Heat and Pressure Treatments on Almond Protein Stability and Change in Immunoreactivity after Simulated Human Digestion




Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Autoclave Processing
- (a)
- Almond autoclaved for 10 min (AC10),
- (b)
- Almond autoclaved for 20 min (AC20),
- (c)
- Almond prehydrated + autoclaved for 10 min (H2O_AC10),
- (d)
- Almond prehydrated + autoclaved for 20 min (H2O_AC20).
2.3. Protein Extraction and Quantification
2.4. Sandwich ELISA for Almond Immunoreactivity
2.5. Almond In Vitro Digestion
- (a)
- untreated almonds submitted only to biological fluids, no enzymes (CTRL-NE),
- (b)
- untreated almonds undergoing the whole GI digestion (CTRL-GI),
- (c)
- AC10 almonds subjected to complete GI digestion (AC10-GI),
- (d)
- H2O-AC10 almonds subjected to the whole GI digestion (H2O-AC10-GI).
2.6. Electrophoretic Analysis of Almond Proteins
2.7. In-Gel Tryptic Digestion
2.8. Separation of Low Molecular Weight Fractions of Duodenal Samples
2.9. Untargeted HPLC-HRMS/MS Analysis
2.10. Bioinformatics Analysis for Assessing the Residual Immunoreactivity of Almond Proteins after GI Digestion
3. Results and Discussion
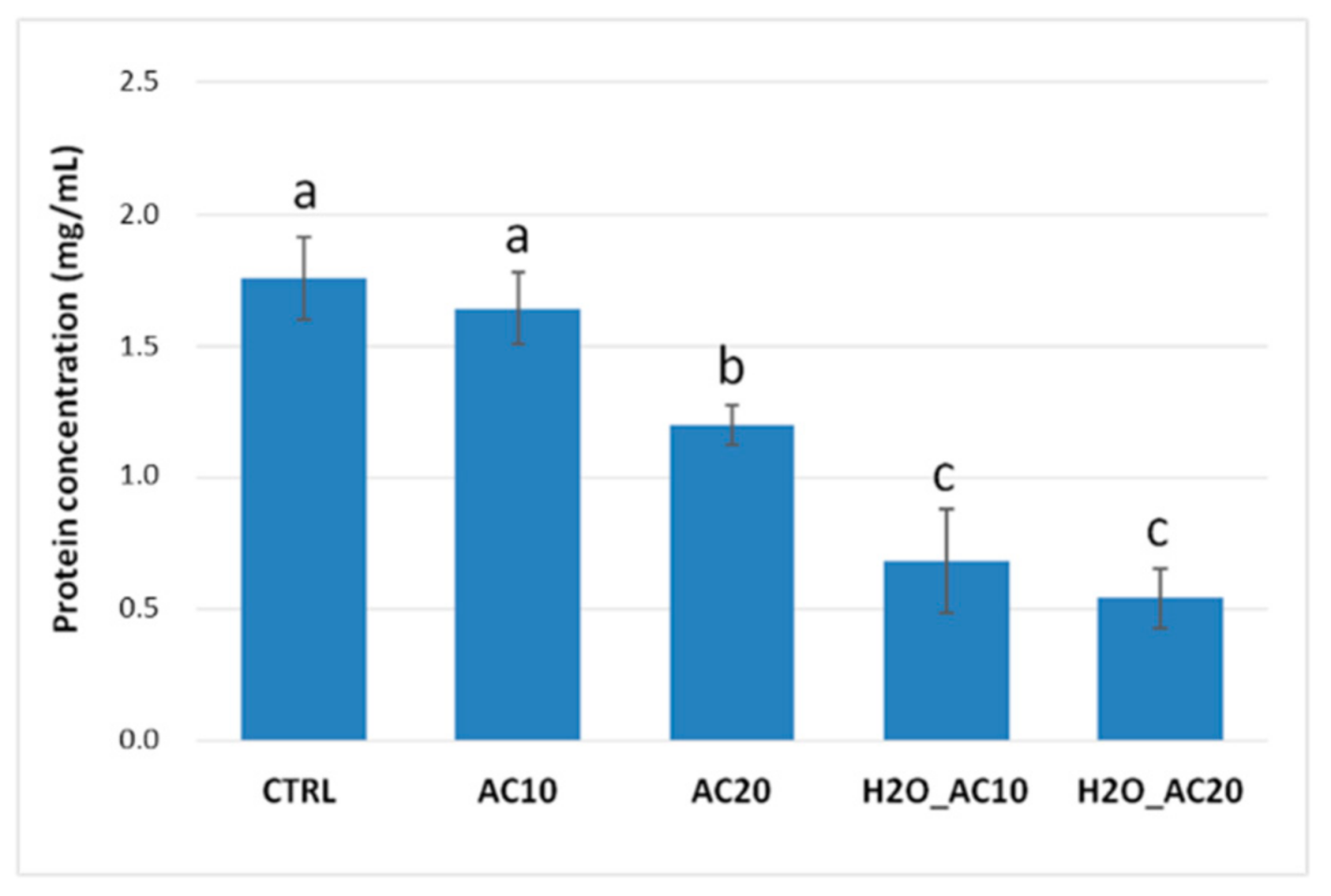
3.1. Effect of Thermal/Pressure Treatments on Solubility/Content of Almond Proteins
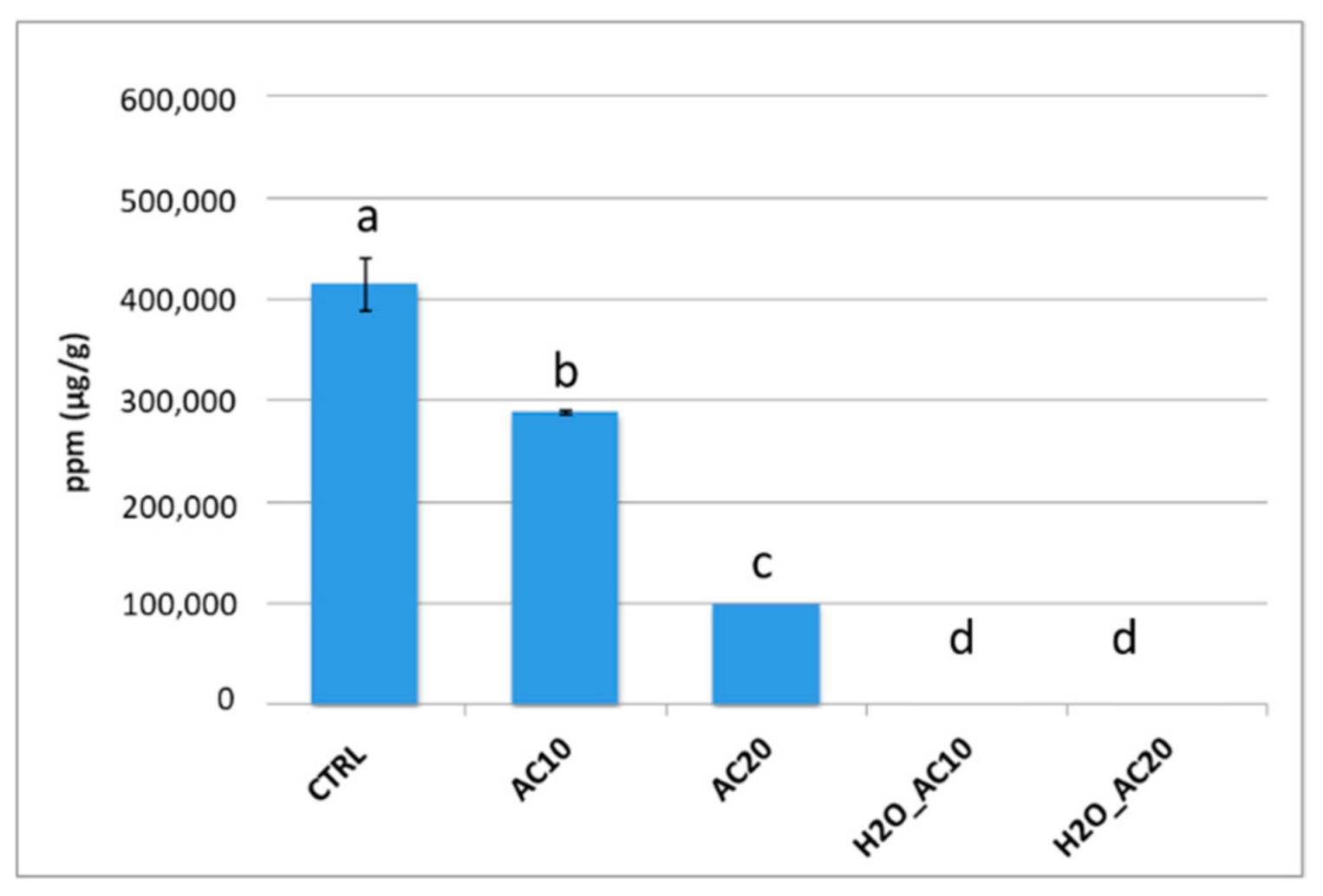
3.2. Impact of Thermal/Pressure Process on Immunoreactivity of Almond Proteins by ELISA Assay
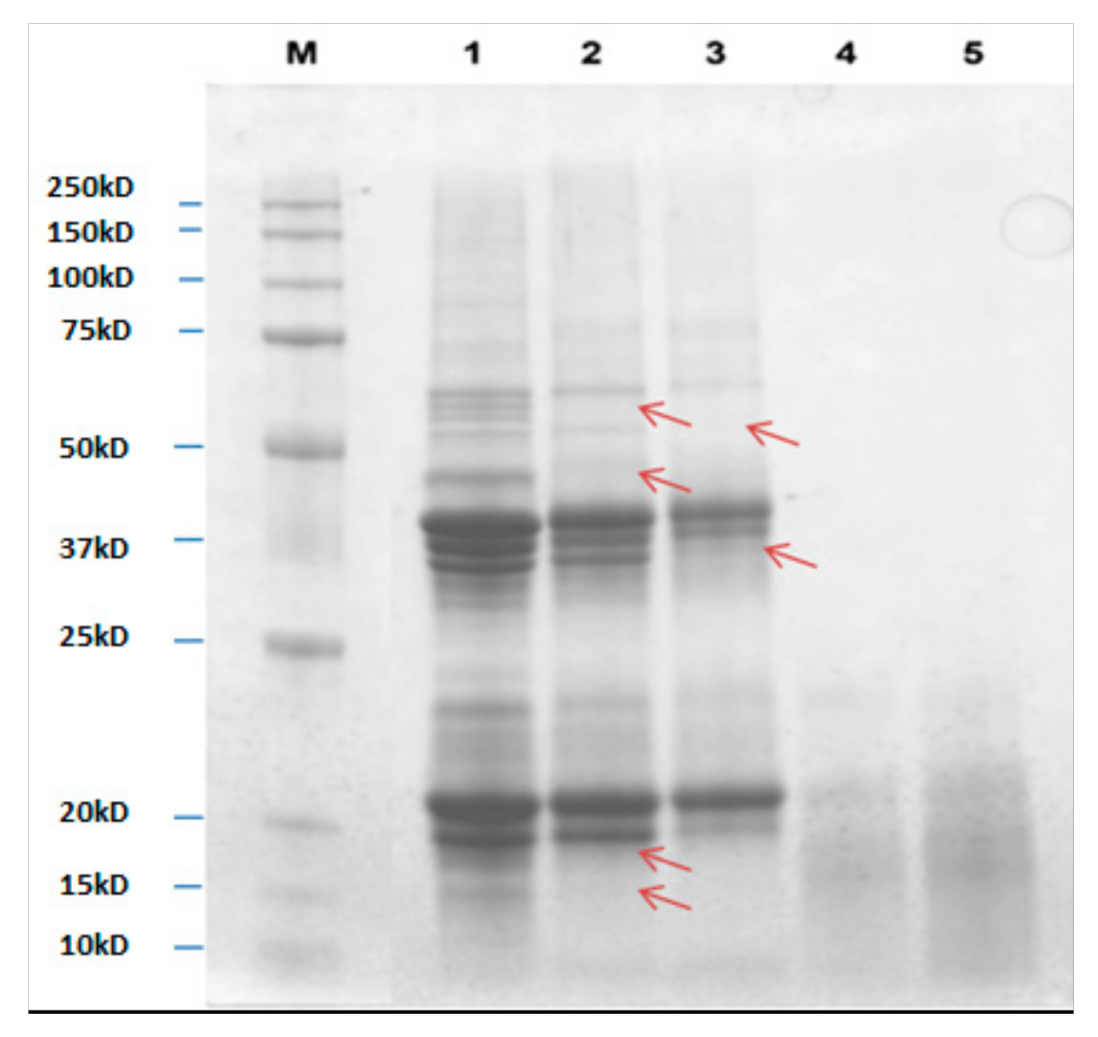
3.3. Protein Profiling of Untreated and Autoclaved Almond by SDS-PAGE Analysis
3.4. Fate of Heat/Pressured Almonds Proteins upon In Vitro Gastro-Duodenal Digestion and Evaluation of Residual Immunoreactivity
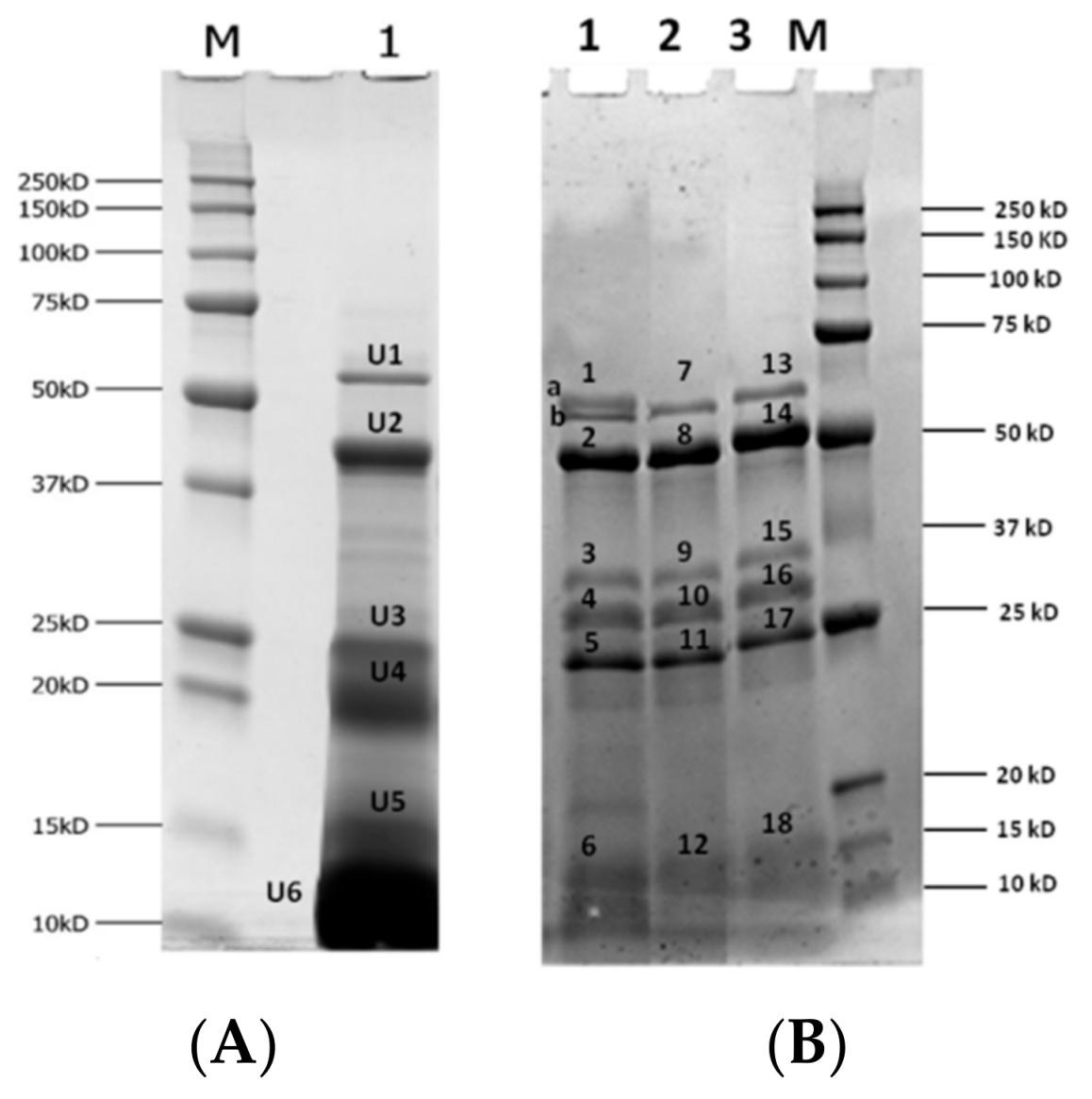
3.4.1. Digestibility of Almond Proteins upon Simulated Gastric and Duodenal Digestion
3.4.2. Assessment of Residual Immunoreactivity of Thermally/Pressure Treated Almonds Submitted to In Vitro Digested
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Costa, J.; Mafra, I.; Carrapatoso, I.; Oliveira, M.B.P.P. Almond allergens: Molecular characterization, detection, and clinical relevance. J. Agric. Food Chem. 2012, 60, 1337–1349. [Google Scholar] [CrossRef] [PubMed]
- Bolling, B.W.; Chen, C.Y.O.; McKay, D.L.; Blumberg, J.B. Tree nut phytochemicals: Composition, antioxidant capacity, bioactivity, impact factors. A systematic review of almonds, Brazils, cashews, hazelnuts, macadamias, pecans, pine nuts, pistachios and walnuts. Nutr. Res. Rev. 2011, 24, 244–275. [Google Scholar] [CrossRef] [PubMed]
- Yada, S.; Lapsley, K.; Huang, G. A review of composition studies of cultivated almonds: Macronutrients and micronutrients. J. Food Compos. Anal. 2011, 24, 469–480. [Google Scholar] [CrossRef]
- Fernández-Cuesta, A.; Kodad, O.; Velasco, L. Phytosterol variability in almond germplasm. J. Am. Soc. Hortic. Sci. 2012, 137, 343–348. [Google Scholar]
- FAOSTAT. Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/home/en/ (accessed on 25 September 2018).
- Elizur, A.; Appel, M.Y.; Nachshon, L.; Levy, M.B.; Epstein-Rigbi, N.; Golobov, K.; Goldberg, M.R. NUT Co Reactivity—Acquiring Knowledge for Elimination Recommendations (NUT CRACKER) study. Allergy Eur. J. Allergy Clin. Immunol. 2018, 73, 593–601. [Google Scholar] [CrossRef] [PubMed]
- McWilliam, V.; Koplin, J.; Lodge, C.; Tang, M.; Dharmage, S.; Allen, K. The Prevalence of Tree Nut Allergy: A Systematic Review. Curr. Allergy Asthma Rep. 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Allergen Nomenclature, Allergen Nomenclature IUIS Allergen Nomenclature Sub-Committee. Available online: http://www.allergen.org (accessed on 18 September 2018).
- Roux, K.H.; Teuber, S.S.; Robotham, J.M.; Sathe, S.K. Detection and stability of the major almond allergen in foods. J. Agric. Food Chem. 2001, 49, 2131–2136. [Google Scholar] [CrossRef] [PubMed]
- Sathe, S.K.; Wolf, W.J.; Roux, K.H.; Teuber, S.S.; Venkatachalam, M.; Sze-Tao, K.W.C. Biochemical characterization of amandin, the major storage protein in almond (Prunus Dulcis L.). J. Agric. Food Chem. 2002, 50, 4333–4341. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Mas, J.; Messeguer, R.; Arús, P.; Puigdomènech, P. Molecular characterization of cDNAs corresponding to genes expressed during almond (Prunus amygdalus Batsch) seed development. Plant Mol. Biol. 1995, 27, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Albillos, S.M.; Jin, T.; Howard, A.; Zhang, Y.; Kothary, M.H.; Fu, T.J. Purification, crystallization and preliminary X-ray characterization of prunin-1, a major component of the almond (Prunus dulcis) allergen amandin. J. Agric. Food Chem. 2008, 56, 5352–5358. [Google Scholar] [CrossRef] [PubMed]
- Alasalvar, C.; Shahidi, F. (Eds.) Tree Nuts: Composition, Phytochemicals, and Health Effects; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2008; pp. 1–326. ISBN 978-1-4200-1939. [Google Scholar]
- EU Regulation, No. 1169/2011 of the European Parliament and of the Council of 25 October 2011 on the provision of food information to consumers. Off. J. Eur. Union 2011, 50, 18–63. [Google Scholar]
- Monaci, L.; De Angelis, E.; Montemurro, N.; Pilolli, R. Comprehensive overview and recent advances in proteomics MS based methods for food allergens analysis. TrAC Trends Anal. Chem. 2018, 106, 21–36. [Google Scholar] [CrossRef]
- Andjelković, U.; Gavrović-Jankulović, M.; Martinović, T.; Josić, D. Omics methods as a tool for investigation of food allergies. TrAC Trends Anal. Chem. 2017, 96, 107–115. [Google Scholar] [CrossRef]
- Pilolli, R.; De Angelis, E.; Monaci, L. Streamlining the analytical workflow for multiplex MS/MS allergen detection in processed foods. Food Chem. 2017, 221, 1747–1753. [Google Scholar] [CrossRef] [PubMed]
- Pilolli, R.; De Angelis, E.; Monaci, L. In house validation of a high resolution mass spectrometry Orbitrap-based method for multiple allergen detection in a processed model food. Anal. Bioanal. Chem. 2018, 410, 5653–5662. [Google Scholar] [CrossRef] [PubMed]
- Rahaman, T.; Vasiljevic, T.; Ramchandran, L. Effect of processing on conformational changes of food proteins related to allergenicity. Trends Food Sci. Technol. 2016, 49, 24–34. [Google Scholar] [CrossRef]
- Su, M.; Liu, C.; Roux, K.H.; Gradziel, T.M.; Sathe, S.K. Effects of processing and storage on almond (Prunus dulcis L.) amandin immunoreactivity. Food Res. Int. 2017, 100, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, M.; Teuber, S.S.; Roux, K.H.; Sathe, S.K. Effects of roasting, blanching, autoclaving, and microwave heating on antigenicity of almond (Prunus dulcis L.) proteins. J. Agric. Food Chem. 2002, 50, 3544–3548. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, J.; Sheng, W.; Wang, S.; Fu, T.J. Effects of heat and high-pressure treatments on the solubility and immunoreactivity of almond proteins. Food Chem. 2016, 199, 856–861. [Google Scholar] [CrossRef] [PubMed]
- Shriver, S.K.; Yang, W.W. Thermal and Nonthermal Methods for Food Allergen Control. Food Eng. Rev. 2011, 3, 26–43. [Google Scholar] [CrossRef]
- Albillos, S.M.; Menhart, N.; Fu, T.J. Structural stability of amandin, a major allergen from almond (Prunus dulcis), and its acidic and basic polypeptides. J. Agric. Food Chem. 2009, 57, 4698–4705. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Venkatachalam, M.; Teuber, S.S.; Roux, K.H.; Sathe, S.K. Impact of γ-irradiation and thermal processing on the antigenicity of almond, cashew nut and walnut proteins. J. Sci. Food Agric. 2004, 84, 1119–1125. [Google Scholar] [CrossRef]
- Li, Y.; Yang, W.; Chung, S.Y.; Chen, H.; Ye, M.; Teixeira, A.A.; Gregory, J.F.; Welt, B.A.; Shriver, S. Effect of Pulsed Ultraviolet Light and High Hydrostatic Pressure on the Antigenicity of Almond Protein Extracts. Food Bioprocess Technol. 2013, 6, 431–440. [Google Scholar] [CrossRef]
- Dhakal, S.; Liu, C.; Zhang, Y.; Roux, K.H.; Sathe, S.K.; Balasubramaniam, V.M. Effect of high pressure processing on the immunoreactivity of almond milk. Food Res. Int. 2014, 62, 215–222. [Google Scholar] [CrossRef]
- Bøgh, K.L.; Madsen, C.B. Food Allergens: Is There a Correlation between Stability to Digestion and Allergenicity? Crit. Rev. Food Sci. Nutr. 2016, 56, 1545–1567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food-an international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Genetically Modified Organisms (GMO), Draft Scientific Opinion on the assessment of allergenicity of GM plants and microorganisms and derived food and feed. EFSA J. 2010, 8, 1–168. [CrossRef]
- Mandalari, G.; Faulks, R.M.; Rich, G.T.; Lo Turco, V.; Picout, D.R.; Lo Curto, R.B.; Bisignano, G.; Dugo, P.; Dugo, G.; Waldron, K.W.; et al. Release of protein, lipid, and vitamin E from almond seeds during digestion. J. Agric. Food Chem. 2008, 56, 3409–3416. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Grundy, M.M.L.; Grassby, T.; Parker, M.L.; Cross, K.L.; Chessa, S.; Bisignano, C.; Barreca, D.; Bellocco, E.; Laganà, G.; et al. The effects of processing and mastication on almond lipid bioaccessibility using novel methods of in vitro digestion modelling and micro-structural analysis. Br. J. Nutr. 2014, 112, 1521–1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandalari, G.; Parker, M.L.; Grundy, M.M.L.; Grassby, T.; Smeriglio, A.; Bisignano, C.; Raciti, R.; Trombetta, D.; Baer, D.J.; Wilde, P.J. Understanding the effect of particle size and processing on almond lipid bioaccessibility through microstructural analysis: From mastication to faecal collection. Nutrients 2018, 10, 213. [Google Scholar] [CrossRef] [PubMed]
- Grassby, T.; Mandalari, G.; Grundy, M.M.L.; Edwards, C.H.; Bisignano, C.; Trombetta, D.; Smeriglio, A.; Chessa, S.; Ray, S.; Sanderson, J.; et al. In vitro and in vivo modeling of lipid bioaccessibility and digestion from almond muffins: The importance of the cell-wall barrier mechanism. J. Funct. Foods 2017, 37, 263–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grassby, T.; Picout, D.R.; Mandalari, G.; Faulks, R.M.; Kendall, C.W.C.; Rich, G.T.; Wickham, M.S.J.; Lapsley, K.; Ellis, P.R. Modelling of nutrient bioaccessibility in almond seeds based on the fracture properties of their cell walls. Food Funct. 2014, 5, 3096–3106. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Tomaino, A.; Rich, G.T.; Lo Curto, R.; Arcoraci, T.; Martorana, M.; Bisignano, C.; Saija, A.; Parker, M.L.; Waldron, K.W.; et al. Polyphenol and nutrient release from skin of almonds during simulated human digestion. Food Chem. 2010, 122, 1083–1088. [Google Scholar] [CrossRef]
- Mandalari, G.; Vardakou, M.; Faulks, R.; Bisignano, C.; Martorana, M.; Smeriglio, A.; Trombetta, D. Food matrix effects of polyphenol bioaccessibility from almond skin during simulated human digestion. Nutrients 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Toomer, O.T.; Do, A.; Pereira, M.; Williams, K. Effect of simulated gastric and intestinal digestion on temporal stability and immunoreactivity of peanut, almond, and pine nut protein allergens. J. Agric. Food Chem. 2013, 61, 5903–5913. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Rigby, N.M.; Bisignano, C.; Lo Curto, R.B.; Mulholland, F.; Su, M.; Venkatachalam, M.; Robotham, J.M.; Willison, L.N.; Lapsley, K.; et al. Effect of food matrix and processing on release of almond protein during simulated digestion. LWT Food Sci. Technol. 2014, 59, 439–447. [Google Scholar] [CrossRef]
- Bavaro, S.L.; Di Stasio, L.; Mamone, G.; De Angelis, E.; Nocerino, R.; Canani, R.B.; Logrieco, A.F.; Montemurro, N.; Monaci, L. Effect of thermal/pressure processing and simulated human digestion on the immunoreactivity of extractable peanut allergens. Food Res. Int. 2018, 109, 126–137. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, E.; Pilolli, R.; Bavaro, S.L.; Monaci, L. Insight into the gastro-duodenal digestion resistance of soybean proteins and potential implications for residual immunogenicity. Food Funct. 2017, 8, 1599–1610. [Google Scholar] [CrossRef] [PubMed]
- Masthoff, L.J.; Hoff, R.; Verhoeckx, K.C.M.; van Os-Medendorp, H.; Michelsen-Huisman, A.; Baumert, J.L.; Pasmans, S.G.; Meijer, Y.; Knulst, A.C. A systematic review of the effect of thermal processing on the allergenicity of tree nuts. Allergy 2013, 68, 983–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabanillas, B.; Cuadrado, C.; Rodriguez, J.; Hart, J.; Burbano, C.; Crespo, J.F.; Novak, N. Potential changes in the allergenicity of three forms of peanut after thermal processing. Food Chem. 2015, 183, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Cabanillas, B.; Maleki, S.J.; Rodríguez, J.; Cheng, H.; Teuber, S.S.; Wallowitz, M.L.; Muzquiz, M.; Pedrosa, M.M.; Linacero, R.; Burbano, C.; et al. Allergenic properties and differential response of walnut subjected to processing treatments. Food Chem. 2014, 157, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Awuah, G.B.; Ramaswamy, H.S.; Economides, A. Thermal processing and quality: Principles and overview. Chem. Eng. Process. Process Intensif. 2007, 46, 584–602. [Google Scholar] [CrossRef]
- Ioannou, I.; Hafsa, I.; Hamdi, S.; Charbonnel, C.; Ghoul, M. Review of the effects of food processing and formulation on flavonol and anthocyanin behaviour. J. Food Eng. 2012, 111, 208–217. [Google Scholar] [CrossRef]
- Lešková, E.; Kubíková, J.; Kováčiková, E.; Košická, M.; Porubská, J.; Holčíková, K. Vitamin losses: Retention during heat treatment and continual changes expressed by mathematical models. J. Food Compos. Anal. 2006, 19, 252–276. [Google Scholar] [CrossRef]
- Tiwari, R.S.; Venkatachalam, M.; Sharma, G.M.; Su, M.; Roux, K.H.; Sathe, S.K. Effect of food matrix on amandin, almond (Prunus dulcis L.) major protein, immunorecognition and recovery. LWT Food Sci. Technol. 2010, 43, 675–683. [Google Scholar] [CrossRef]
- Mandalari, G.; Genovese, T.; Bisignano, C.; Mazzon, E.; Wickham, M.S.J.; Di Paola, R.; Bisignano, G.; Cuzzocrea, S. Neuroprotective effects of almond skins in experimental spinal cord injury. Clin. Nutr. 2011, 30, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Bisignano, C.; Genovese, T.; Mazzon, E.; Wickham, M.S.J.; Paterniti, I.; Cuzzocrea, S. Natural almond skin reduced oxidative stress and inflammation in an experimental model of inflammatory bowel disease. Int. Immunopharmacol. 2011, 11, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Ozdal, T.; Capanoglu, E.; Altay, F. A review on protein-phenolic interactions and associated changes. Food Res. Int. 2013, 51, 954–970. [Google Scholar] [CrossRef]
- Jakobek, L. Interactions of polyphenols with carbohydrates, lipids and proteins. Food Chem. 2015, 175, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Naegeli, H.; Birch, A.N.; Casacuberta, J.; De Schrijver, A.; Gralak, M.A.; Guerche, P.; Jones, H.; Manachini, B.; Messéan, A.; Nielsen, E.E.; et al. EFSA Panel on Genetically Modified Organisms (GMO). EFSA J. 2017, 15, 4862–4911. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Band | Accession Number | Type of Protein (Organism) | Allergen | Coverage (%) | Unique Peptides | Score |
|---|---|---|---|---|---|---|---|
| CTRL-NE | U1 | Q945K2 | (R)-mandelonitrile lyase 2 (Prunus dulcis (Mill.) D. A. Webb) | 30.37 | 18 (18) | 30.34 | |
| U2 | Q945K2 | (R)-mandelonitrile lyase 2 (Prunus dulcis (Mill.) D. A. Webb) | 11.55 | 7 (7) | 0 | ||
| U3 | Q9SW89 | Abscisic acid response protein (Prunus dulcis (Mill.) D. A. Webb) | 43.07 | 8 (8) | 19.35 | ||
| U4 | Q43607 | Prunin (Prunus dulcis (Mill.) D. A. Webb) | Pru du 6 | 12.16 | 8 (8) | 1.65 | |
| U5 | Q43607 | Prunin (Prunus dulcis (Mill.) D. A. Webb) | Pru du 6 | 8.53 | 6 (1) | 8.59 | |
| E3SH28 | Prunin 1 (Prunus dulcis (Mill.) D. A. Webb) | Pru du 6, Pru du 6.0101 | 8.17 | 6 (1) | 6.94 | ||
| U6 | Q43607 | Prunin (Prunus dulcis (Mill.) D. A. Webb) | Pru du 6 | 16.33 | 11 (2) | 21.8 | |
| E3SH28 | Prunin 1 (Prunus dulcis (Mill.) D. A. Webb) | Pru du 6, Pru du 6.0101 | 15.25 | 10 (1) | 16.56 | ||
| A7Y7K3 | Putative lipid-transfer protein (Prunus dulcis (Mill.) D. A. Webb) | 40.91 | 3 (3) | 9.77 | |||
| E3SH29 | Prunin 2 (Fragment) (Prunus dulcis (Mill.) D. A. Webb) | Pru du 6, Pru du 6.0201 | 13.10 | 7 (7) | 4.43 | ||
| CTRL-GI | 1 | Q945K2 | (R)-mandelonitrile lyase 2 (Prunus dulcis (Mill.) D. A. Webb) | 36.41 | 31 (31) | 17.64 | |
| P06278 | Alpha-amylase (Bacillus licheniformis) | 9.96 | 10 (10) | 1.77 | |||
| 2 | P00690 | Pancreatic alpha-amylase (Sus scrofa Linnaeus) | 70.45 | 55 (28) | 100.16 | ||
| P04745 | Alpha-amylase (Homo sapiens Linnaeus) | 33.27 | 26 (4) | 58.45 | |||
| Q945K2 | (R)-mandelonitrile lyase 2 (Prunus dulcis (Mill.) D. A. Webb) | 21.31 | 12 (12) | 1.83 | |||
| 3 | P00690 | Pancreatic alpha-amylase (Sus scrofa Linnaeus) | 15.66 | 8 (8) | 0 | ||
| 4 | P00690 | Pancreatic alpha-amylase (Sus scrofa Linnaeus) | 25.83 | 21 (20) | 10.21 | ||
| Q7M3E1 | Chymotrypsin-C (Bos taurus Linnaeus) | 7.09 | 3 (2) | 3.18 | |||
| 6 | P00761 | Trypsin (Sus scrofa Linnaeus) | 22.08 | 5 (4) | 3.73 | ||
| AC10-GI | 7 | P06278 | Alpha-amylase (Bacillus licheniformis) | 3.32 | 3 (3) | 2.05 | |
| 8 | P00690 | Pancreatic alpha-amylase (Sus scrofa Linnaeus) | 60.86 | 47 (23) | 80.76 | ||
| 9 | P00690 | Pancreatic alpha-amylase (Sus scrofa Linnaeus) | 33.86 | 24 (23) | 18.86 | ||
| 10 | P00690 | Pancreatic alpha-amylase (Sus scrofa Linnaeus) | 19.37 | 13 (13) | 10.92 | ||
| 11 | P00766 | Chymotrypsinogen A (Bos taurus) | 15.10 | 5 (4) | 3.53 | ||
| 12 | P00761 | Trypsin (Sus scrofa Linnaeus) | 18.61 | 5 (4) | 6.36 | ||
| P00690 | Pancreatic alpha-amylase (Sus scrofa Linnaeus) | 13.50 | 10 (9) | 5.46 | |||
| H2O-AC10-GI | 14 | P00690 | Pancreatic alpha-amylase (Sus scrofa Linnaeus) | 50.68 | 44 (22) | 58.65 | |
| 15 | P00690 | Pancreatic alpha-amylase (Sus scrofa Linnaeus) | 30.14 | 20 (19) | 9.4 | ||
| 16 | P00690 | Pancreatic alpha-amylase (Sus scrofa Linnaeus) | 19.77 | 14 (14) | 9.04 | ||
| Q7M3E1 | Chymotrypsin-C (Bos taurus Linnaeus) | 7.09 | 3 (3) | 5.08 | |||
| 17 | P00766 | Chymotrypsinogen A (Bos taurus Linnaeus) | 11.02 | 3 (2) | 9.49 | ||
| 18 | P00761 | Trypsin (Sus scrofa Linnaeus) | 39.83 | 9 (9) | 11.92 | ||
| P00690 | Pancreatic alpha-amylase (Sus scrofa Linnaeus) | 12.72 | 7 (7) | 6.03 |
| Sample | Accession Number | Type of Protein (Organism) | Allergen | Coverage (%) | Unique Peptides | Score |
|---|---|---|---|---|---|---|
| CTRL-GI | C0L0I5 | Non-specific lipid-transfer protein (Prunus dulcis (Mill.) D. A. Webb) | Pru du 3, Pru du 3.0101 | 40.65 | 11 (10) | 4.11 |
| P82944 | Seed allergenic protein 1 (Fragments) (Prunus dulcis (Mill.) D. A. Webb) | Pru du 2S Albumin | 60.71 | 5 (4) | 10.21 | |
| Q43608 | Pru2 protein (Fragment) (Prunus dulcis (Mill.) D. A. Webb) | Pru du 6 | 71.83 | 195 (7) | 297.12 | |
| E3SH28 | Prunin 1 (Prunus dulcis (Mill.) D. A. Webb) | Pru du 6, Pru du 6.0101 | 80.76 | 323 (4) | 672.17 | |
| Q8GSL5 | Profilin (Prunus dulcis (Mill.) D. A. Webb) | Pru du 4 | 40.46 | 17 (17) | 9.7 | |
| P82952 | Seed allergenic protein 2 (Fragment) (Prunus dulcis (Mill.) D. A. Webb) | Pru du AP | 48.00 | 15 (15) | 39.02 | |
| Q43607 | Prunin (Prunus dulcis (Mill.) D. A. Webb) | Pru du 6 | 81.85 | 335 (14) | 697.42 | |
| Q8H2B9 | 60S acidic ribosomal protein (Prunus dulcis (Mill.) D. A. Webb) | Pru du 5, Pru du 5.0101 | 37.17 | 23 (22) | 9.42 | |
| AC10-GI | E3SH29 | Prunin 2 (Fragment) (Prunus dulcis (Mill.) D. A. Webb) | Pru du 6, Pru du 6.0201 | 68.65 | 213 (4) | 363.33 |
| C0L0I5 | Non-specific lipid-transfer protein (Prunus dulcis (Mill.) D. A. Webb) | Pru du 3, Pru du 3.0101 | 32.52 | 9 (9) | 2.57 | |
| P82944 | Seed allergenic protein 1 (Fragments) (Prunus dulcis (Mill.) D. A. Webb) | Pru du 2S Albumin | 60.71 | 5 (4) | 11.76 | |
| Q43608 | Pru2 protein (Fragment) (Prunus dulcis (Mill.) D. A. Webb) | Pru du 6 | 69.05 | 209 (6) | 359.74 | |
| E3SH28 | Prunin 1 (Prunus dulcis (Mill.) D. A. Webb) | Pru du 6, Pru du 6.0101 | 78.40 | 292 (4) | 700.81 | |
| Q8GSL5 | Profilin (Prunus dulcis (Mill.) D. A. Webb) | Pru du 4 | 30.53 | 13 (13) | 4.48 | |
| P82952 | Seed allergenic protein 2 (Fragment) (Prunus dulcis (Mill.) D. A. Webb) | Pru du AP | 48.00 | 10 (10) | 17.94 | |
| Q43607 | Prunin (Prunus dulcis (Mill.) D. A. Webb) | Pru du 6 | 78.77 | 300 (10) | 712.31 | |
| Q8H2B9 | 60S acidic ribosomal protein (Prunus dulcis (Mill.) D. A. Webb) | Pru du 5, Pru du 5.0101 | 37.17 | 13 (13) | 5.05 | |
| H2O-AC10-GI | E3SH29 | Prunin 2 (Fragment) (Prunus dulcis (Mill.) D. A. Webb) | Pru du 6, Pru du 6.0201 | 78.97 | 272 (16) | 630.16 |
| C0L0I5 | Non-specific lipid-transfer protein (Prunus dulcis (Mill.) D. A. Webb) | Pru du 3, Pru du 3.0101 | 27.64 | 7 (6) | 1.43 | |
| P82944 | Seed allergenic protein 1 (Fragments) (Prunus dulcis (Mill.) D. A. Webb) | Pru du 2S Albumin | 78.57 | 6 (5) | 8.77 | |
| Q43608 | Pru2 protein (Fragment) (Prunus dulcis (Mill.) D. A. Webb) | Pru du 6 | 77.58 | 261 (10) | 592.97 | |
| E3SH28 | Prunin 1 (Prunus dulcis (Mill.) D. A. Webb) | Pru du 6, Pru du 6.0101 | 83.30 | 321 (8) | 941.16 | |
| Q8GSL5 | Profilin (Prunus dulcis (Mill.) D. A. Webb) | Pru du 4 | 21.37 | 10 (10) | 8.1 | |
| P82952 | Seed allergenic protein 2 (Fragment) (Prunus dulcis (Mill.) D. A. Webb) | Pru du AP | 48.00 | 9 (9) | 21.22 | |
| Q43607 | Prunin (Prunus dulcis (Mill.) D. A. Webb) | Pru du 6 | 82.94 | 320 (5) | 954.46 | |
| Q8H2B9 | 60S acidic ribosomal protein (Prunus dulcis (Mill.) D. A. Webb) | Pru du 5, Pru du 5.0101 | 37.17 | 16 (16) | 8.63 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Angelis, E.; Bavaro, S.L.; Forte, G.; Pilolli, R.; Monaci, L. Heat and Pressure Treatments on Almond Protein Stability and Change in Immunoreactivity after Simulated Human Digestion. Nutrients 2018, 10, 1679. https://doi.org/10.3390/nu10111679
De Angelis E, Bavaro SL, Forte G, Pilolli R, Monaci L. Heat and Pressure Treatments on Almond Protein Stability and Change in Immunoreactivity after Simulated Human Digestion. Nutrients. 2018; 10(11):1679. https://doi.org/10.3390/nu10111679
Chicago/Turabian StyleDe Angelis, Elisabetta, Simona L. Bavaro, Graziana Forte, Rosa Pilolli, and Linda Monaci. 2018. "Heat and Pressure Treatments on Almond Protein Stability and Change in Immunoreactivity after Simulated Human Digestion" Nutrients 10, no. 11: 1679. https://doi.org/10.3390/nu10111679
APA StyleDe Angelis, E., Bavaro, S. L., Forte, G., Pilolli, R., & Monaci, L. (2018). Heat and Pressure Treatments on Almond Protein Stability and Change in Immunoreactivity after Simulated Human Digestion. Nutrients, 10(11), 1679. https://doi.org/10.3390/nu10111679