Determinants of Aboveground Biomass across an Afromontane Landscape Mosaic in Kenya

 ,
,  , , , and
, , , and 
Abstract
:
1. Introduction
2. Materials and Methods
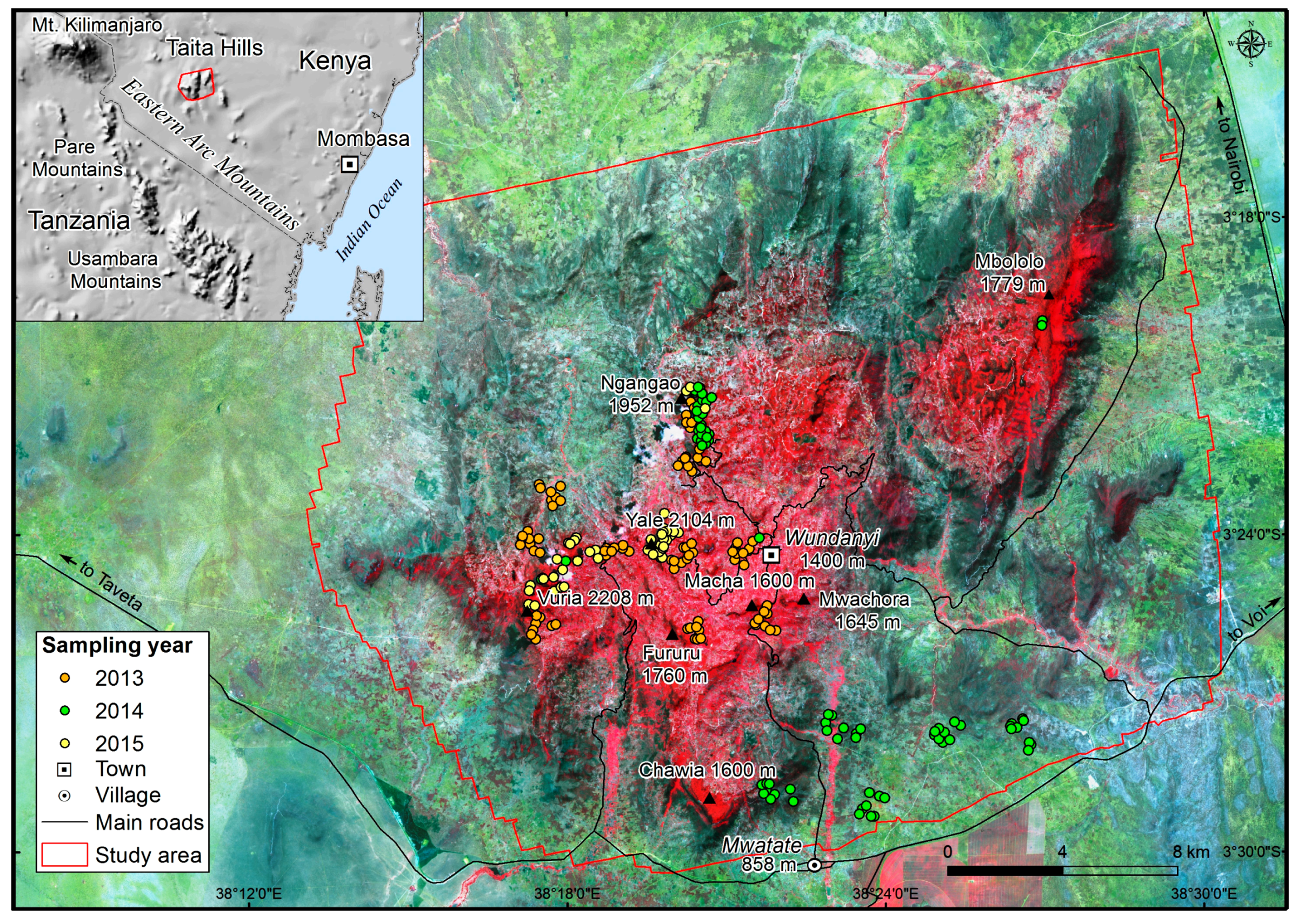
2.1. Study Area
2.2. Field Measurements and Aboveground Biomass Computations
2.3. Airborne Laser Scanning Data
2.4. Aboveground Biomass Mapping
2.5. Explanatory Variables for Modelling Distribution of Aboveground Biomass
2.6. Statistical Analysis
3. Results
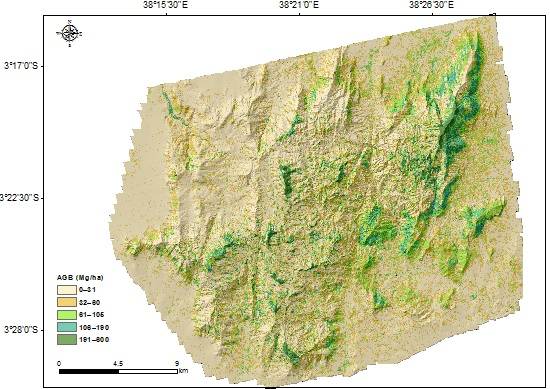
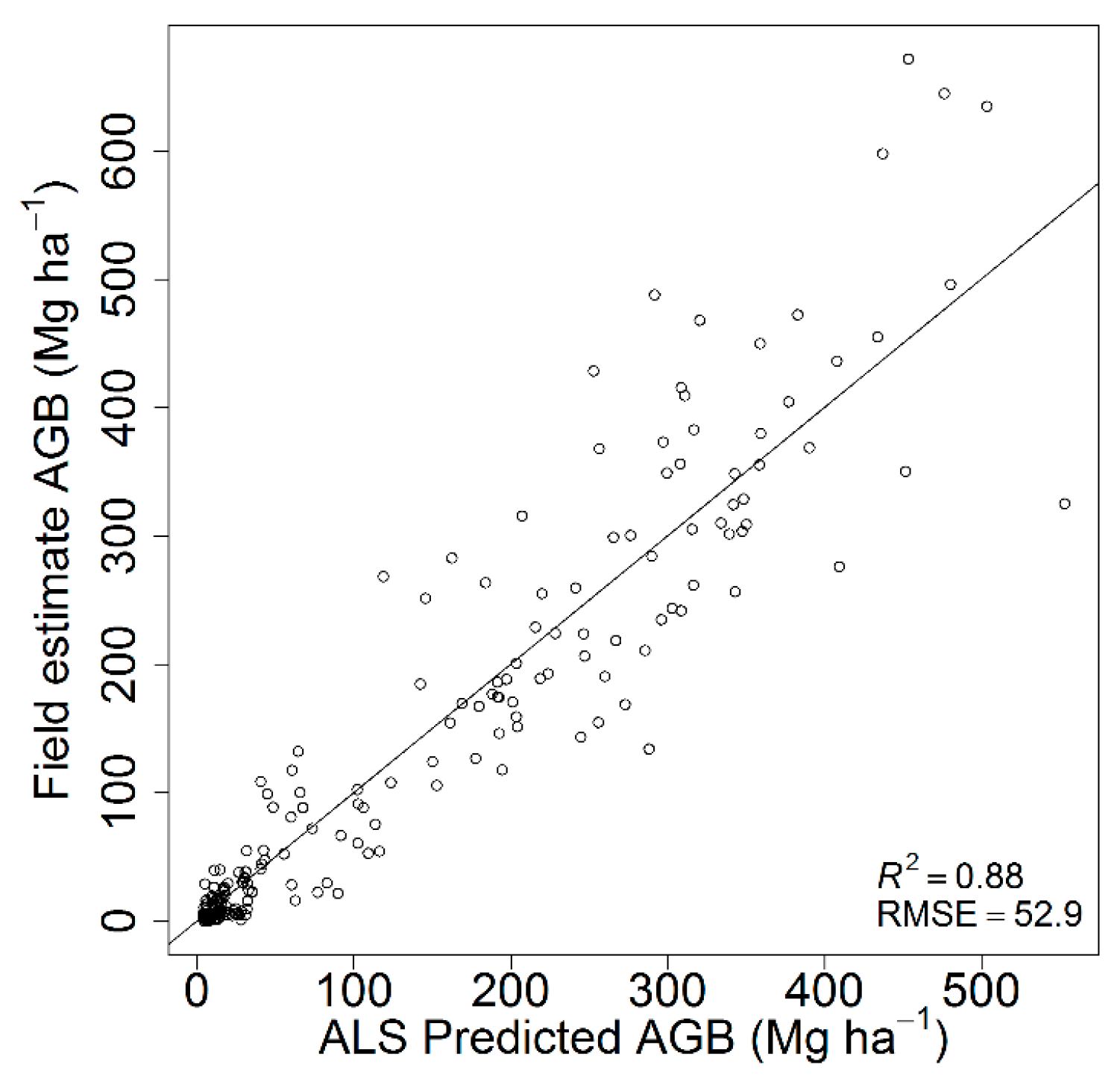
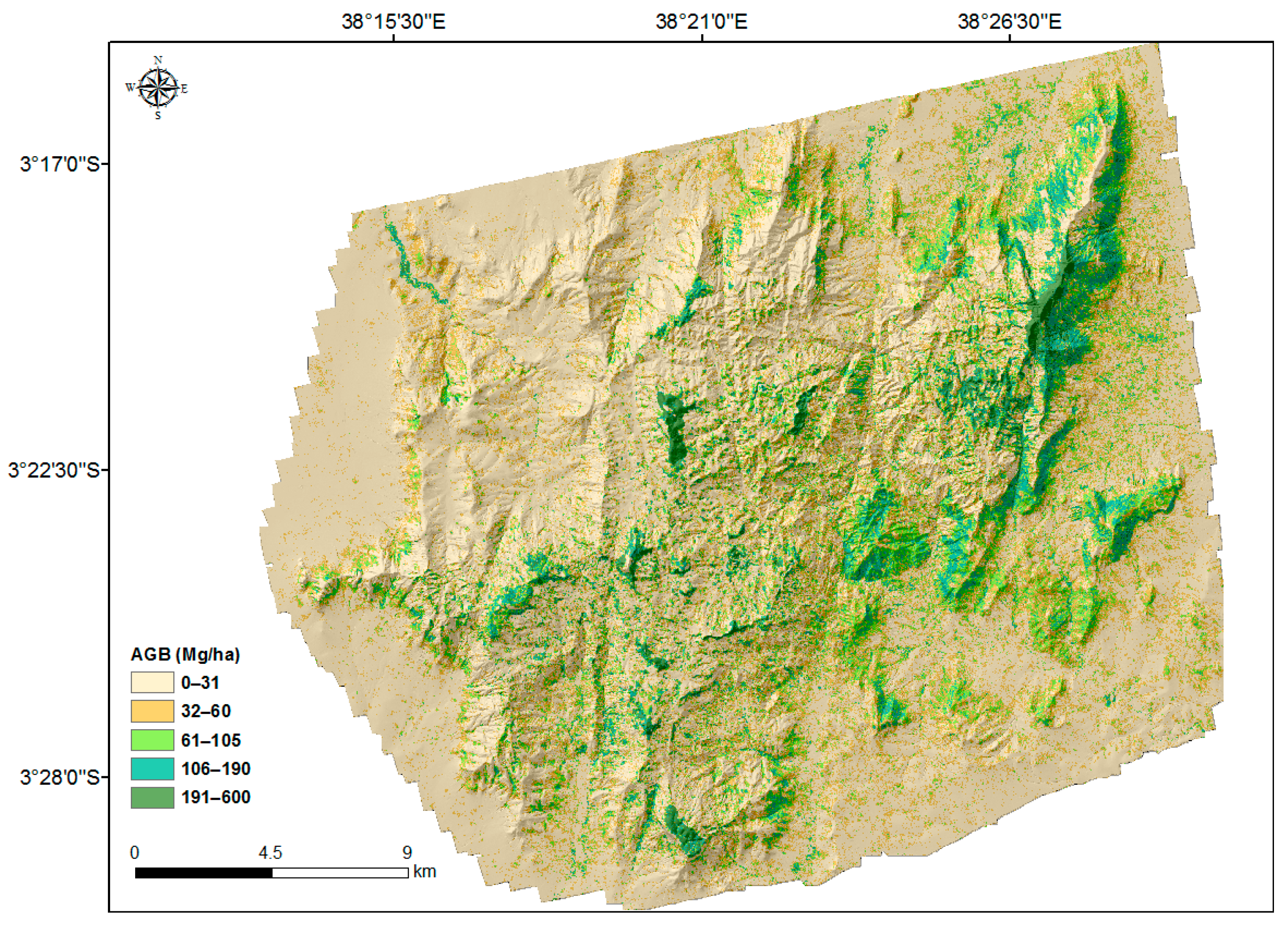
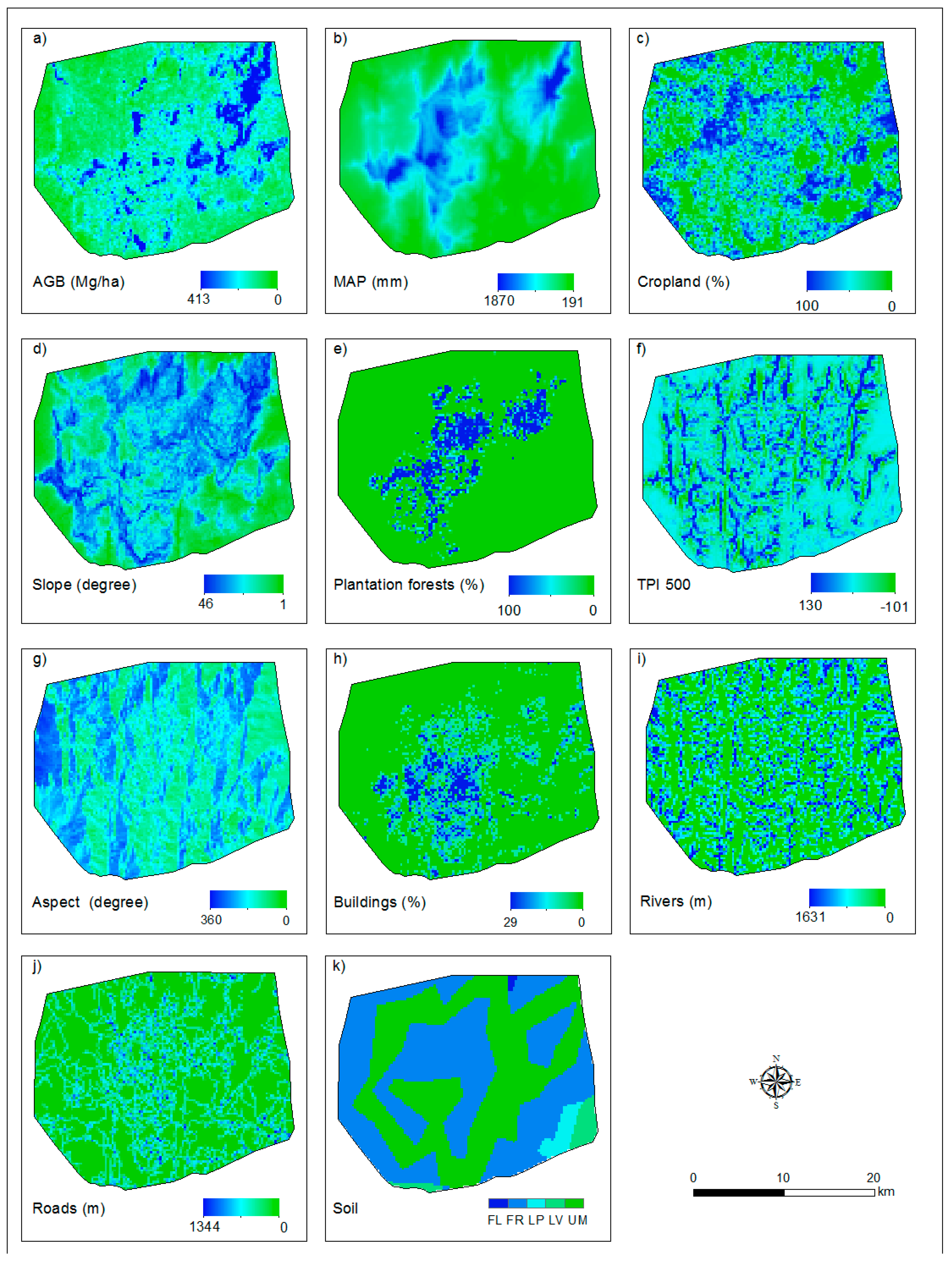
3.1. Aboveground Biomass Map
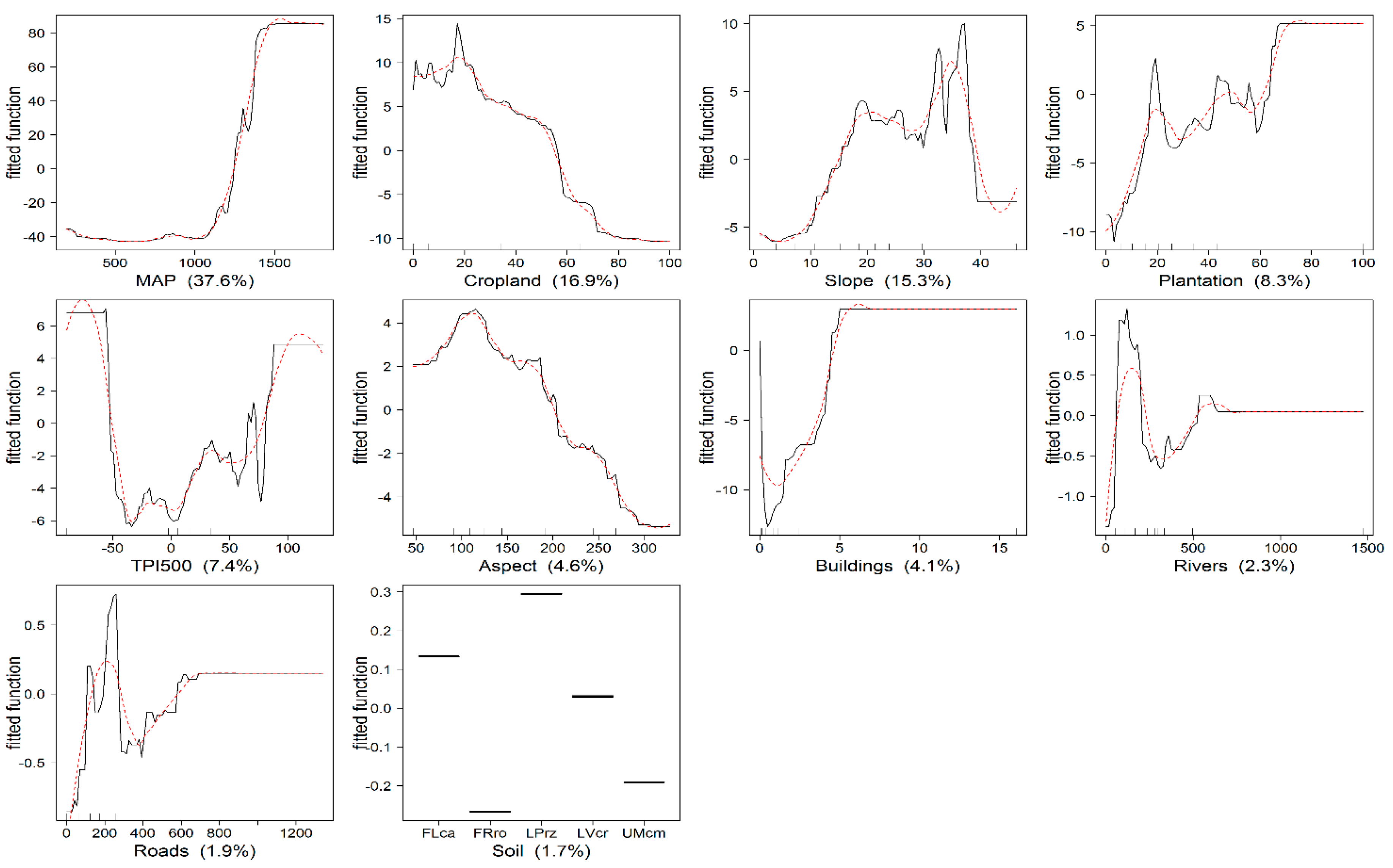
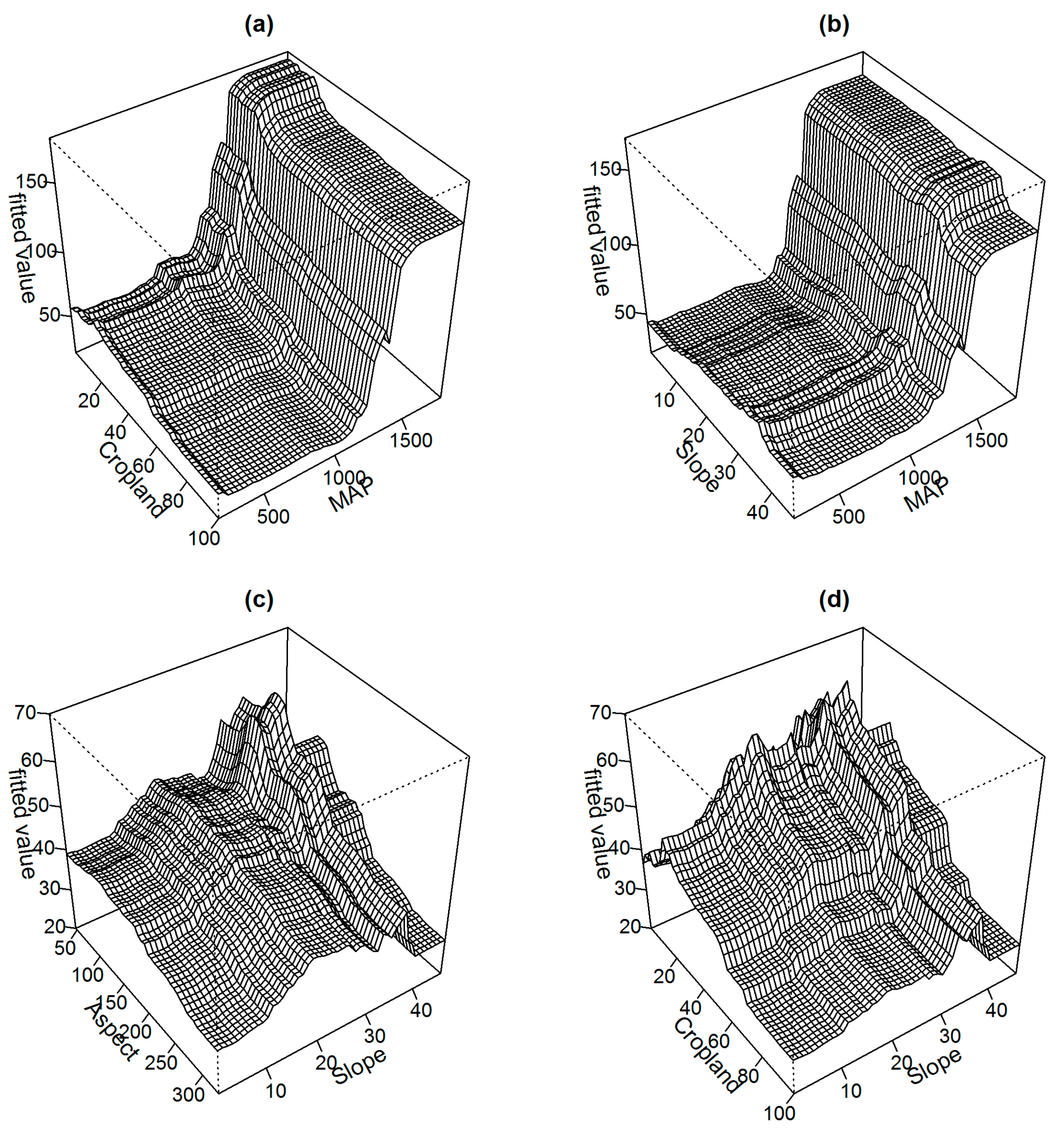
3.2. Determinants of Aboveground Biomass
4. Discussion
4.1. ALS-Based Aboveground Biomass Map
4.2. Determinants of Aboveground Biomass Distribution
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A



References
- Lovett, J.C.; Wasser, S.K. Biogeography and Ecology of the Rainforests of Eastern Africa; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Ioki, K.; Tsuyuki, S.; Hirata, Y.; Phua, M.-H.; Wong, W.V.C.; Ling, Z.-Y.; Saito, H.; Takao, G. Estimating above-ground biomass of tropical rainforest of different degradation levels in Northern Borneo using airborne LiDAR. Forest Ecol. Manag. 2014, 328, 335–341. [Google Scholar] [CrossRef]
- Pellikka, P.K.E.; Clark, B.J.F.; Gosa, A.G.; Himberg, N.; Hurskainen, P.; Maeda, E.; Mwang’ombe, J.; Omoro, L.M.A.; Siljander, M. Chapter 13—Agricultural expansion and its consequences in the Taita Hills, Kenya. In Developments in Earth Surface Processes; Paolo Paron, D.O.O., Christian Thine, O., Eds.; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Pfeifer, M.; Platts, P.J.; Burgess, N.D.; Swetnam, R.D.; Willcock, S.; Lewis, S.L.; Marchant, R. Land use change and carbon fluxes in East Africa quantified using earth observation data and field measurements. Environ. Conserv. 2012, 40, 241–252. [Google Scholar] [CrossRef]
- United Nations Framework Convention on Climate Change. Available online: http://unfccc.int/meetings/bali_dec_2007/meeting/6319.php (accessed on 11 August 2017).
- Asner, G.P.; Clark, J.K.; Mascaro, J.; Vaudry, R.; Chadwick, K.D.; Vieilledent, G.; Rasamoelina, M.; Balaji, A.; Kennedy-Bowdoin, T.; Maatoug, L.; et al. Human and environmental controls over aboveground carbon storage in Madagascar. Carbon Balance Manag. 2012, 7, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, D. The potential and challenge of remote sensing-based biomass estimation. Int. J. Remote Sens. 2006, 27, 1297–1328. [Google Scholar] [CrossRef]
- Zolkos, S.G.; Goetz, S.J.; Dubayah, R. A meta-analysis of terrestrial aboveground biomass estimation using LiDAR remote sensing. Remote Sens. Environ. 2013, 128, 289–298. [Google Scholar] [CrossRef]
- Singh, K.K.; Chen, G.; McCarter, J.B.; Meentemeyer, R.K. Effects of LiDAR point density and landscape context on estimates of urban forest biomass. ISPRS J. Photogramm. Remote Sens. 2015, 101, 310–322. [Google Scholar] [CrossRef]
- Vaglio Laurin, G.; Chen, Q.; Lindsell, J.A.; Coomes, D.A.; Frate, F.D.; Guerriero, L.; Pirotti, F.; Valentini, R. Above ground biomass estimation in an African tropical forest with LiDAR and hyperspectral data. ISPRS J. Photogramm. Remote Sens. 2014, 89, 49–58. [Google Scholar] [CrossRef]
- Hansen, E.H.; Gobakken, T.; Bollandsas, O.M.; Zahabu, E.; Naesset, E. Modeling aboveground biomass in dense tropical submontane rainforest using airborne laser scanner data. Remote Sens. 2015, 7, 788–807. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Vaglio Laurin, G.; Valentini, R. Uncertainty of remotely sensed aboveground biomass over an African tropical forest: Propagating errors from trees to plots to pixels. Remote Sens. Environ. 2015, 160, 134–143. [Google Scholar] [CrossRef]
- Vaglio Laurin, G.; Puletti, N.; Chen, Q.; Corona, P.; Papale, D.; Valentini, R. Above ground biomass and tree species richness estimation with airborne LiDAR in tropical Ghana forests. Int. J. Appl. Earth Obs. 2016, 52, 371–379. [Google Scholar] [CrossRef]
- Zomer, R.J.; Neufeldt, H.; Xu, J.; Ahrends, A.; Bossio, D.; Trabucco, A.; van Noordwijk, M.; Wang, M. Global tree cover and biomass carbon on agricultural land: The contribution of agroforestry to global and national carbon budgets. Sci. Rep. 2016, 6, 29987. [Google Scholar] [CrossRef] [PubMed]
- Pellikka, P.K.E.; Heikinheimo, V.; Hietanen, J.; Schäfer, E.; Siljander, M.; Heiskanen, J. Assessment of the impact of land cover change on aboveground carbon stocks using airborne LiDAR data and satellite imagery in Afromontane landscape in Kenya. 2017; Submitted. [Google Scholar]
- Asner, G.P.; Hughes, R.F.; Varga, T.A.; Knapp, D.E.; Kennedy-Bowdoin, T. Environmental and biotic controls over aboveground biomass throughout a tropical rain forest. Ecosystems 2009, 12, 261–278. [Google Scholar] [CrossRef]
- Singh, K.K.; Bianchetti, R.A.; Chen, G.; Meentemeyer, R.K. Assessing effect of dominant land-cover types and pattern on urban forest biomass estimated using LiDAR metrics. Urban Ecosyst. 2017, 20, 265–275. [Google Scholar] [CrossRef]
- Clark, D.B.; Clark, D.A. Landscape-scale variation in forest structure and biomass in a tropical rain forest. Forest Ecol. Manag. 2000, 137, 185–198. [Google Scholar] [CrossRef]
- Taylor, P.; Asner, G.; Dahlin, K.; Anderson, C.; Knapp, D.; Martin, R.; Mascaro, J.; Chazdon, R.; Cole, R.; Wanek, W.; et al. Landscape-scale controls on aboveground forest carbon stocks on the Osa Peninsula, Costa Rica. PLoS ONE 2015, 10, e0126748. [Google Scholar] [CrossRef] [PubMed]
- Marshall, A.R.; Willcock, S.; Platts, P.J.; Lovett, J.C.; Balmford, A.; Burgess, N.D.; Latham, J.E.; Munishi, P.K.T.; Salter, R.; Shirima, D.D.; et al. Measuring and modelling above-ground carbon and tree allometry along a tropical elevation gradient. Biol. Conserv. 2012, 154, 20–33. [Google Scholar] [CrossRef]
- Pfeifer, M.; Gonsamo, A.; Disney, M.; Pellikka, P.; Marchant, R. Leaf area index for biomes of the Eastern Arc Mountains: Landsat and spot observations along precipitation and altitude gradients. Remote Sens. Environ. 2012, 118, 103–115. [Google Scholar] [CrossRef]
- Commission on Revenue Allocation. Available online: http://www.crakenya.org/ (accessed on 11 August 2017).
- Erdogan, H.E.; Pellikka, P.K.E.; Clark, B. Modelling the impact of land-cover change on potential soil loss in the Taita Hills, Kenya, between 1987 and 2003 using remote-sensing and geospatial data. Int. J. Remote Sens. 2011, 32, 5919–5945. [Google Scholar] [CrossRef]
- Pellikka, P.K.E.; Lötjönen, M.; Siljander, M.; Lens, L. Airborne remote sensing of spatiotemporal change (1955–2004) in indigenous and exotic forest cover in the Taita Hills, Kenya. Int. J. Appl. Earth Obs. 2009, 11, 221–232. [Google Scholar] [CrossRef]
- Aerts, R.; Thijs, K.W.; Lehouck, V.; Beentje, H.; Bytebier, B.; Matthysen, E.; Gulinck, H.; Lens, L.; Muys, B. Woody plant communities of isolated afromontane cloud forests in Taita Hills, Kenya. Plant Ecol. 2011, 212, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Stam, Å.; Enroth, J.; Malombe, I.; Pellikka, P.; Rikkinen, J. Experimental transplants reveal strong environmental effects on the growth on non-vascular epiphytes in Afromontane forests. Biotropica 2017. in print. [Google Scholar] [CrossRef]
- Enroth, J.; Nyqvist, P.; Malombe, I.; Pellikka, P.; Rikkinen, J. Additions to the moss flora of the Taita Hills and mount Kasigau, Kenya. Polish Bot. J. 2013, 58, 495. [Google Scholar] [CrossRef]
- Thijs, K.W.; Aerts, R.; Musila, W.; Siljander, M.; Matthysen, E.; Lens, L.; Pellikka, P.; Gulinck, H.; Muys, B. Potential tree species extinction, colonization and recruitment in Afromontane forest relicts. Basic Appl. Ecol. 2014, 15, 288–296. [Google Scholar] [CrossRef] [Green Version]
- Thijs, K.W.; Aerts, R.; van de Moortele, P.; Aben, J.; Musila, W.; Pellikka, P.; Gulinck, H.; Muys, B. Trees in a human-modified tropical landscape: Species and trait composition and potential ecosystem services. Landscape Urban. Plan. 2015, 144, 49–58. [Google Scholar] [CrossRef]
- European Spatial Agency. Sentinel-2 User Handbook. ESA Standard Document. Available online: https://sentinel.esa.int/documents/247904/685211/Sentinel-2_User_Handbook (accessed on 24 July 2015).
- Heiskanen, J.; Korhonen, L.; Hietanen, J.; Pellikka, P.K.E. Use of airborne LiDAR for estimating canopy gap fraction and leaf area index of tropical montane forests. Int. J. Remote Sens. 2015, 36, 2569–2583. [Google Scholar] [CrossRef]
- Piiroinen, R.; Heiskanen, J.; Mõttus, M.; Pellikka, P. Classification of crops across heterogeneous agricultural landscape in Kenya using Aisaeagle imaging spectroscopy data. Int. J. Appl. Earth Obs. 2015, 39, 1–8. [Google Scholar] [CrossRef]
- Curtis, R.O. Height-diameter and height-diameter-age equations for second-growth douglas-fir. Forest Sci. 1967, 13, 365–375. [Google Scholar]
- Valbuena, R.; Heiskanen, J.; Aynekulu, E.; Pitkänen, S.; Packalen, P. Sensitivity of above-ground biomass estimates to height-diameter modelling in mixed-species West African woodlands. PLoS ONE 2016, 11, e0158198. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1–118. Available online: http://CRAN.R-project.org/package=nlme (accessed on 6 February 2017).
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: http://www.gbif.org/resource/81287 (accessed on 11 August 2017).
- Henry, M.; Picard, N.; Trotta, C.; Manlay, R.; Valentini, R.; Bernoux, M.; Saint-André, L. Estimating tree biomass of sub-Saharan African forests: A review of available allometric equations. Silva Fenn. 2011, 45, 477–569. [Google Scholar] [CrossRef]
- Paul, K.I.; Roxburgh, S.H.; England, J.R.; Ritson, P.; Hobbs, T.; Brooksbank, K.; John Raison, R.; Larmour, J.S.; Murphy, S.; Norris, J.; et al. Development and testing of allometric equations for estimating above-ground biomass of mixed-species environmental plantings. Forest Ecol. Manag. 2013, 310, 483–494. [Google Scholar] [CrossRef]
- Frangi, J.L.; Lugo, A.E. Ecosystem dynamics of a subtropical floodplain forest. Ecol. Monogr. 1985, 55, 351–369. [Google Scholar] [CrossRef]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.C.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved allometric models to estimate the aboveground biomass of tropical trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef] [PubMed]
- ICRAF. World Agroforestry Center Wood Density Database. Available online: https://www.worldagroforestry.org/output/wood-density-database (accessed on 2 February 2016).
- Data from: Towards A Worldwide Wood Economics Spectrum. Available online: http://datadryad.org/resource/doi:10.5061/dryad.234 (accessed on 4 February 2009).
- Cao, L.; Coops, N.C.; Innes, J.L.; Sheppard, S.R.J.; Fu, L.; Ruan, H.; She, G. Estimation of forest biomass dynamics in subtropical forests using multi-temporal airborne LiDAR data. Remote Sens. Environ. 2016, 178, 158–171. [Google Scholar] [CrossRef]
- McGaughey, R.J. Fusion/LDV: Software for LiDAR Data Analysis and Visualization; US Department of Agriculture, Forest Service, Pacific Northwest Research Station: Seattle, WA, USA, 2016.
- Asner, G.P.; Mascaro, J. Mapping tropical forest carbon: Calibrating plot estimates to a simple LiDAR metric. Remote Sens. Environ. 2014, 140, 614–624. [Google Scholar] [CrossRef]
- Bouvier, M.; Durrieu, S.; Fournier, R.A.; Renaud, J.-P. Generalizing predictive models of forest inventory attributes using an area-based approach with airborne LiDAR data. Remote Sens. Environ. 2015, 156, 322–334. [Google Scholar] [CrossRef]
- Package ‘Leaps’. Available online: https://cran.r-project.org/web/packages/leaps/leaps.pdf (accessed on 10 January 2017).
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Weiss, A. Topographic Position and Landforms Analysis. Available online: http://www.jennessent.com/downloads/TPI-poster-TNC_18x22.pdf (accessed on 11 August 2017).
- Guisan, A.; Weiss, S.B.; Weiss, A.D. GLM versus CCA spatial modeling of plant species distribution. Plant Ecol. 1999, 143, 107–122. [Google Scholar] [CrossRef]
- Böhner, J.; Köthe, R.; Conrad, O.; Gross, J.; Ringeler, A.; Selige, T. Soil regionalisation by means of terrain analysis and process parameterisation. In Soil Classification; The European Soil Bureau, Joint Research Centre: Ispra, Italy, 2002. [Google Scholar]
- Costa-Cabral, M.C.; Burges, S.J. Digital elevation model networks (DEMON): A model of flow over hillslopes for computation of contributing and dispersal areas. Water Resour. Res. 1994, 30, 1681–1692. [Google Scholar] [CrossRef]
- Soil Atlas of Africa and Its Associated Soil Map (Data). Available online: http://esdac.jrc.ec.europa.eu/content/soil-map-soil-atlas-africa (accessed on 21 June 2017).
- Virtanen, E. Fine-Resolution Climate Grids for Species Studies in Data-Poor Regions. Available online: https://helda.helsinki.fi/handle/10138/156514 (accessed on 11 September 2015).
- Hutchinson, M.F. The application of thin plate splines to continent-wide data assimilation. BMRC Res. Rep. 1991, 27, 104–113. [Google Scholar]
- Heikinheimo, V. Impact of Land Change on Aboveground Carbon Stocks in the Taita Hills, Kenya. Master’s Thesis, University of Helsinki, Helsinki, Finland, 25 May 2015. [Google Scholar]
- ESRI. Redlands, CA: 1999–2015. Available online: http://www.esri.com/data/basemaps (accessed on 21 June 2017).
- Elith, J.; Leathwick, J.R.; Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Riihimäki, H.; Heiskanen, J.; Luoto, M. The effect of topography on arctic-alpine aboveground biomass and ndvi patterns. Int. J. Appl. Earth Obs. 2017, 56, 44–53. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Phillips, S.J.; Leathwick, J.; Elith, J. Package ‘dismo’. Available online: http://www12.frugalware.org/mirrors/cran.r-project.org/web/packages/dismo/dismo.pdf (accessed on 9 January 2017).
- Gregoire, T.G.; Lin, Q.F.; Boudreau, J.; Nelson, R. Regression estimation following the square root transformation of the response. Forest Sci. 2008, 54, 597–606. [Google Scholar]
- Mascaro, J.; Detto, M.; Asner, G.P.; Muller-Landau, H.C. Evaluating uncertainty in mapping forest carbon with airborne LiDAR. Remote Sens. Environ. 2011, 115, 3770–3774. [Google Scholar] [CrossRef]
- Chave, J.; Condit, R.; Muller-Landau, H.C.; Thomas, S.C.; Ashton, P.S.; Bunyavejchewin, S.; Co, L.L.; Dattaraja, H.S.; Davies, S.J.; Esufali, S.; et al. Assessing evidence for a pervasive alteration in tropical tree communities. PLoS Biol. 2008, 6, e45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keränen, J.; Peuhkurinen, J.; Packalen, P.; Maltamo, M. Effect of minimum diameter at breast height and standing dead wood field measurements on the accuracy of als-based forest inventory. Can. J. Forest Res. 2015, 45, 1280–1288. [Google Scholar] [CrossRef]
- Breidenbach, J.; Koch, B.; Kändler, G.; Kleusberg, A. Quantifying the influence of slope, aspect, crown shape and stem density on the estimation of tree height at plot level using LiDAR and insar data. Int. J. Remote Sens. 2008, 29, 1511–1536. [Google Scholar] [CrossRef]
- Duan, Z.; Zhao, D.; Zeng, Y.; Zhao, Y.; Wu, B.; Zhu, J. Assessing and correcting topographic effects on forest canopy height retrieval using airborne LiDAR data. Sensors 2015, 15, 12133. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.L.; Sonké, B.; Sunderland, T.; Begne, S.K.; Lopez-Gonzalez, G.; van der Heijden, G.M.F.; Phillips, O.L.; Affum-Baffoe, K.; Baker, T.R.; Banin, L.; et al. Above-ground biomass and structure of 260 African tropical forests. Philos. Trans. R. Soc. B 2013, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruijnzeel, L.A.; Veneklaas, E.J. Climatic conditions and tropical montane forest productivity: The fog has not lifted yet. Ecology 1998, 79, 3–9. [Google Scholar] [CrossRef]
- McEwan, R.W.; Lin, Y.-C.; Sun, I.F.; Hsieh, C.-F.; Su, S.-H.; Chang, L.-W.; Song, G.-Z.M.; Wang, H.-H.; Hwong, J.-L.; Lin, K.-C.; et al. Topographic and biotic regulation of aboveground carbon storage in subtropical broad-leaved forests of Taiwan. Forest Ecol. Manag. 2011, 262, 1817–1825. [Google Scholar] [CrossRef]
- Mensah, S.; Veldtman, R.; Assogbadjo, A.E.; Glèlè Kakaï, R.; Seifert, T. Tree species diversity promotes aboveground carbon storage through functional diversity and functional dominance. Ecol. Evol. 2016, 6, 7546–7557. [Google Scholar] [CrossRef] [PubMed]
- Himberg, N.; Omoro, L.; Pellikka, P.; Luukkanen, O. The benefits and constraints of participation in forest management. The case of Taita Hills, Kenya. Fennia Int. J. Geogr. 2009, 187, 61–76. [Google Scholar]
- Himberg, N. Traditionally Protected Forests´ Role within Transforming Natural Resource Management Regimes in Taita Hills, Kenya. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 5 December 2011. [Google Scholar]
- Holl, K.D.; Zahawi, R.A. Factors explaining variability in woody above-ground biomass accumulation in restored tropical forest. Forest Ecol. Manag. 2014, 319, 36–43. [Google Scholar] [CrossRef]
- Siljander, M.; Clark, B.J.F.; Pellikka, P.K.E. A predictive modelling technique for human population distribution and abundance estimation using remote-sensing and geospatial data in a rural mountainous area in kenya. Int J. Remote Sens. 2011, 32, 5997–6023. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Attribute | Range | Median | Mean | SD |
|---|---|---|---|---|
| Density (stems/ha) | 10–1214 | 160 | 309 | 301 |
| Basal area (m2) | 0.1–94.3 | 9.2 | 19.9 | 21.6 |
| Mean DBH (cm) | 10.4–46.1 | 22.4 | 23.5 | 7.2 |
| Lorey’s mean height (m) | 3.0–39.3 | 12.6 | 13.8 | 7.0 |
| AGB (Mg/ha) | 0.1–671.3 | 37.9 | 123.0 | 153.0 |
| Parameter | Value |
|---|---|
| Dates of acquisition | 2014 (26 January, 6 February and 8 February) and 2015 (5, 6, 11 and 13 February) |
| Sensor | Leica ALS60 |
| Mean range (m) | 1460 |
| Pulse rate (kHz) | 58 |
| Scan rate (Hz) | 66 |
| Scan angle (°) | ±16 |
| Mean Pulse density (pulses m−2) | 3.1 |
| Range of Pulse density (pulses m−2) | 1.0–4.9 |
| Mean return density (returns m−2) | 3.4 |
| Beam divergence at 1/e2 (mrad) | 0.22 |
| Mean footprint diameter (cm) | 32 |
| Metric | Description |
|---|---|
| H.p01, H.p05, H.p10, H.p20, H.p25, H.p30, H.p40, H.p50, H.p60, H.p70, H.p75, H.p80, H.p90, H.p95, H.p99 | 1st, 5th, 10th … and 99th percentile of return heights >3 m |
| H.max | Maximum of return heights >3 m |
| H.mean | Mean of return heights >3 m |
| H.cv | Coefficient of variation of return heights >3 m |
| H.stdev | Standard deviation of return heights >3 m |
| H.skewness | Skewness of return heights >3 m |
| H.kurtosis | Kurtosis of return heights >3 m |
| CC.first | First returns >3 m/total first returns × 100 |
| CC.all | All returns >3 m/total all returns × 100 |
| CC.all.first | All returns >3 m/total first returns × 100 |
| CC.first.mean | First returns above mean/total first returns × 100 |
| CC.all.mean | All returns above mean/total all returns × 100 |
| CC.all.mean.first | All returns above mean/total first returns × 100 |
| Variable | Description | Resolution (m) |
|---|---|---|
| Topography, hydrology and soil | ||
| Elevation | Elevation (m a.s.l.) based on DEM | 5 |
| Slope | Slope (°) based on DEM | 5 |
| Aspect | Aspect (°) based on DEM | 5 |
| TPI50–TPI500 | Topographic position index (TPI) with 50 m, 100 m, 150 m, 200 m, 300 m and 500 m radii based on DEM | 5 |
| TWI | Topographic wetness index based on DEM | 5 |
| Rivers | Length of rivers (m); river network extracted from DEM | 10 |
| Soil | Soil type vector layer from the Soil Atlas of Africa | - |
| Climate | ||
| MAT | Mean annual temperature (°C) | 20 |
| MAP | Mean annual precipitation (mm) | 20 |
| Land use | ||
| Cropland | Cropland cover (%) based on the LULC map | 20 |
| Plantation | Plantation forest cover (%) based on the LULC map | 20 |
| Building | Cover (%) of buildings extracted from ALS point cloud | 2 |
| Road | Length (m) of road digitized from the high-resolution imagery | - |
| Dependent Variable | Explanatory Variables | Estimate | SE of Estimate |
|---|---|---|---|
| Intercept | 0.423 *** | 0.268 | |
| H.p25 | 0.372 *** | 0.033 | |
| CC.all.first | 0.086 *** | 0.005 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adhikari, H.; Heiskanen, J.; Siljander, M.; Maeda, E.; Heikinheimo, V.; K. E. Pellikka, P. Determinants of Aboveground Biomass across an Afromontane Landscape Mosaic in Kenya. Remote Sens. 2017, 9, 827. https://doi.org/10.3390/rs9080827
Adhikari H, Heiskanen J, Siljander M, Maeda E, Heikinheimo V, K. E. Pellikka P. Determinants of Aboveground Biomass across an Afromontane Landscape Mosaic in Kenya. Remote Sensing. 2017; 9(8):827. https://doi.org/10.3390/rs9080827
Chicago/Turabian StyleAdhikari, Hari, Janne Heiskanen, Mika Siljander, Eduardo Maeda, Vuokko Heikinheimo, and Petri K. E. Pellikka. 2017. "Determinants of Aboveground Biomass across an Afromontane Landscape Mosaic in Kenya" Remote Sensing 9, no. 8: 827. https://doi.org/10.3390/rs9080827
APA StyleAdhikari, H., Heiskanen, J., Siljander, M., Maeda, E., Heikinheimo, V., & K. E. Pellikka, P. (2017). Determinants of Aboveground Biomass across an Afromontane Landscape Mosaic in Kenya. Remote Sensing, 9(8), 827. https://doi.org/10.3390/rs9080827