; Bock, B.C.; Rand, A.S. Inhibition of evoked calling of Dendrobates pumilio due to acoustic interference from cicada calling. Biotropica 1993, 25, 242–245. [Google Scholar] [CrossRef]
- Stanley, C.Q.; Walter, M.H.; Venkatraman, M.X.; Wilkinson, G.S. Insect noise avoidance in the dawn chorus of Neotropical birds. Anim. Behav. 2016, 112, 255–265. [Google Scholar] [CrossRef]
- Riede, K. Monitoring biodiversity: Analysis of Amazonian rainforest sounds. Ambio 1993, 22, 546–548. [Google Scholar]
- Riede, K. Acoustic monitoring of Orthoptera and its potential for conservation. J. Insect Conserv. 1998, 2, 217–223. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Country | Lat | Long | Elevation (m) | Precipitation (cm) | Forest Type | Date | Recordings | Sampling Days |
|---|---|---|---|---|---|---|---|---|---|
| Amarakaeri | Peru | −12.99E | −71.01S | 1000 | 6000 | Premontane | Feb. 2015 | 1897 | 13.2 |
| La Selva | Costa Rica | 10.43E | −83.98N | 37 | 4000 | Lowland wet | Jul. 2015 | 1043 | 7.2 |
| Las Cruces | Costa Rica | 8.78E | −82.95N | 1100 | 2500 | Premontane | Jul. 2015 | 1251 | 8.7 |
| Palo Verde | Costa Rica | 10.35E | −85.35N | 10 | 1500 | Lowland dry | Jun. 2015 | 616 | 4.3 |
| Cabo Blanco | Costa Rica | 9.57E | −85.10N | 30 | 1500 | Lowland dry | Jun. 2015 | 736 | 5.1 |
| Las Alturas | Costa Rica | 8.93E | −82.83N | 1800 | 2800 | Lower Montane | Jun. 2015 | 1251 | 8.7 |
| Cuerici | Costa Rica | 9.53E | −83.58N | 2600 | 3500 | Montane | Jul. 2015 | 1554 | 10.8 |
| El Verde | Puerto Rico | 18.32E | −65.82N | 363 | 3000 | Premontane | Jul. 2015 | 1696 | 11.8 |
| Site | % ASU | Regional Richness | Acoustic Morphospecies | |||
|---|---|---|---|---|---|---|
| Total | Anuran | Bird | Insect | |||
| Amarakaeri | 34.3 | 583 | 84 | 4 | 27 | 53 |
| La Selva | 20.3 | 423 | 56 | 3 | 20 | 33 |
| Las Cruces | 28.2 | 352 | 91 | 5 | 31 | 55 |
| Palo Verde | 14.9 | 330 | 38 | 0 | 21 | 17 |
| Cabo Blanco | 11.8 | 311 | 46 | 1 | 23 | 22 |
| Las Alturas | 18.5 | 262 | 50 | 3 | 26 | 21 |
| Cuerici | 13.3 | 190 | 17 | 0 | 14 | 3 |
| El Verde | 11.6 | 184 | 37 | 6 | 12 | 19 |
| Model Variables | Residual SS | Akaike Weight | Delta AICc | Adjusted R2 |
|---|---|---|---|---|
| Insects | 92.0 | 0.925 | 0 | 0.77 |
| Birds | 223.7 | 0.026 | 7.1 | 0.45 |
| Anurans | 389.6 | 0.003 | 11.5 | 0.04 |
| Anurans | Birds | Insects | |
|---|---|---|---|
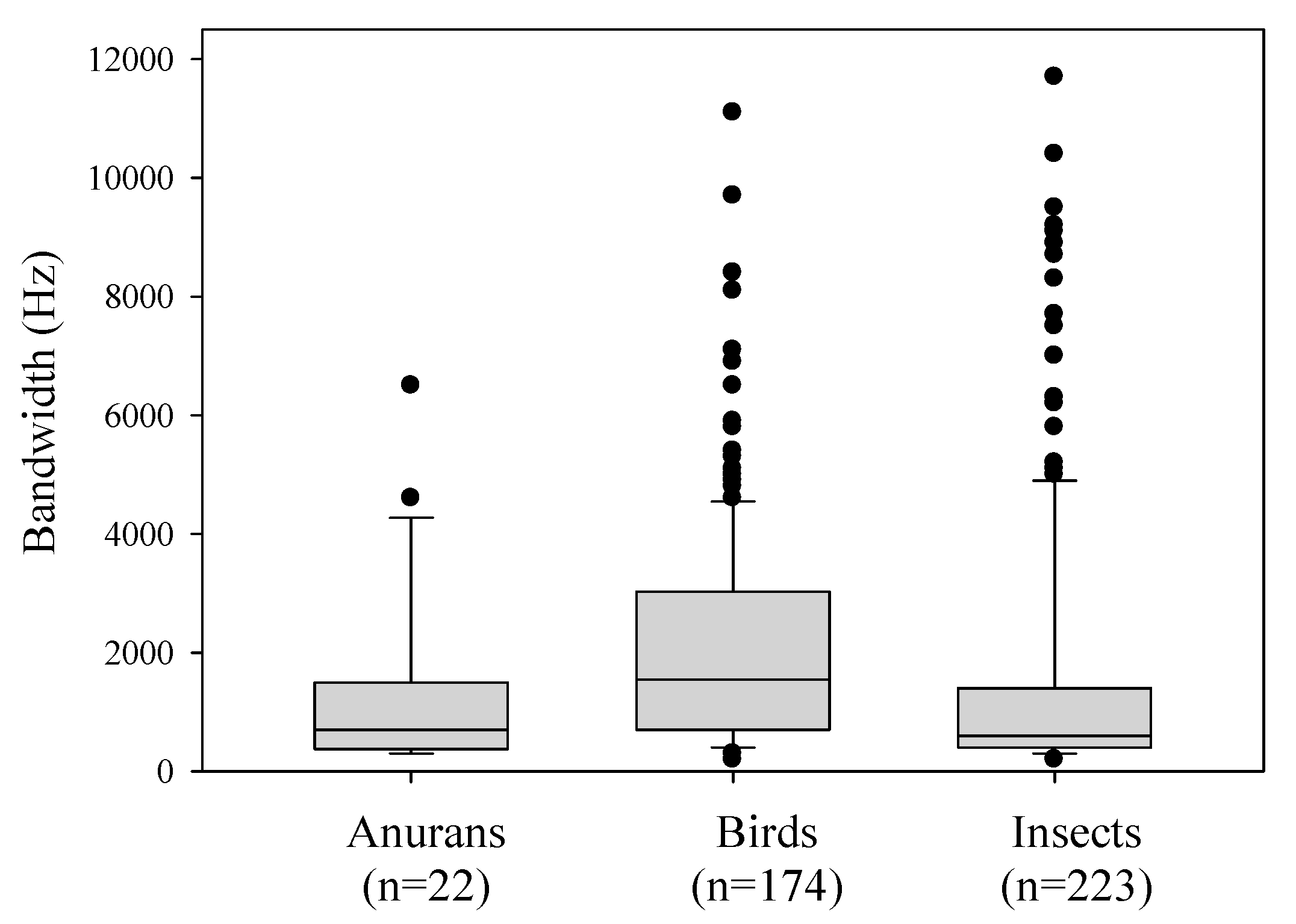
| Bandwidth (max–min) | + | +++ | ++ |
| Call/activity duration | ++ | + | +++ |
| Impact on ASU | Low | Medium | High |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aide, T.M.; Hernández-Serna, A.; Campos-Cerqueira, M.; Acevedo-Charry, O.; Deichmann, J.L. Species Richness (of Insects) Drives the Use of Acoustic Space in the Tropics. Remote Sens. 2017, 9, 1096. https://doi.org/10.3390/rs9111096
Aide TM, Hernández-Serna A, Campos-Cerqueira M, Acevedo-Charry O, Deichmann JL. Species Richness (of Insects) Drives the Use of Acoustic Space in the Tropics. Remote Sensing. 2017; 9(11):1096. https://doi.org/10.3390/rs9111096
Chicago/Turabian StyleAide, T. Mitchell, Andres Hernández-Serna, Marconi Campos-Cerqueira, Orlando Acevedo-Charry, and Jessica L. Deichmann. 2017. "Species Richness (of Insects) Drives the Use of Acoustic Space in the Tropics" Remote Sensing 9, no. 11: 1096. https://doi.org/10.3390/rs9111096
APA StyleAide, T. M., Hernández-Serna, A., Campos-Cerqueira, M., Acevedo-Charry, O., & Deichmann, J. L. (2017). Species Richness (of Insects) Drives the Use of Acoustic Space in the Tropics. Remote Sensing, 9(11), 1096. https://doi.org/10.3390/rs9111096