Chronic Consumption of Fructose Induces Behavioral Alterations by Increasing Orexin and Dopamine Levels in the Rat Brain

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Treatments
2.2. Blood Metabolic Parameters Analysis
2.3. Electrode Implantation
2.4. EEG Recordings, Sleep Stages and Spectral Power Analysis
2.5. Locomotor Activity
2.6. Enzyme-Linked Immunoassay (ELISA) Quantification of Orexin A
2.7. Determination of Monoamines by High-Performance Liquid Chromatography (HPLC)
2.8. Immunohistochemistry
2.9. Statistical Analysis
3. Results
3.1. Metabolic Parameters
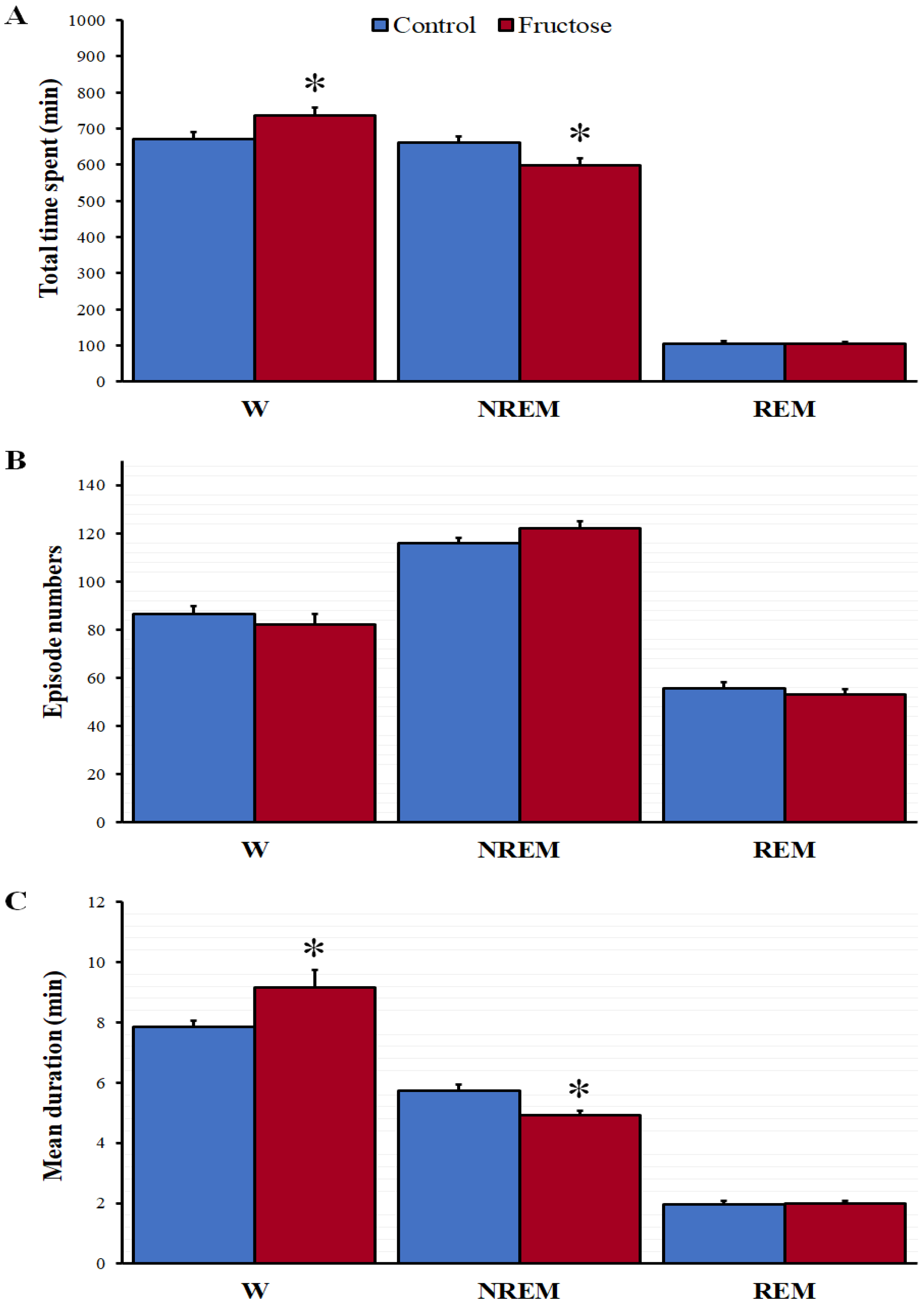
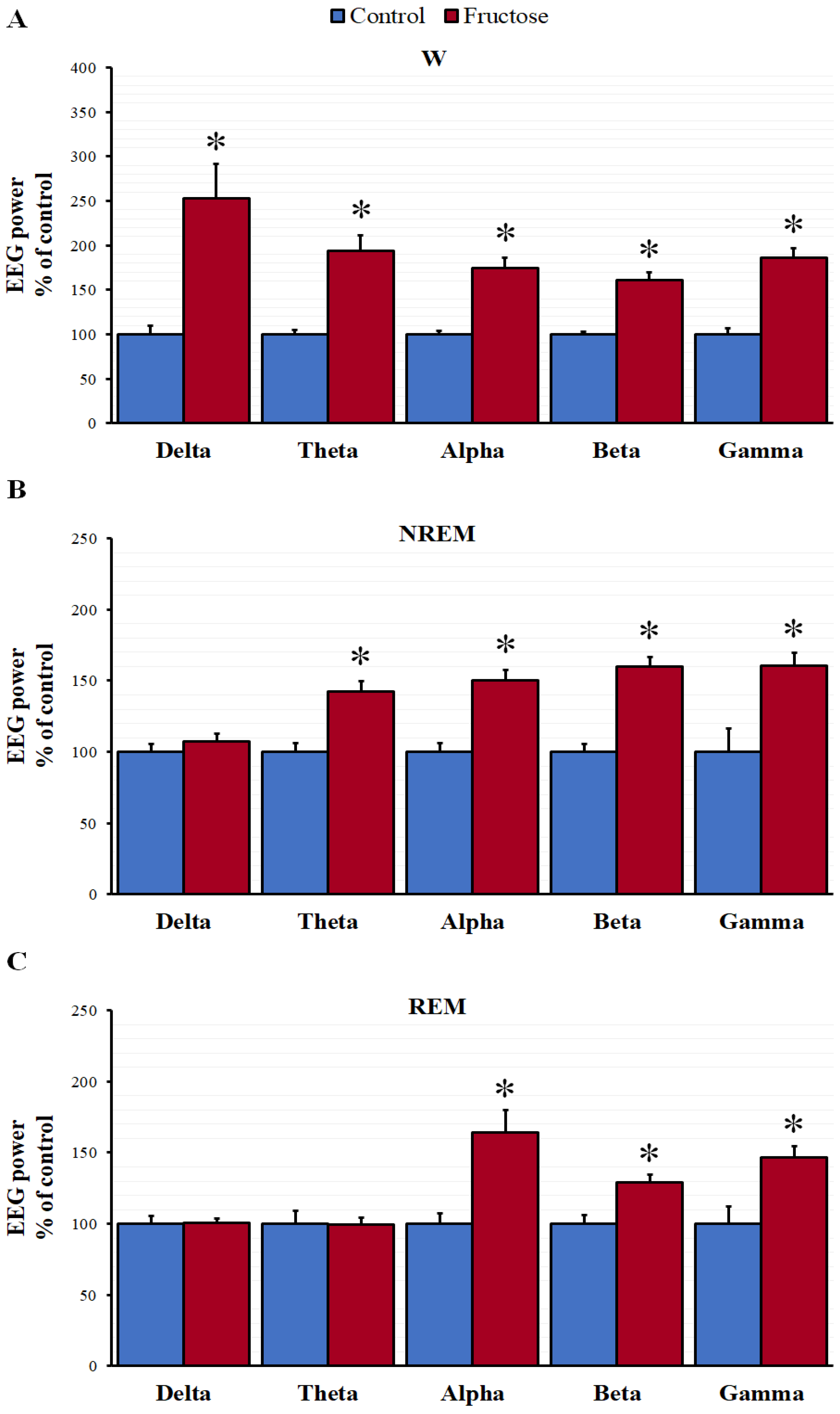
3.2. Sleep-Wake Cycle and EEG Spectral Power Analysis
3.3. Locomotor Activity
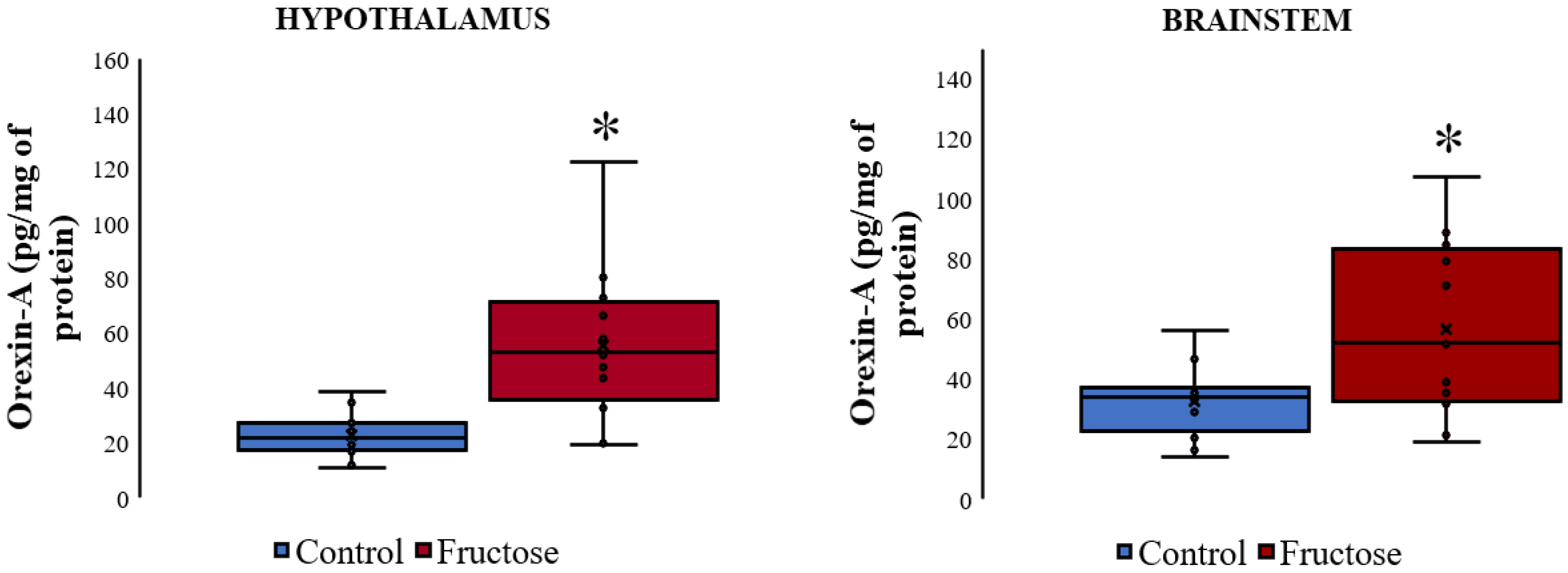
3.4. Orexin A Levels
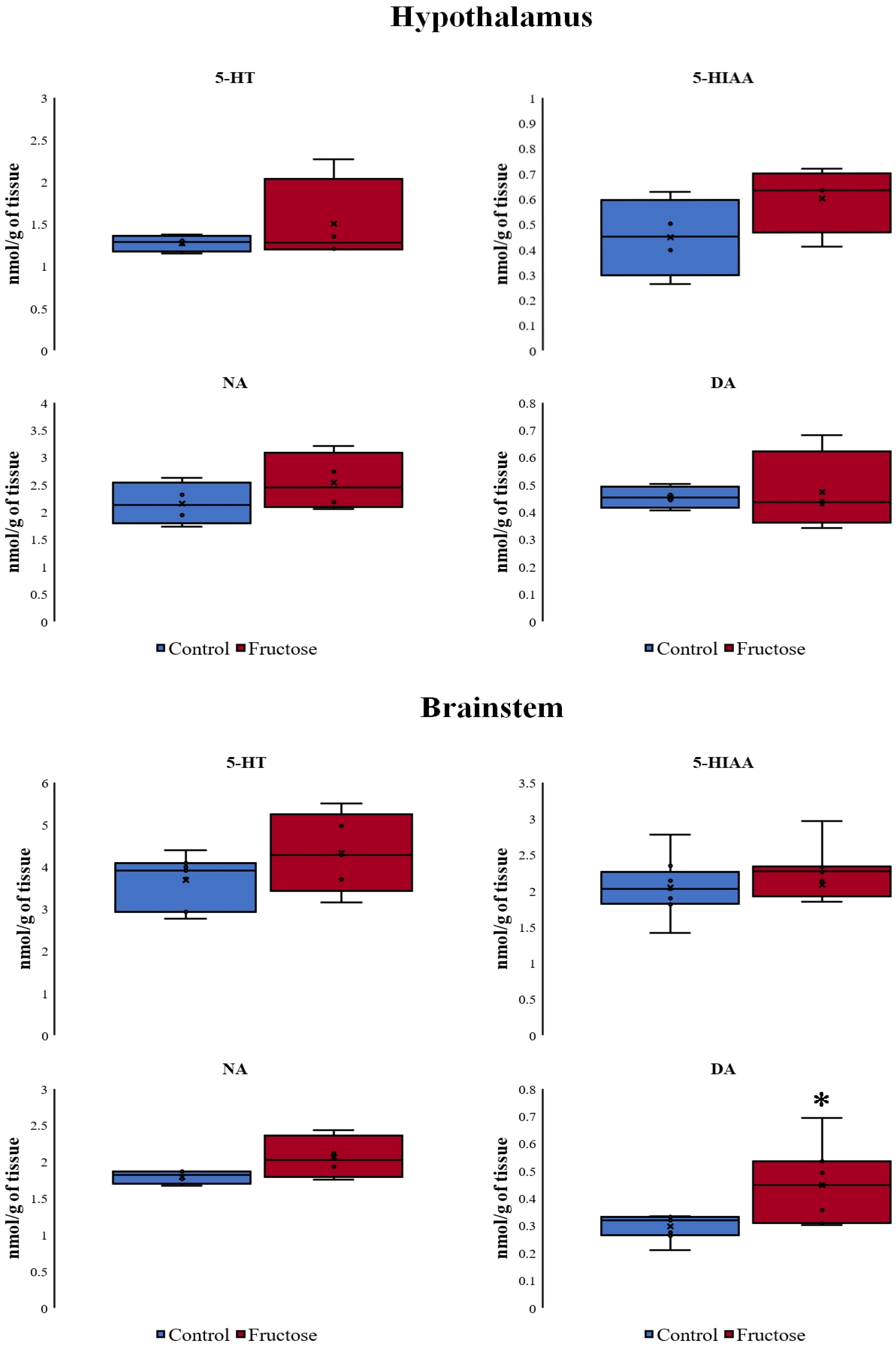
3.5. Concentration of Monoamines
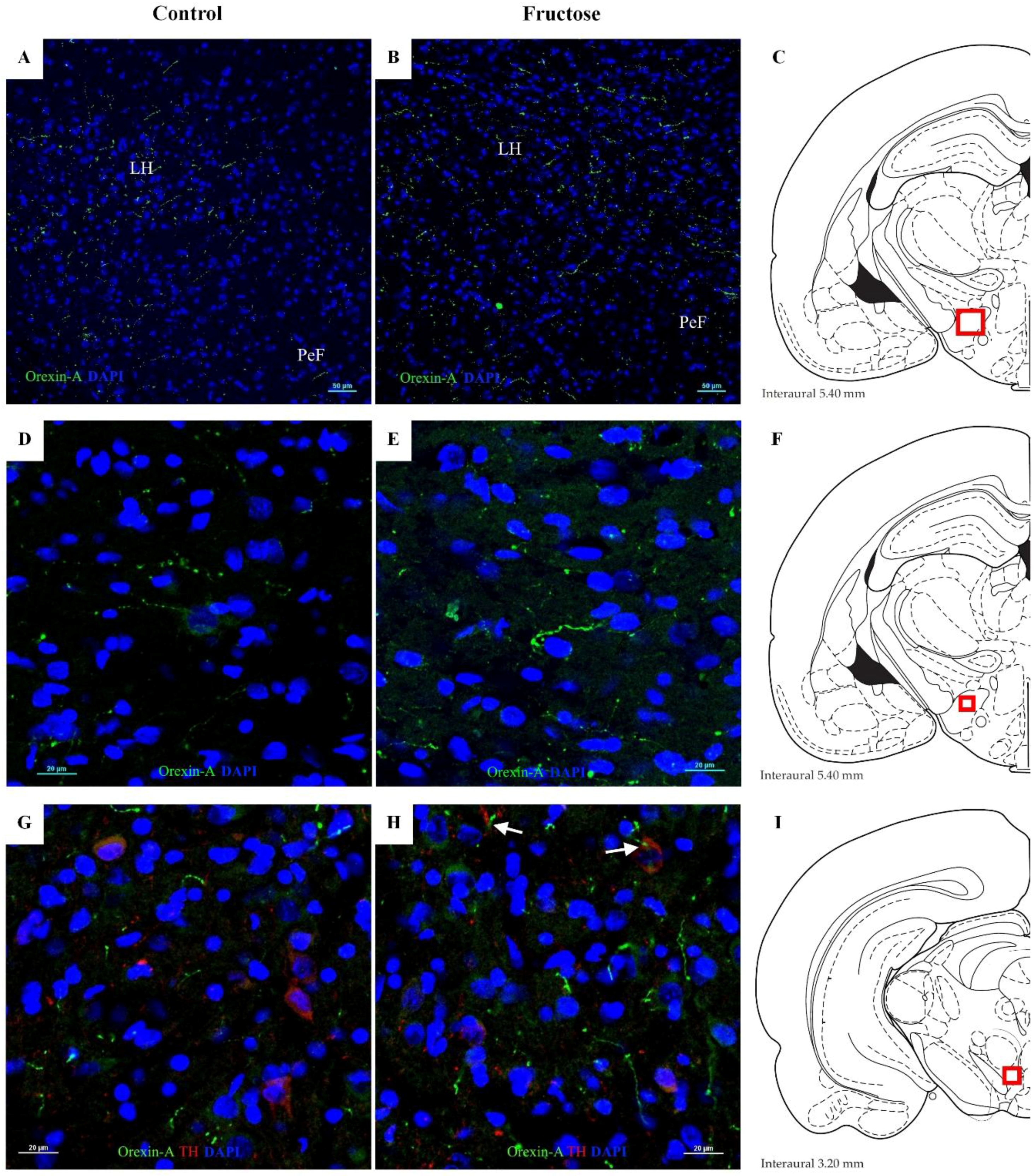
3.6. Immunohistochemical Findings
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Park, Y.K.; Yetley, E.A. Intakes and food sources of fructose in the United States. Am. J. Clin. Nutr. 1993, 58, 737S–747S. [Google Scholar] [CrossRef] [PubMed]
- Marriott, B.P.; Cole, N.; Lee, E. National estimates of dietary fructose intake increased from 1977 to 2004 in the United States. J. Nutr. 2009, 139, 1228S–1235S. [Google Scholar] [CrossRef] [PubMed]
- Stanhope, K.L. Role of fructose-containing sugars in the epidemics of obesity and metabolic syndrome. Annu. Rev. Med. 2012, 63, 329–343. [Google Scholar] [CrossRef] [PubMed]
- Bidwell, A.J. Chronic fructose ingestion as a major health concern: Is a sedentary lifestyle making it worse? A review. Nutrients 2017, 9, 549. [Google Scholar] [CrossRef] [PubMed]
- Egli, L.; Lecoultre, V.; Theytaz, F.; Campos, V.; Hodson, L.; Schneiter, P.; Mittendorfer, B.; Patterson, B.W.; Fielding, B.A.; Gerber, P.A.; et al. Exercise prevents fructose-induced hypertriglyceridemia in healthy young subjects. Diabetes 2013, 62, 2259–2265. [Google Scholar] [CrossRef] [PubMed]
- Jalal, D.I.; Smits, G.; Johnson, R.J.; Chonchol, M. Increased fructose associates with elevated blood pressure. J. Am. Soc. Nephrol. 2010, 21, 1543–1549. [Google Scholar] [CrossRef] [PubMed]
- Bernardes, N.; Ayyappan, P.; De Angelis, K.; Bagchi, A.; Akolkar, G.; da Silva Dias, D.; Belló-Klein, A.; Singal, P.K. Excessive consumption of fructose causes cardiometabolic dysfunctions through oxidative stress and inflammation. Can. J. Physiol. Pharmacol. 2017, 95, 1078–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, K.S.; Simon, M.C.; Strassburger, K.; Markgraf, D.F.; Buyken, A.E.; Szendroedi, J.; Müssig, K.; Roden, M.; GDS Group. Habitual fructose intake relates to insulin sensitivity and fatty liver index in recent-onset type 2 diabetes patients and individuals without diabetes. Nutrients 2018, 10, 774. [Google Scholar] [CrossRef] [PubMed]
- Stanhope, K.L.; Griffen, S.C.; Bremer, A.A.; Vink, R.G.; Schaefer, E.J.; Nakajima, K.; Schwarz, J.M.; Beysen, C.; Berglund, L.; Keim, N.L.; et al. Metabolic responses to prolonged consumption of glucose- and fructose-sweetened beverages are not associated with postprandial or 24-h glucose and insulin excursions. Am. J. Clin. Nutr. 2011, 94, 112–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dekker, M.J.; Su, Q.; Baker, C.; Rutledge, A.C.; Adeli, K. Fructose: A highly lipogenic nutrient implicated in insulin resistance, hepatic steatosis, and the metabolic syndrome. Am. J. Physiol. Endocrinol. Metab. 2010, 299, E685–E694. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.M.; El-Denshary, E.S.; Abdallah, D.M. Geraniol, alone and in combination with pioglitazone, ameliorates fructose-induced metabolic syndrome in rats via the modulation of both inflammatory and oxidative stress status. PLoS ONE 2015, 10, e0117516. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, F.; Senese, R.; Lasala, P.; Ziello, A.; Mazzoli, A.; Crescenzo, R.; Liverini, G.; Lanni, A.; Goglia, F.; Iossa, S. Fructose-rich diet affects mitochondrial DNA damage and repair in rats. Nutrients 2017, 9, 323. [Google Scholar] [CrossRef] [PubMed]
- Van der Borght, K.; Köhnke, R.; Göransson, N.; Deierborg, T.; Brundin, P.; Erlanson-Albertsson, C.; Lindqvist, A. Reduced neurogenesis in the rat hippocampus following high fructose consumption. Regul. Pept. 2011, 167, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Bursać, B.N.; Vasiljević, A.D.; Nestorović, N.M.; Veličković, N.A.; Vojnović-Milutinović, D.D.; Matić, G.M.; Djordjevic, A.D. High-fructose diet leads to visceral adiposity and hypothalamic leptin resistance in male rats—Do glucocorticoids play a role? J. Nutr. Biochem. 2014, 25, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Yin, Q.; Ma, Y.; Hong, Y.; Hou, X.; Chen, J.; Shen, C.; Sun, M.; Shang, Y.; Dong, S.; Zeng, Z.; et al. Lycopene attenuates insulin signaling deficits, oxidative stress, neuroinflammation, and cognitive impairment in fructose-drinking insulin resistant rats. Neuropharmacology 2014, 86, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Spagnuolo, M.S.; Bergamo, P.; Crescenzo, R.; Iannotta, L.; Treppiccione, L.; Iossa, S.; Cigliano, L. Brain Nrf2 pathway, autophagy, and synaptic function proteins are modulated by a short-term fructose feeding in young and adult rats. Nutr. Neurosci. 2018, 24, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Maldonado, A.; Ying, Z.; Byun, H.R.; Gomez-Pinilla, F. Short-term fructose ingestion affects the brain independently from establishment of metabolic syndrome. Biochim. Biophys. Acta 2018, 1864, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Cigliano, L.; Spagnuolo, M.S.; Crescenzo, R.; Cancelliere, R.; Iannotta, L.; Mazzoli, A.; Liverini, G.; Iossa, S. Short-term fructose feeding induces inflammation and oxidative stress in the hippocampus of young and adult rats. Mol. Neurobiol. 2018, 55, 2869–2883. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.P.; Bartness, T.J.; Mielke, J.G.; Parent, M.B. A high fructose diet impairs spatial memory in male rats. Neurobiol. Learn. Mem. 2009, 92, 410–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sangüesa, G.; Cascales, M.; Griñán, C.; Sánchez, R.M.; Roglans, N.; Pallàs, M.; Laguna, J.C.; Alegret, M. Impairment of novel object recognition memory and brain insulin signaling in fructose- but not glucose-drinking female rats. Mol. Neurobiol. 2018, 55, 6984–6999. [Google Scholar] [CrossRef] [PubMed]
- Killgore, W.D. Effects of sleep deprivation on cognition. Prog. Brain Res. 2010, 185, 105–129. [Google Scholar] [CrossRef] [PubMed]
- McCoy, J.G.; Strecker, R.E. The cognitive cost of sleep lost. Neurobiol. Learn. Mem. 2011, 96, 564–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco-Pérez, J.; Ballesteros-Zebadúa, P.; Custodio, V.; Paz, C. Major neurotransmitters involved in the regulation of sleep-wake cycle. Rev. Investig. Clin. 2012, 64, 182–191. [Google Scholar]
- Ikemoto, S. Dopamine reward circuitry: Two projection systems from the ventral midbrain to the nucleus accumbens-olfactory tubercle complex. Brain Res. Rev. 2007, 56, 27–78. [Google Scholar] [CrossRef] [PubMed]
- Meye, F.J.; Adan, R.A. Feelings about food: The ventral tegmental area in food reward and emotional eating. Trends Pharmacol. Sci. 2014, 35, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Oishi, Y.; Lazarus, M. The control of sleep and wakefulness by mesolimbic dopamine systems. Neurosci. Res. 2017, 118, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Korotkova, T.M.; Sergeeva, O.A.; Eriksson, K.S.; Haas, H.L.; Brown, R.E. Excitation of ventral tegmental area dopaminergic and nondopaminergic neurons by orexins/hypocretins. J. Neurosci. 2003, 23, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; McNeill, J.H. Fructose-induced hypertension in rats is concentration- and duration-dependent. J. Pharmacol. Toxicol. Methods 1995, 33, 101–107. [Google Scholar] [CrossRef]
- Ballesteros-Zebadua, P.; Custodio, V.; Franco-Perez, J.; Rubio, C.; González, E.; Trejo, C.; Celis, M.A.; Paz, C. Whole-brain irradiation increases NREM sleep and hypothalamic expression of IL-1β in rats. Int. J. Radiat. Biol. 2014, 90, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Franco-Pérez, J.; Paz, C. Quinine, a selective gap junction blocker, decreases REM sleep in rats. Pharmacol. Biochem. Behav. 2009, 94, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Vidal, Y.; Monroy-Noyola, A.; Anaya-Ramos, L.; Arteaga-Silva, M.; Mendez-Armenta, M.; Ostoa-Saloma, P.; Díaz-Zaragoza, M.; Morales-Montor, J.; Ríos, C.; Montes, S. β-Estradiol-3-benzoate confers neuroprotection in Parkinson MPP+ rat model through inhibition of lipid peroxidation. Steroids 2017, 126, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 3rd ed.; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Dos Santos, F.; Moraes-Silva, I.C.; Moreira, E.D.; Irigoyen, M.C. The role of the baroreflex and parasympathetic nervous system in fructose-induced cardiac and metabolic alterations. Sci. Rep. 2018, 8, 10970. [Google Scholar] [CrossRef] [PubMed]
- Olaniyi, K.S.; Olatunji, L.A. Oral ethinylestradiol-levonorgestrel attenuates cardiac glycogen and triglyceride accumulation in high fructose female rats by suppressing pyruvate dehydrogenase kinase-4. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Rasch, B.; Born, J. About sleep’s role in memory. Physiol. Rev. 2013, 93, 681–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, S.; Mavanji, V.; Ulloor, J.; Patterson, E.H. Activation of phasic pontine-wave generator prevents rapid eye movement sleep deprivation-induced learning impairment in the rat: A mechanism for sleep-dependent plasticity. J. Neurosci. 2004, 24, 1416–1427. [Google Scholar] [CrossRef] [PubMed]
- Aleisa, A.M.; Helal, G.; Alhaider, I.A.; Alzoubi, K.H.; Srivareerat, M.; Tran, T.T.; Al-Rejaie, S.S.; Alkadhi, K.A. Acute nicotine treatment prevents REM sleep deprivation-induced learning and memory impairment in rat. Hippocampus 2011, 21, 899–909. [Google Scholar] [CrossRef] [PubMed]
- Ravassard, P.; Hamieh, A.M.; Joseph, M.A.; Fraize, N.; Libourel, P.A.; Lebarillier, L.; Arthaud, S.; Meissirel, C.; Touret, M.; Malleret, G.; et al. REM sleep-dependent bidirectional regulation of hippocampal-based emotional memory and LTP. Cereb. Cortex 2016, 26, 1488–1500. [Google Scholar] [CrossRef] [PubMed]
- Born, J. Slow-wave sleep and the consolidation of long-term memory. World J. Biol. Psychiatry 2010, 11, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Oyanedel, C.N.; Binder, S.; Kelemen, E.; Petersen, K.; Born, J.; Inostroza, M. Role of slow oscillatory activity and slow wave sleep in consolidation of episodic-like memory in rats. Behav. Brain Res. 2014, 275, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Colavito, V.; Fabene, P.F.; Grassi-Zucconi, G.; Pifferi, F.; Lamberty, Y.; Bentivoglio, M.; Bertini, G. Experimental sleep deprivation as a tool to test memory deficits in rodents. Front. Syst. Neurosci. 2013, 7, 106. [Google Scholar] [CrossRef] [PubMed]
- Hairston, I.S.; Little, M.T.; Scanlon, M.D.; Barakat, M.T.; Palmer, T.D.; Sapolsky, R.M.; Heller, H.C. Sleep restriction suppresses neurogenesis induced by hippocampus-dependent learning. J. Neurophysiol. 2005, 94, 4224–4233. [Google Scholar] [CrossRef] [PubMed]
- Inostroza, M.; Binder, S.; Born, J. Sleep-dependency of episodic-like memory consolidation in rats. Behav. Brain Res. 2013, 237, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Ohno, K.; Sakurai, T. Orexin neuronal circuitry: Role in the regulation of sleep and wakefulness. Front. Neuroendocrinol. 2008, 29, 70–87. [Google Scholar] [CrossRef] [PubMed]
- Rorabaugh, J.M.; Stratford, J.M.; Zahniser, N.R. A relationship between reduced nucleus accumbens shell and enhanced lateral hypothalamic orexin neuronal activation in long-term fructose bingeing behavior. PLoS ONE 2014, 9, e95019. [Google Scholar] [CrossRef] [PubMed]
- Fadel, J.; Deutch, A.Y. Anatomical substrates of orexin-dopamine interactions: Lateral hypothalamic projections to the ventral tegmental area. Neuroscience 2002, 111, 379–387. [Google Scholar] [CrossRef]
- Gruner, J.A.; Marcy, V.R.; Lin, Y.G.; Bozyczko-Coyne, D.; Marino, M.J.; Gasior, M. The roles of dopamine transport inhibition and dopamine release facilitation in wake enhancement and rebound hypersomnolence induced by dopaminergic agents. Sleep 2009, 32, 1425–1438. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, S.; Nakamichi, K.; Imai, H.; Ichihara, J. Effect of dopamine D4 receptor agonists on sleep architecture in rats. Prog. Neuropsychopharmacol. Biol. Psychiatry 2015, 63, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Vyazovskiy, V.V.; Tobler, I. Theta activity in the waking EEG is a marker of sleep propensity in the rat. Brain Res. 2005, 1050, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Montaron, M.F.; Bouyer, J.J.; Rougeul, A.; Buser, P. Ventral mesencephalic tegmentum (VMT) controls electrocortical beta rhythms and associated attentive behaviour in the cat. Behav. Brain Res. 1982, 6, 129–145. [Google Scholar] [CrossRef]
- Orzeł-Gryglewska, J.; Matulewicz, P.; Jurkowlaniec, E. Brainstem system of hippocampal theta induction: The role of the ventral tegmental area. Synapse 2015, 69, 553–575. [Google Scholar] [CrossRef] [PubMed]
- Gerashchenko, D.; Salin-Pascual, R.; Shiromani, P.J. Effects of hypocretin-saporin injections into the medial septum on sleep and hippocampal theta. Brain Res. 2001, 913, 106–115. [Google Scholar] [CrossRef]
- Vassalli, A.; Franken, P. Hypocretin (orexin) is critical in sustaining theta/gamma-rich waking behaviors that drive sleep need. Proc. Natl. Acad. Sci. USA 2017, 114, E5464–E5473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, J.A.; Jensen, L.T.; Fugger, L.; Burdakov, D. Metabolism-independent sugar sensing in central orexin neurons. Diabetes 2008, 57, 2569–2576. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.Q.; Karatayev, O.; Davydova, Z.; Leibowitz, S.F. Circulating triglycerides impact on orexigenic peptides and neuronal activity in hypothalamus. Endocrinology 2004, 145, 3904–3912. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Fructose | Significance | |
|---|---|---|---|
| Weight gain (g) | 119.7 ± 3.3 | 106.2 ± 6.5 | ns |
| Glucose (mg/dL) | 111.5 ± 1.7 | 126.7 ± 1.5 * | p = 0.001 |
| Triglycerides (mg/dL) | 68.8 ± 6.8 | 116.4 ± 15.6 * | p = 0.015 |
| Cholesterol (mg/dL) | 41.5 ± 1.8 | 52.8 ± 3.3 * | p = 0.011 |
| ALT (U/L) | 83.4 ± 6.0 | 68.6 ± 4.8 | ns |
| AST (U/L) | 220.0 ± 17.3 | 197.0 ± 11.4 | ns |
| ALP (U/L) | 194.9 ± 18.2 | 161.1 ± 20.4 | ns |
| LDH (U/L) | 1554.9 ± 123.8 | 1522.6 ± 149.2 | ns |
| Urea (mg/dL) | 38.1 ± 1.5 | 38.3 ± 4.4 | ns |
| Creatinine (mg/dL) | 0.34 ± 0.01 | 0.35 ± 0.02 | ns |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franco-Pérez, J.; Manjarrez-Marmolejo, J.; Ballesteros-Zebadúa, P.; Neri-Santos, A.; Montes, S.; Suarez-Rivera, N.; Hernández-Cerón, M.; Pérez-Koldenkova, V. Chronic Consumption of Fructose Induces Behavioral Alterations by Increasing Orexin and Dopamine Levels in the Rat Brain. Nutrients 2018, 10, 1722. https://doi.org/10.3390/nu10111722
Franco-Pérez J, Manjarrez-Marmolejo J, Ballesteros-Zebadúa P, Neri-Santos A, Montes S, Suarez-Rivera N, Hernández-Cerón M, Pérez-Koldenkova V. Chronic Consumption of Fructose Induces Behavioral Alterations by Increasing Orexin and Dopamine Levels in the Rat Brain. Nutrients. 2018; 10(11):1722. https://doi.org/10.3390/nu10111722
Chicago/Turabian StyleFranco-Pérez, Javier, Joaquín Manjarrez-Marmolejo, Paola Ballesteros-Zebadúa, Adriana Neri-Santos, Sergio Montes, Norma Suarez-Rivera, Miguel Hernández-Cerón, and Vadim Pérez-Koldenkova. 2018. "Chronic Consumption of Fructose Induces Behavioral Alterations by Increasing Orexin and Dopamine Levels in the Rat Brain" Nutrients 10, no. 11: 1722. https://doi.org/10.3390/nu10111722
APA StyleFranco-Pérez, J., Manjarrez-Marmolejo, J., Ballesteros-Zebadúa, P., Neri-Santos, A., Montes, S., Suarez-Rivera, N., Hernández-Cerón, M., & Pérez-Koldenkova, V. (2018). Chronic Consumption of Fructose Induces Behavioral Alterations by Increasing Orexin and Dopamine Levels in the Rat Brain. Nutrients, 10(11), 1722. https://doi.org/10.3390/nu10111722