The Effect of Dietary Mushroom Agaricus bisporus on Intestinal Microbiota Composition and Host Immunological Function


Abstract
:1. Introduction
2. Experimental Design
2.1. Animals and Diets
2.2. Immune Parameters
2.3. Ex-Vivo Measurement of Intestinal Epithelial Cell Function
2.4. Intestinal Sample Processing
2.5. Fecal Specimen Collection and Processing for 16S rDNA Amplicon Multi-Tag Sequencing
2.6. Predicted Metabolic Profiles of Microbiome
2.7. Statistical Analyses
3. Results
3.1. Clinical Signs
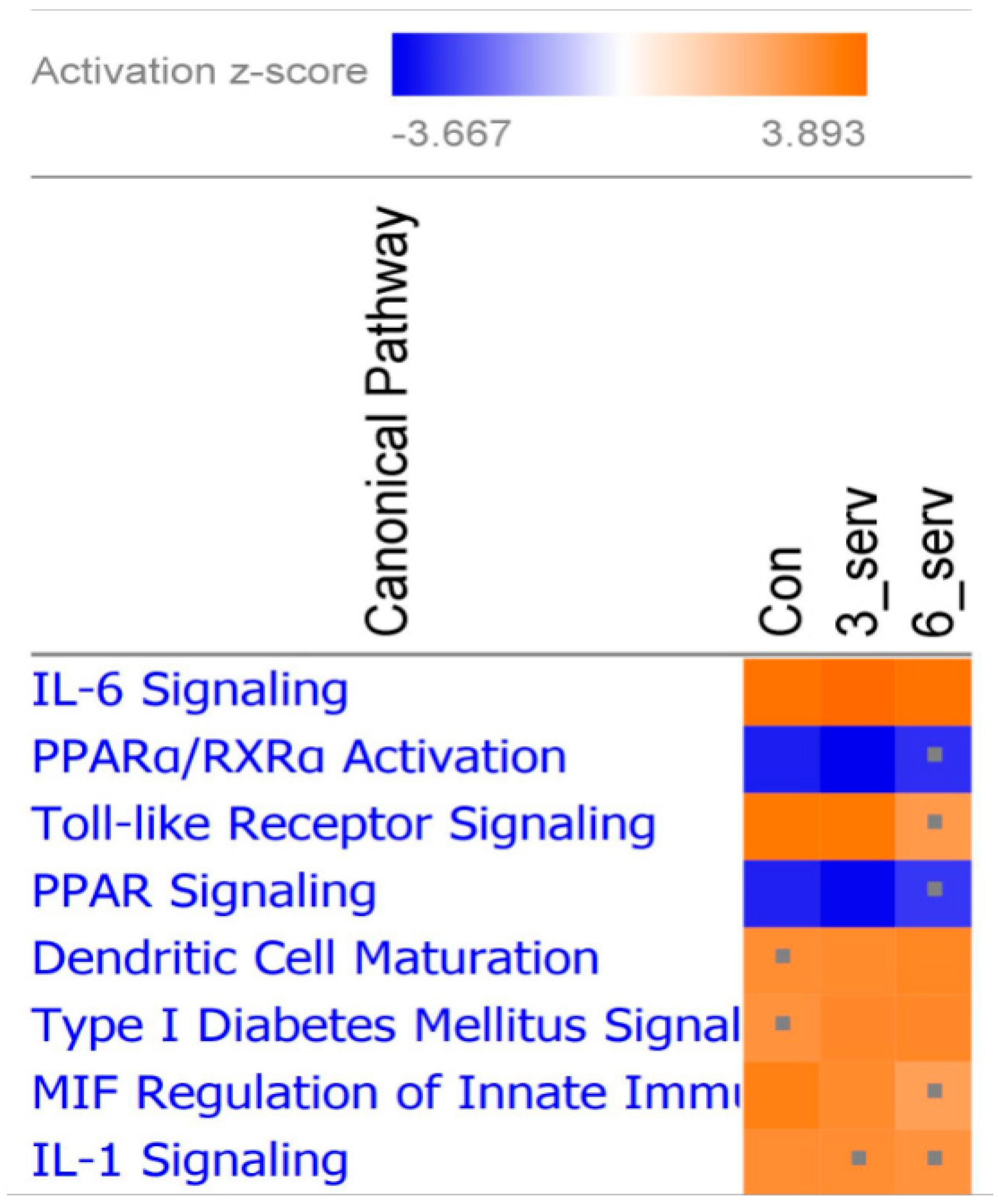
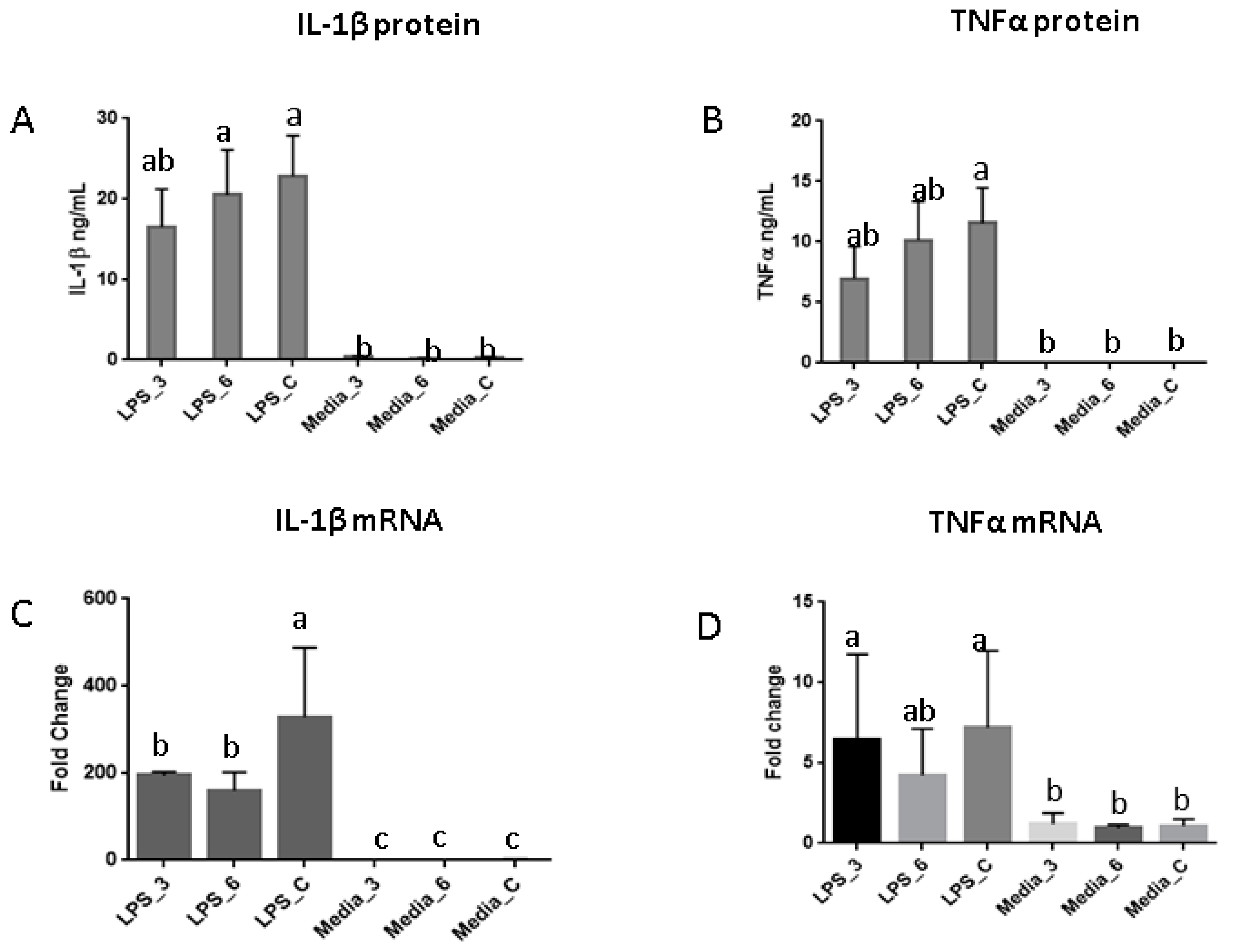
3.2. Immune Response Evaluation
3.3. Intestinal Response
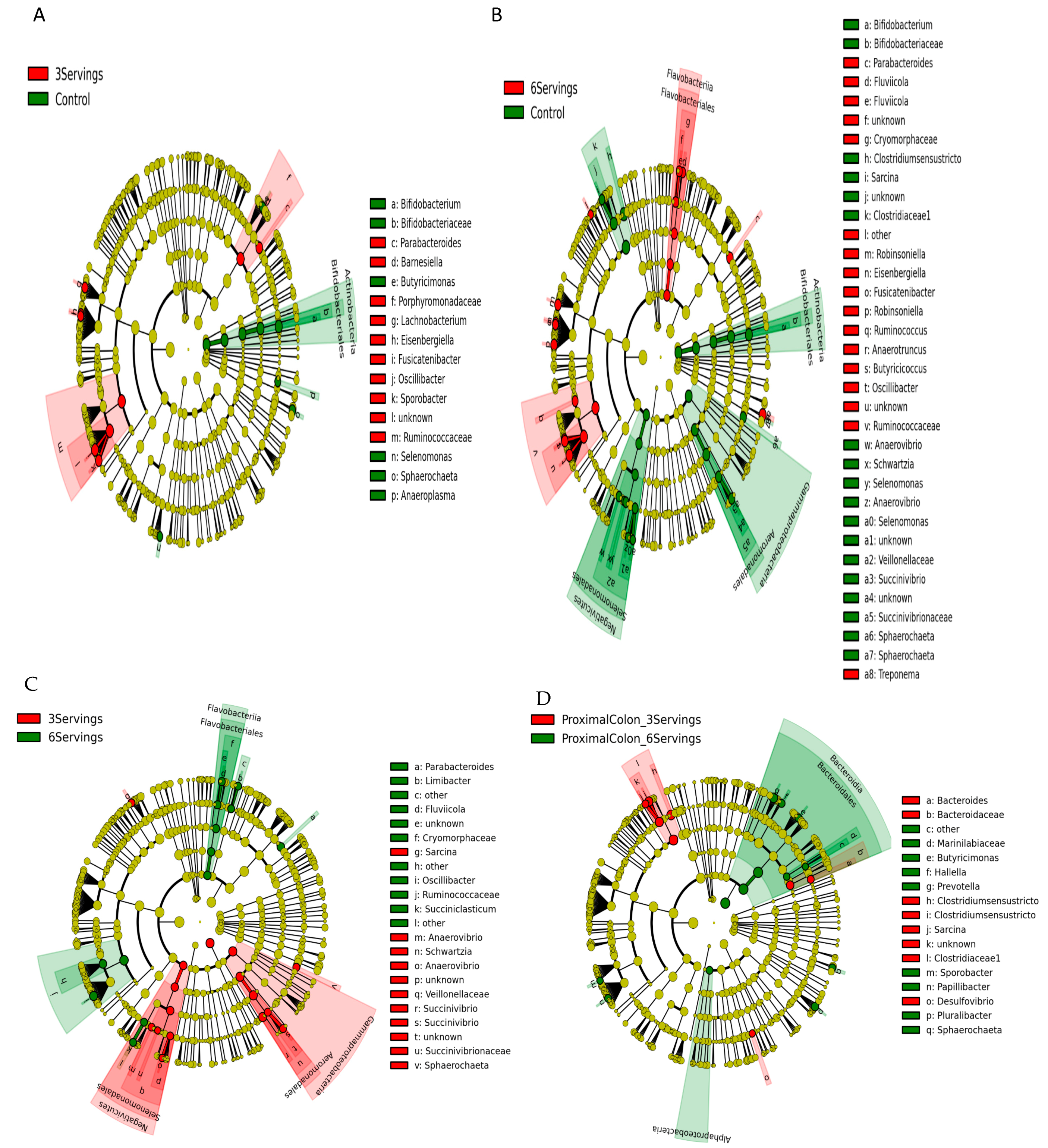
3.4. Discriminant Bacterial Microbiota at Six Weeks after Diets Supplemented with Three or Six Servings of Mushrooms
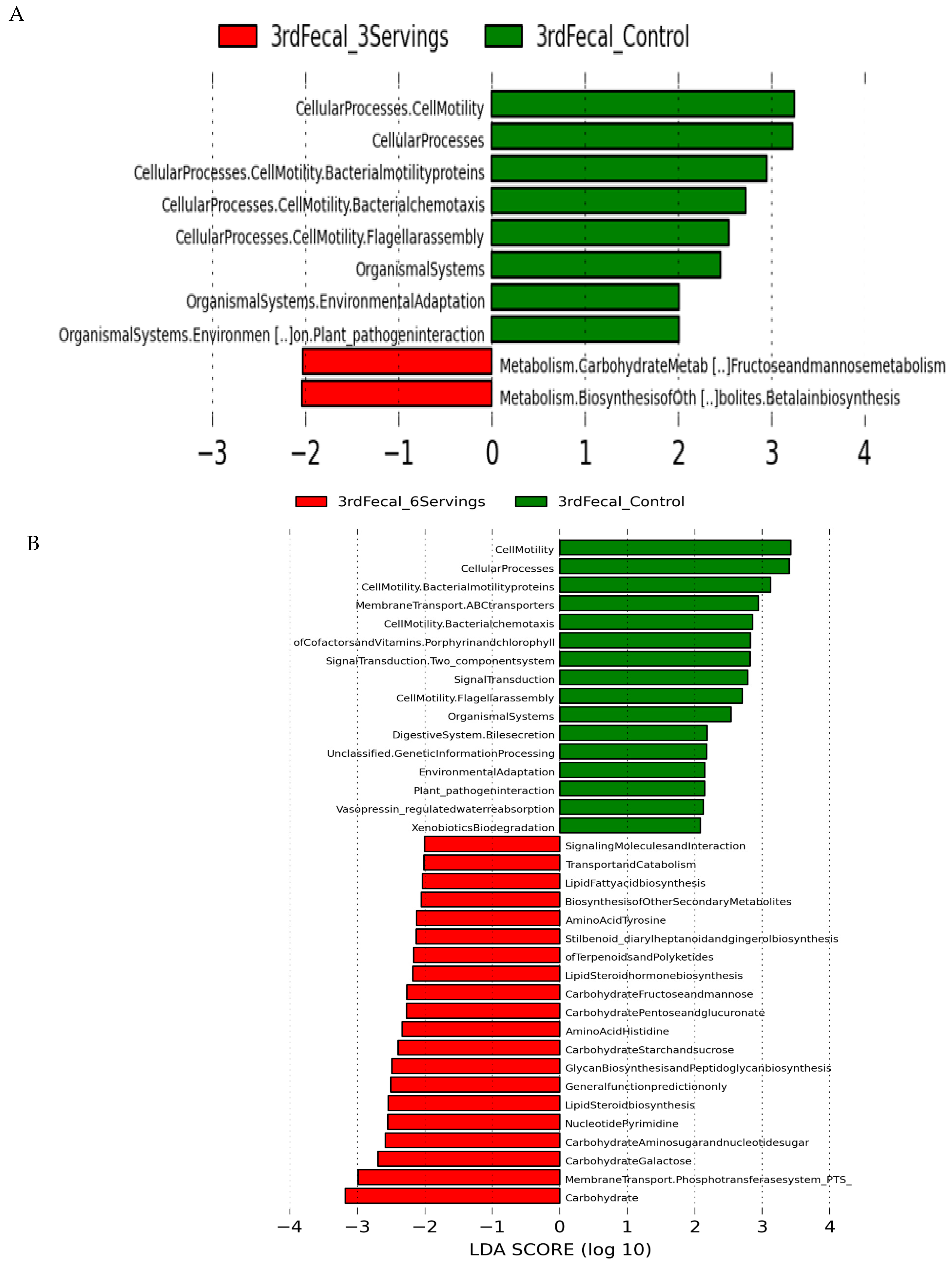
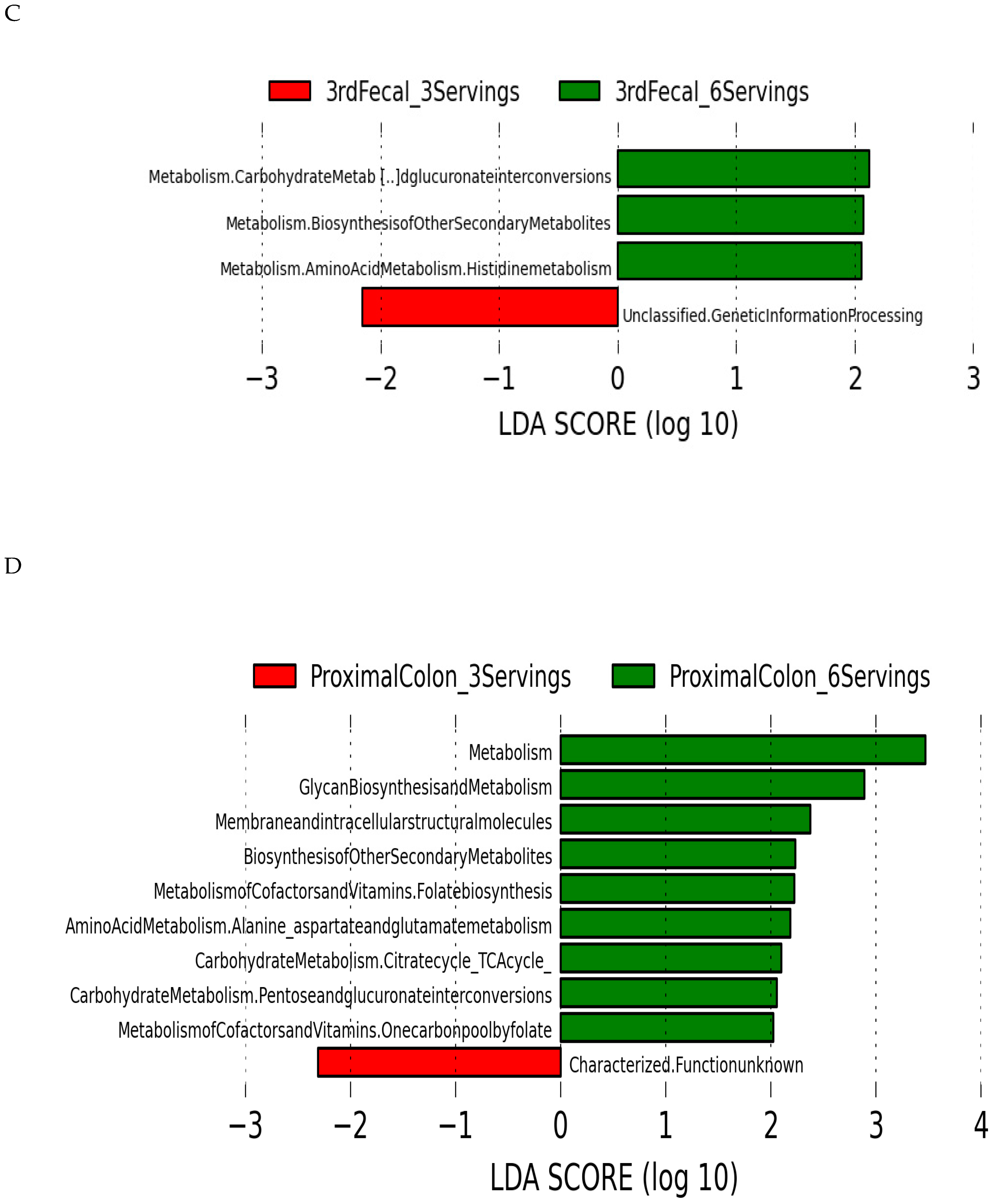
3.5. Predicted Microbial Function
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| QIIME | Quantitative Insights into Microbial Ecology |
| LDA | Linear Discriminant Analysis |
| LEfSe | Effect Size |
| PICRUSt | Phylogenetic Investigation of Communities by Reconstruction of Unobserved States |
References
- Amirullah, N.A.; Abidin, N.Z.; Abdullah, N. The potential applications of mushrooms against some facets of atherosclerosis: A review. Food Res. Int. 2018, 105, 517–536. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y. Protective effect of polysaccharide from Agaricus bisporus in Tibet area of China against tetrachloride-induced acute liver injury in mice. Int. J. Biol. Macromol. 2018, 118, 1488–1493. [Google Scholar] [CrossRef] [PubMed]
- Smiderle, F.R. Polysaccharides from Agaricus bisporus and Agaricus brasiliensis show similarities in their structures and their immunomodulatory effects on human monocytic THP-1 cells. BMC Complement. Altern. Med. 2011, 11, 58. [Google Scholar] [CrossRef] [PubMed]
- Ditamo, Y. In vivo immunomodulatory effect of the lectin from edible mushroom Agaricus bisporus. Food Funct. 2016, 7, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.C. White button mushroom (Agaricus bisporus) lowers blood glucose and cholesterol levels in diabetic and hypercholesterolemic rats. Nutr. Res. 2010, 30, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Caz, V. Modulation of cholesterol-related gene expression by dietary fiber fractions from edible mushrooms. J. Agric. Food Chem. 2015, 63, 7371–7380. [Google Scholar] [CrossRef] [PubMed]
- Wasser, S.P. Medicinal mushrooms in human clinical studies. Part I. anticancer, oncoimmunological, and immunomodulatory activities: A Review. Int. J. Med. Mushrooms 2017, 19, 279–317. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M. Mushroom polysaccharides: Chemistry and antiobesity, antidiabetes, anticancer, and antibiotic properties in cells, rodents, and humans. Foods 2016, 5, 80. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Nie, S. The structure of mushroom polysaccharides and their beneficial role in health. Food Funct. 2015, 6, 3205–3217. [Google Scholar] [CrossRef] [PubMed]
- Martel, J. Immunomodulatory properties of plants and mushrooms. Trends Pharmacol. Sci. 2017, 38, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.C. Immunomodulatory properties of medicinal mushrooms: Differential effects of water and ethanol extracts on NK cell-mediated cytotoxicity. Innate Immun. 2016, 22, 522–533. [Google Scholar] [CrossRef] [PubMed]
- Komura, D.L. Structure of Agaricus spp. fucogalactans and their anti-inflammatory and antinociceptive properties. Bioresour. Technol. 2010, 101, 6192–6199. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y. Effect of dietary supplementation with white button mushrooms on host resistance to influenza infection and immune function in mice. Br. J. Nutr. 2013, 109, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z. White button mushroom enhances maturation of bone marrow-derived dendritic cells and their antigen presenting function in mice. J. Nutr. 2008, 138, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Varshney, J. White button mushrooms increase microbial diversity and accelerate the resolution of Citrobacter rodentium infection in mice. J. Nutr. 2013, 143, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Wang, J. Dietary supplementation with white button mushrooms augments the protective immune response to Salmonella vaccine in mice. J. Nutr. 2014, 144, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Forland, D.T. Effect of an extract based on the medicinal mushroom Agaricus blazei Murill on expression of cytokines and calprotectin in patients with ulcerative colitis and Crohn’s disease. Scand. J. Immunol. 2011, 73, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Therkelsen, S.P. Effect of the medicinal Agaricus blazei murill-based mushroom extract, andosantm, on symptoms, fatigue and quality of life in patients with crohn’s disease in a randomized single-blinded placebo controlled study. PLoS ONE 2016, 11, e0159288. [Google Scholar] [CrossRef] [PubMed]
- Therkelsen, S.P. Cytokine levels after consumption of a medicinal Agaricus blazei murill-based mushroom extract, andosan, in patients with crohn’s disease and ulcerative colitis in a randomized single-blinded placebo-controlled study. Scand. J. Immunol. 2016, 84, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, B.; Hadar, Y. Possible mechanisms of action of mushroom-derived glucans on inflammatory bowel disease and associated cancer. Ann. Transl. Med. 2014, 2, 19. [Google Scholar] [PubMed]
- Chang, C.J. Ganoderma lucidum reduces obesity in mice by modulating the composition of the gut microbiota. Nat. Commun. 2015, 6, 7489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.J. Antrodia cinnamomea reduces obesity and modulates the gut microbiota in high-fat diet-fed mice. Int. J. Obes. 2018, 42, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Yu, S. The effects of whole mushrooms during inflammation. BMC Immunol. 2009, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y. Macrophage immunomodulatory activity of the polysaccharide isolated from Collybia radicata mushroom. Int. J. Biol. Macromol. 2018, 108, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y. Prebiotic effects of white button mushroom (Agaricus bisporus) feeding on succinate and intestinal gluconeogenesis in C57BL/6 mice. J. Funct. Foods 2018, 45, 223–232. [Google Scholar] [CrossRef]
- Turnbaugh, P.J. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Backhed, F. Host-bacterial mutualism in the human intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.M. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Delzenne, N.M.; Cani, P.D. Interaction between obesity and the gut microbiota: Relevance in nutrition. Annu. Rev. Nutr. 2011, 31, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W. The dynamic distribution of porcine microbiota across different ages and gastrointestinal tract segments. PLoS ONE 2015, 10, e0117441. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Donovan, S.M. Human microbiota-associated swine: Current progress and future opportunities. ILAR J. 2015, 56, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, L.M.; Moeser, A.J.; Blikslager, A.T. Porcine models of digestive disease: The future of large animal translational research. Transl. Res. 2015, 166, 12–27. [Google Scholar] [CrossRef] [PubMed]
- Charepalli, V. Pigs, unlike mice, have two distinct colonic stem cell populations similar to humans that respond to high-calorie diet prior to insulin resistance. Cancer Prev. Res. 2017, 10, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Garthoff, L.H. The Autosow raised miniature swine as a model for assessing the effects of dietary soy trypsin inhibitor. Food Chem. Toxicol. 2002, 40, 487–500. [Google Scholar] [CrossRef]
- Pang, X. Inter-species transplantation of gut microbiota from human to pigs. ISME J. 2007, 1, 156–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L. A reference gene catalogue of the pig gut microbiome. Nat. Microbiol. 2016, 1, 16161. [Google Scholar] [CrossRef] [PubMed]
- Feeney, M.J. Mushrooms and health summit proceedings. J. Nutr. 2014, 144, S1128–S1136. [Google Scholar] [CrossRef] [PubMed]
- Chan, G.C.; Chan, W.K.; Sze, D.M. The effects of beta-glucan on human immune and cancer cells. J. Hematol. Oncol. 2009, 2, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, A.D.; Scholz, A.M.; Conway, J.M. Body composition analysis of small pigs by dual-energy x-ray absorptiometry. J. Anim. Sci. 1998, 76, 2392–2398. [Google Scholar] [CrossRef] [PubMed]
- Solano, G.I. Effect of porcine reproductive and respiratory syndrome virus infection on the clearance of Haemophilus parasuis by porcine alveolar macrophages. Can. J. Vet. Res. 1998, 62, 251–256. [Google Scholar] [PubMed]
- Jang, S. Flavanol-rich cocoa powder interacts with Lactobacillus rhamnossus LGG to alter the antibody response to infection with the parasitic nematode Ascaris suum. Nutrients 2017, 9, 1113. [Google Scholar] [CrossRef] [PubMed]
- Solano-Aguilar, G. Transcriptomic profile of whole blood cells from elderly subjects fed probiotic bacteria Lactobacillus rhamnosus GG ATCC 53103 (LGG) in a phase, I. open label study. PLoS ONE 2016, 11, e0147426. [Google Scholar] [CrossRef] [PubMed]
- Solano-Aguilar, G.I. Characterization of lymphocyte subsets from mucosal tissues in neonatal swine. Dev. Comp. Immunol. 2001, 25, 245–263. [Google Scholar] [CrossRef]
- Lammers, K.M. Gliadin induces an increase in intestinal permeability and zonulin release by binding to the chemokine receptor CXCR3. Gastroenterology 2008, 135, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Dawson, H.D. Localized multigene expression patterns support an evolving Th1/Th2-like paradigm in response to infections with Toxoplasma gondii and Ascaris suum. Infect. Immun. 2005, 73, 1116–1128. [Google Scholar] [CrossRef] [PubMed]
- Solano-Aguilar, G.I. The effect of feeding cocoa powder and Lactobacillus rhamnosus on the composition and function of pig intestinal microbiome. Curr. Dev. Nutr. 2018, 2, nzy011. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.J. Rifaximin exerts beneficial effects independent of its ability to alter microbiota composition. Clin. Transl. Gastroenterol. 2016, 7, e187. [Google Scholar] [CrossRef] [PubMed]
- Sikaroodi, M.; Gillevet, P.M. Quality control in multi-tag pyrosequencing of microbial communities. Biotechniques 2012, 53, 381–383. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://qiime2.org/ (accessed on 8 November 2018).
- Vazquez-Baeza, Y. EMPeror: A tool for visualizing high-throughput microbial community data. Gigascience 2013, 2, 16. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [PubMed]
- Langille, M.G. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markowitz, V.M. IMG: The integrated microbial genomes database and comparative analysis system. Nucleic. Acids. Res. 2012, 40, D115–D122. [Google Scholar] [CrossRef] [PubMed]
- Segata, N. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y. Controlling the false discovery rate in behavior genetics research. Behav. Brain Res. 2001, 125, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Kramer, A. Causal analysis approaches in ingenuity pathway analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Meehan, C.J.; Beiko, R.G. A phylogenomic view of ecological specialization in the Lachnospiraceae, a family of digestive tract-associated bacteria. Genome Biol. Evol. 2014, 6, 703–713. [Google Scholar] [CrossRef] [PubMed]
- Vital, M.; Howe, A.C.; Tiedje, J.M. Revealing the bacterial butyrate synthesis pathways by analyzing (meta)genomic data. MBio 2014, 5, e00889. [Google Scholar] [CrossRef] [PubMed]
- Hamer, H.M. Review article: The role of butyrate on colonic function. Aliment Pharmacol. Ther. 2008, 27, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Clemente, J.C. The impact of the gut microbiota on human health: An integrative view. Cell 2012, 148, 1258–1270. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.W. Dominant and diet-responsive groups of bacteria within the human colonic microbiota. ISME J. 2011, 5, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.H. Oscillibacter ruminantium sp. nov., isolated from the rumen of Korean native cattle. Int. J. Syst. Evol. Microbiol. 2013, 63, 1942–1946. [Google Scholar] [CrossRef] [PubMed]
- Myint, H. Functional modulation of caecal fermentation and microbiota in rat by feeding bean husk as a dietary fibre supplement. Benef. Microbes 2018, 1–12. [Google Scholar]
- Eeckhaut, V. Butyricicoccus pullicaecorum in inflammatory bowel disease. Gut 2013, 62, 1745–1752. [Google Scholar] [CrossRef] [PubMed]
- Le Floc’h, N. Impact of feed restriction on health, digestion and faecal microbiota of growing pigs housed in good or poor hygiene conditions. Animal 2014, 8, 1632–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devriese, S. Reduced mucosa-associated butyricicoccus activity in patients with ulcerative colitis correlates with aberrant claudin-1 expression. J. Crohns. Colitis. 2017, 11, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Chassard, C. Ruminococcus champanellensis sp. nov. a cellulose-degrading bacterium from human gut microbiota. Int. J. Syst. Evol. Microbiol. 2012, 62, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Kang, S. Dysbiosis of fecal microbiota in Crohn’s disease patients as revealed by a custom phylogenetic microarray. Inflamm. Bowel. Dis. 2010, 16, 2034–2042. [Google Scholar] [CrossRef] [PubMed]
- Prieto, I. Influence of a diet enriched with virgin olive oil or butter on mouse gut microbiota and its correlation to physiological and biochemical parameters related to metabolic syndrome. PLoS ONE 2018, 13, e0190368. [Google Scholar] [CrossRef] [PubMed]
- Togo, A.H. ‘Eisenbergiella massiliensis’, a new species isolated from human stool collected after bariatric surgery. New Microbes. New Infect. 2016, 13, 15–16. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.X. Alterations and correlations of the gut microbiome, metabolism and immunity in patients with primary biliary cirrhosis. Environ. Microbiol. 2016, 18, 2272–2286. [Google Scholar] [CrossRef] [PubMed]
- Baumler, A.J.; Sperandio, V. Interactions between the microbiota and pathogenic bacteria in the gut. Nature 2016, 535, 85–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Rodas, B. Microbiome profiling of commercial pigs from farrow to finish. J. Anim. Sci. 2018, 96, 1778–1794. [Google Scholar] [CrossRef] [PubMed]
- Kovatcheva-Datchary, P. Dietary fiber-induced improvement in glucose metabolism is associated with increased abundance of prevotella. Cell Metab. 2015, 22, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Duque, G.A.; Descoteaux, A. Macrophage cytokines: Involvement in immunity and infectious diseases. Front. Immunol. 2014, 5, 1–12. [Google Scholar]
- Elsayed, E.A. Mushrooms: A potential natural source of anti-inflammatory compounds for medical applications. Mediators Inflamm. 2014, 2014, 805841. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient | Per 100 g Fresh Mushroom |
|---|---|
| Protein, g/100 g | 2.72 |
| Carbohydrate, g/100 g | 3.35 |
| Fat, g/100 g | 0.36 |
| Ash, g/100 g | 0.84 |
| Moisture, g/100 g | 92.72 |
| Dietary Fiber, g/100 g | 2.39 |
| Calorie, kcal/100 g | 27.53 |
| Gene Symbol | Gene Name | Location | Type(s) | Control_LPS | 3-Serv_LPS | 6-Serv_LPS | Log Difference (Control-3-Serv) | Log Difference (Control-6-Serv) | Expression Effect | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Log Ratio | FDR (q Value) | Log Ratio | FDR (q Value) | Log Ratio | FDR (q Value) | |||||||
| RETN | Resistin | Extracellular space | Peptide hormone | 5.117 a | 3.70 × 10−5 | 8.485 | 1.00 × 10−4 | 6.284 | 2.81 × 10−5 | 3.37 | 1.17 | increased |
| APOA1 | Apolipoprotein A1 | Extracellular space | Lipoprotein | −5.845 | 3.28 × 10−5 | −3.988 | 9.46 × 10−5 | −4.057 | 4.70 × 10−5 | 1.86 | 1.79 | increased |
| CSF3 | Colony stimulating factor 3 | Cytoplasm | Growth factor | 8.816 | 3.20 × 10−6 | 7.058 | 2.00 × 10−5 | 6.982 | 4.97 × 10−6 | −1.758 | −1.834 | reduced |
| IL12B | Interleukin 12B | Plasma membrane | Cytokine | 8.486 | 6.51 × 10−4 | 5.527 | 2.57 × 10−3 | 5.944 | 2.68 × 10−4 | −2.959 | −2.542 | reduced |
| IL6 | Interleukin 6 | Extracellular space | Cytokine | 8.428 | 5.35 × 10−5 | 5.587 | 3.45 × 10−4 | 5.05 | 1.30 × 10−4 | −2.841 | −3.378 | reduced |
| IL1B | Interleukin 1 beta | Plasma membrane | Cytokine | 8.289 | 1.58 × 10−5 | 7.192 | 1.17 × 10−4 | 6.985 | 3.65 × 10−5 | −1.097 | −1.304 | reduced |
| CSF2 | Colony stimulating factor 2 | Plasma membrane | Growth factor | 7.537 | 4.60 × 10−5 | 5.661 | 2.04 × 10−4 | 5.344 | 4.28 × 10−5 | −1.876 | −2.193 | reduced |
| CXCL13 | C-X-C motif chemokine ligand 13 | Cytoplasm | Chemokine | 4.92 | 4.31 × 10−4 | 3.869 | 6.78 × 10−3 | 3.51 | 5.49 × 10−3 | −1.051 | −1.41 | reduced |
| IL1R1 | Interleukin 1 receptor type 1 | Plasma membrane | IL1 type receptor | 3.623 | 3.55 × 10−4 | 1.798 | 2.26 × 10−02 | 1.61 | 1.86 × 10−02 | −1.825 | −2.013 | reduced |
| IL1R2 | Interleukin 1 receptor type 2 | Cytoplasm | IL1 type receptor | 3.532 | 9.77 × 10−5 | 1.62 | 1.83 × 10−02 | 2.226 | 1.04 × 10−3 | −1.912 | −1.306 | reduced |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solano-Aguilar, G.I.; Jang, S.; Lakshman, S.; Gupta, R.; Beshah, E.; Sikaroodi, M.; Vinyard, B.; Molokin, A.; Gillevet, P.M.; Urban, J.F., Jr. The Effect of Dietary Mushroom Agaricus bisporus on Intestinal Microbiota Composition and Host Immunological Function. Nutrients 2018, 10, 1721. https://doi.org/10.3390/nu10111721
Solano-Aguilar GI, Jang S, Lakshman S, Gupta R, Beshah E, Sikaroodi M, Vinyard B, Molokin A, Gillevet PM, Urban JF Jr. The Effect of Dietary Mushroom Agaricus bisporus on Intestinal Microbiota Composition and Host Immunological Function. Nutrients. 2018; 10(11):1721. https://doi.org/10.3390/nu10111721
Chicago/Turabian StyleSolano-Aguilar, Gloria I., Saebyeol Jang, Sukla Lakshman, Richi Gupta, Ethiopia Beshah, Masoumeh Sikaroodi, Bryan Vinyard, Aleksey Molokin, Patrick M. Gillevet, and Joseph F. Urban, Jr. 2018. "The Effect of Dietary Mushroom Agaricus bisporus on Intestinal Microbiota Composition and Host Immunological Function" Nutrients 10, no. 11: 1721. https://doi.org/10.3390/nu10111721
APA StyleSolano-Aguilar, G. I., Jang, S., Lakshman, S., Gupta, R., Beshah, E., Sikaroodi, M., Vinyard, B., Molokin, A., Gillevet, P. M., & Urban, J. F., Jr. (2018). The Effect of Dietary Mushroom Agaricus bisporus on Intestinal Microbiota Composition and Host Immunological Function. Nutrients, 10(11), 1721. https://doi.org/10.3390/nu10111721