Evidence of Some Natural Products with Antigenotoxic Effects. Part 2: Plants, Vegetables, and Natural Resin

 ,
,  , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Antigenotoxic Plants and Vegetables
2.1. Blue Green Algae Spirulina (Arthrospira maxima and Arthrospira Platensis)
2.2. Broccoli (Brassica Oleracea Itálica)
2.3. Chamomile (Matricaria chamomilla or Chamomilla Recutita)
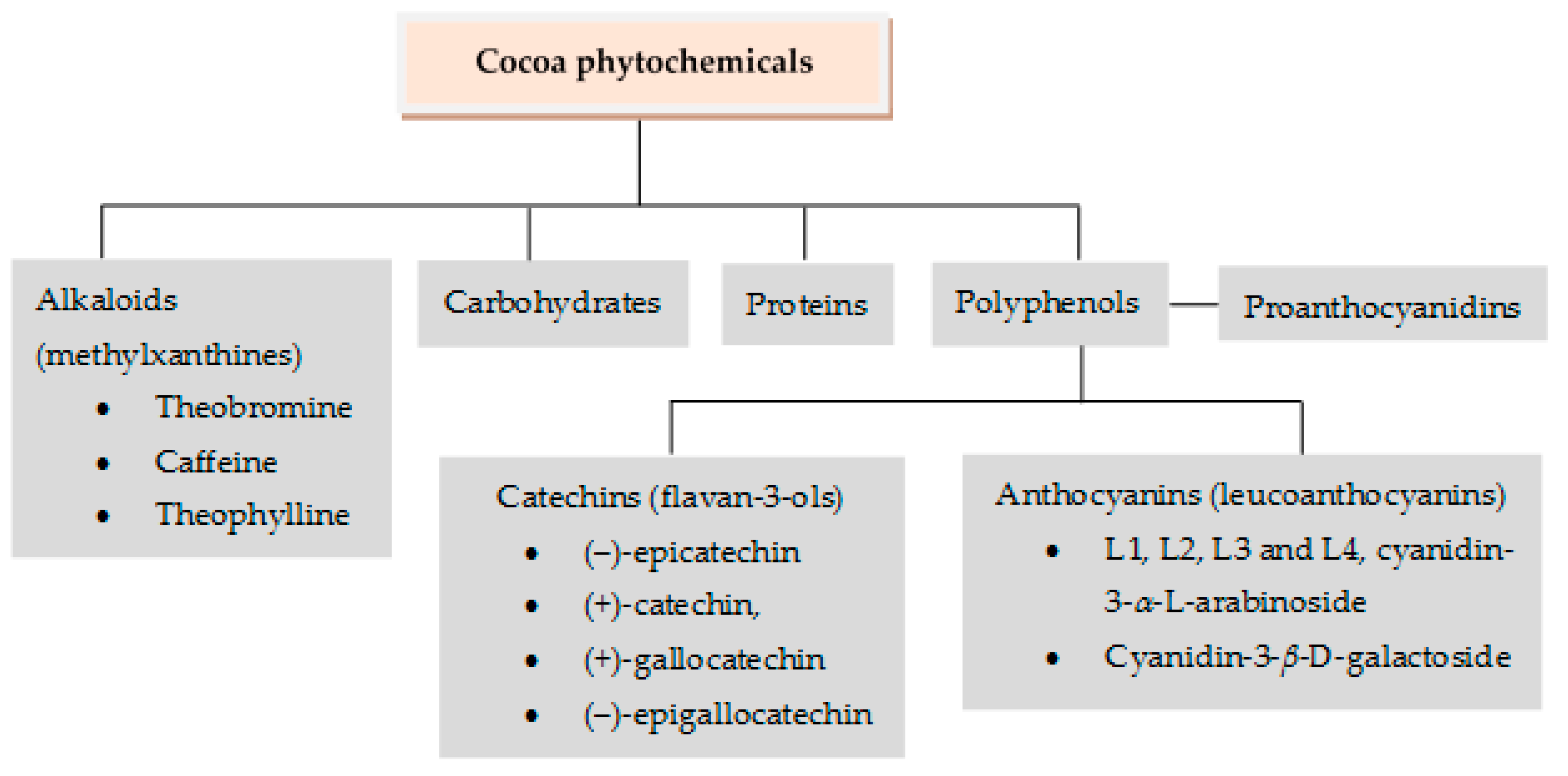
2.4. Cocoa (Theobroma cacao L.)
2.5. Ginger (Zingiber Officinale)
2.6. Bay Laurel (Laurus nobilis L.)
2.7. Marigold (Calendula Officinalis Linn.)
2.8. Roselle (Hibiscus Sabdariffa Linn.)
2.9. Rosemary (Rosmarinus Officinalis L.)
3. Natural Resin
3.1. Propolis (Bee Glue)
3.2. Antigenotoxic Biomarkers in Cancer
4. Conclusions
Author Contributions
Funding
Note
Conflicts of Interest
References
- Bhattachar, S. Natural antimutagens: A review. Res. J. Med. Plant 2011, 5, 116–126. [Google Scholar] [CrossRef]
- Nagarathna, P.; Johnson Wesley, M.; Sriram Reddy, P.; Reena, K. Review on genotoxicity, its molecular mechanisms and prevention. Int. J. Pharm. Sci. Rev. Res. 2013, 22, 236–243. [Google Scholar]
- Słoczyńska, K.; Powroźnik, B.; Pękala, E.; Waszkielewicz, A.M. Antimutagenic compounds and their possible mechanisms of action. J. Appl. Genet. 2014, 55, 273–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferguson, L.R.; Philpott, M. Nutrition and mutagenesis. Annu. Rev. Nutr. 2008, 28, 313–329. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, L.R. Natural and human-made mutagens and carcinogens in the human diet. Toxicology 2002, 181–182, 79–82. [Google Scholar] [CrossRef]
- Teaf, C.M.; Middendorf, P.J. Mutagenesis and genetic toxicology. In Principles of Toxicology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2000; pp. 239–264. [Google Scholar]
- De Flora, S.; Ferguson, L.R. Overview of mechanisms of cancer chemopreventive agents. Mutat. Res. 2005, 591, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, L.R.; Bronzetti, G.; De Flora, S. Mechanistic approaches to chemoprevention of mutation and cancer. Mutat. Res. 2005, 591, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.H. Potential synergy of phytochemicals in cancer prevention: Mechanism of action. J. Nutr. 2004, 134, S3479–S3485. [Google Scholar] [CrossRef]
- Ferguson, L.R.; Philpott, M.; Karunasinghe, N. Dietary cancer and prevention using antimutagens. Toxicology 2004, 198, 147–159. [Google Scholar] [CrossRef]
- Brusick, D. Evolution of testing strategies for genetic toxicity. Mutat. Res. 1988, 205, 69–78. [Google Scholar] [CrossRef]
- Abilev, S.K.; Glaser, V.M. Genetic toxicology: Findings and challenges. Russ. J. Genet. 2013, 49, 70–80. [Google Scholar] [CrossRef]
- Marzin, D. Theory and practice of genetic toxicology tests. Tests on eukaryotes. Ann. Biol. Clin. 1986, 44, 656–661. [Google Scholar]
- Mortelmans, K.; Zeiger, E. The Ames Salmonella/microsome mutagenicity assay. Mutat. Res. 2000, 455, 29–60. [Google Scholar] [CrossRef]
- Costa, S.; Carvalho, S.; Costa, C.; Coelho, P.; Silva, S.; Santos, L.S.; Gaspar, J.F.; Porto, B.; Laffon, B.; Teixeira, J.P. Increased levels of chromosomal aberrations and DNA damage in a group of workers exposed to formaldehyde. Mutagenesis 2015, 30, 463–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majer, B.J.; Laky, B.; Knasmüller, S.; Kassie, F. Use of the micronucleus assay with exfoliated epithelial cells as a biomarker for monitoring individuals at elevated risk of genetic damage and in chemoprevention trials. Mutat. Res. 2001, 489, 147–172. [Google Scholar] [CrossRef]
- Liao, W.; McNutt, M.A.; Zhu, W.-G. The comet assay: A sensitive method for detecting DNA damage in individual cells. Methods 2009, 48, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Madrigal-Santillán, E.; Madrigal-Bujaidar, E.; Álvarez-González, I.; Sumaya-Martínez, M.T.; Gutiérrez-Salinas, J.; Bautista, M.; Morales-González, Á.; García-Luna González-Rubio, M.; Aguilar-Faisal, J.L.; Morales-González, J.A. Review of natural products with hepatoprotective effects. World J. Gastroenterol. 2014, 20, 14787–14804. [Google Scholar] [CrossRef]
- Patel, S.; Goyal, A. Current and prospective insights on food and pharmaceutical applications of spirulina. Curr. Trends Biotechnol. Pharm. 2013, 7, 681–695. [Google Scholar]
- Nicoletti, M. Microalgae nutraceuticals. Foods Basel Switz. 2016, 5, 54. [Google Scholar] [CrossRef]
- Deng, R.; Chow, T.-J. Hypolipidemic, Antioxidant, and antiinflammatory activities of microalgae spirulina. Cardiovasc. Ther. 2010, 28, e33–e45. [Google Scholar] [CrossRef]
- Oliveira, E.G.; Rosa, G.S.; Moraes, M.A.; Pinto, L.A.A. Characterization of thin layer drying of Spirulina platensis utilizing perpendicular air flow. Bioresour. Technol. 2009, 100, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Qishen, P.; Guo, B.J.; Kolman, A. Radioprotective effect of extract from Spirulina platensis in mouse bone marrow cells studied by using the micronucleus test. Toxicol. Lett. 1989, 48, 165–169. [Google Scholar] [CrossRef]
- Premkumar, K.; Pachiappan, A.; Abraham, S.K.; Santhiya, S.T.; Gopinath, P.M.; Ramesh, A. Effect of Spirulina fusiformis on cyclophosphamide and mitomycin-C induced genotoxicity and oxidative stress in mice. Fitoterapia 2001, 72, 906–911. [Google Scholar] [CrossRef]
- Ruiz Flores, L.E.; Madrigal-Bujaidar, E.; Salazar, M.; Chamorro, G. Anticlastogenic effect of Spirulina maxima extract on the micronuclei induced by maleic hydrazide in Tradescantia. Life Sci. 2003, 72, 1345–1351. [Google Scholar] [CrossRef]
- Kim, M.Y.; Cheong, S.H.; Lee, J.H.; Kim, M.J.; Sok, D.E.; Kim, M.R. Spirulina improves antioxidant status by reducing oxidative stress in rabbits fed a high-cholesterol diet. J. Med. Food 2010, 13, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.M.; Abdel-Aziem, S.H.; Abdel-Wahhab, M.A. Modulation of DNA damage and alteration of gene expression during aflatoxicosis via dietary supplementation of Spirulina (Arthrospira) and whey protein concentrate. Ecotoxicol. Environ. Saf. 2012, 79, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, W.A.; Ismail, S.A.; El-Hakim, Y.M.A. Spirulina platensis ameliorative effect against GSM 900-MHz cellular phone radiation-induced genotoxicity in male Sprague-Dawley rats. Comp. Clin. Pathol. 2014, 23, 1719–1726. [Google Scholar] [CrossRef]
- Chamorro-Cevallos, G.; Garduño-Siciliano, L.; Barrón, B.L.; Madrigal-Bujaidar, E.; Cruz-Vega, D.E.; Pages, N. Chemoprotective effect of Spirulina (Arthrospira) against cyclophosphamide-induced mutagenicity in mice. Food Chem. Toxicol. 2008, 46, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Chamorro-Cevallos, G.; Garduño-Siciliano, L.; Martínez-Galero, E.; Mojica-Villegas, A.; Pages, N.; Gutiérrez-Salmeán, G. The protective effect of dietary Arthrospira (Spirulina) maxima against mutagenicity induced by benzo[alpha]pyrene in mice. J. Med. Food 2014, 17, 527–534. [Google Scholar] [CrossRef]
- Chen, T.; Wong, Y.-S. In vitro antioxidant and antiproliferative activities of selenium-containing phycocyanin from selenium-enriched Spirulina platensis. J. Agric. Food Chem. 2008, 56, 4352–4358. [Google Scholar] [CrossRef]
- Álvarez-González, I.; Vázquez-Sánchez, J.; Chamorro-Cevallos, G.; Madrigal-Bujaidar, E. Effect of Spirulina maxima and its protein extract on micronuclei induction by hydroxyurea in pregnant mice and their fetuses. J. Med. Food 2013, 16, 992–996. [Google Scholar] [CrossRef] [PubMed]
- Latté, K.P.; Appel, K.-E.; Lampen, A. Health benefits and possible risks of broccoli—An overview. Food Chem. Toxicol. 2011, 49, 3287–3309. [Google Scholar] [CrossRef]
- Food and Drug Administration (FDA), Center for Food Safety and Applied Nutrition (CFSAN). A Food Labeling Guide. Available online: https://www.fda.gov/downloads/food/guidance%20complianceregulatoryinformation/%20guidancedocuments/foodlabelingnutrition/foodlabelingguide/ucm265446.pdf (accessed on 16 July 2018).
- USDA. Food Composition Databases. Available online: https://ndb.nal.usda.gov/ndb/ (accessed on 22 June 2018).
- Owis, A.I. Broccoli; The green beauty: A Review. J. Pharm. Sci. Res. 2015, 7, 696–703. [Google Scholar]
- Kapusta-Duch, J.; Kopeć, A.; Piatkowska, E.; Borczak, B.; Leszczyńska, T. The beneficial effects of Brassica vegetables on human health. Rocz. Panstw. Zakl. Hig. 2012, 63, 389–395. [Google Scholar] [PubMed]
- Slavin, J.L.; Lloyd, B. Health benefits of fruits and vegetables. Adv. Nutr. 2012, 3, 506–516. [Google Scholar] [CrossRef]
- Moreno, D.A.; Carvajal, M.; López-Berenguer, C.; García-Viguera, C. Chemical and biological characterisation of nutraceutical compounds of broccoli. J. Pharm. Biomed. Anal. 2006, 41, 1508–1522. [Google Scholar] [CrossRef]
- Conzatti, A.; Telles da Silva Fróes, F.C.; Schweigert Perry, I.D.; de Souza, C.G. Clinical and molecular evidence of the consumption of broccoli, glucoraphanin and sulforaphane in humans. Nutr. Hosp. 2014, 31, 559–569. [Google Scholar] [PubMed]
- Barcelo, S.; Gardiner, J.M.; Gescher, A.; Chipman, J.K. CYP2E1-mediated mechanism of anti-genotoxicity of the broccoli constituent sulforaphane. Carcinogenesis 1996, 17, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Shishu, I.P. Inhibition of cooked food-induced mutagenesis by dietary constituents: Comparison of two natural isothiocyanates. Food Chem. 2009, 112, 977–981. [Google Scholar] [CrossRef]
- Singla, A.K.; Kaur, I.P. Inhibition of mutagenicity of food-derived heterocyclic amines by sulforaphane—A constituent of broccoli. Indian J. Exp. Biol. 2003, 41, 216–219. [Google Scholar]
- Barcelo, S.; Mace, K.; Pfeifer, A.M.A.; Chipman, J.K. Production of DNA strand breaks by N-nitrosodimethylamine and w x 2-amino-3-methylimidazo 4,5-f quinoline in THLE cells expressing human CYP isoenzymes and inhibition by sulforaphane. Mutat. Res. 1998, 402, 111–120. [Google Scholar] [CrossRef]
- Bonnesen, C.; Eggleston, I.M.; Hayes, J.D. Dietary indoles and isothiocyanates that are generated from cruciferous vegetables can both stimulate apoptosis and confer protection against DNA damage in human colon cell lines. Cancer Res. 2001, 61, 6120–6130. [Google Scholar] [PubMed]
- Chakraborty, S.; Roy, M.; Bhattacharya, R.K. Prevention and repair of DNA damage by selected phytochemicals as measured by single cell gel electrophoresis. J. Environ. Pathol. Toxicol. Oncol. 2004, 23, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Arranz, N.; Haza, A.I.; García, A.; Möller, L.; Rafter, J.; Morales, P. Protective effects of isothiocyanates towards N-nitrosamine-induced DNA damage in the single-cell gel electrophoresis (SCGE)/HepG2 assay. J. Appl. Toxicol. 2006, 26, 466–473. [Google Scholar] [CrossRef] [PubMed]
- García, A.; Haza, A.I.; Arranz, N.; Rafter, J.; Morales, P. Protective effects of isothiocyanates alone or in combination with vitamin C towardsN-nitrosodibutylamine orN-nitrosopiperidine-induced oxidative DNA damage in the single-cell gel electrophoresis (SCGE)/HepG2 assay. J. Appl. Toxicol. 2008, 28, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Topè, A.M.; Rogers, P.F. Evaluation of protective effects of sulforaphane on DNA damage caused by exposure to low levels of pesticide mixture using comet assay. J. Environ. Sci. Health Part B 2009, 44, 657–662. [Google Scholar] [CrossRef]
- Liu, H.; Smith, A.J.O.; Lott, M.C.; Bao, Y.; Bowater, R.P.; Reddan, J.R.; Wormstone, I.M. Sulforaphane can protect lens cells against oxidative stress: Implications for cataract prevention. Investig. Opthalmology Vis. Sci. 2013, 54, 5236. [Google Scholar] [CrossRef]
- Agrawal, R.C.; Mehrotra, N.K. Assessment of mutagenic potential of propoxur and its modulation by indole-3-carbinol. Food Chem. Toxicol 1997, 35, 1081–1084. [Google Scholar] [CrossRef]
- Lai, C.-N.; Butler, M.A.; Matney, T.S. Antimutagenic activities of common vegetables and their chlorophyll content. Mutat. Res. Toxicol. 1980, 77, 245–250. [Google Scholar] [CrossRef]
- Shinohara, K.; Kuroki, S.; Miwa, M.; Kong, Z.-L.; Hosoda, H. Antimutagenicity of Dialyzates of Vegetables and Fruits. Agric. Biol. Chem. 1988, 52, 1369–1375. [Google Scholar]
- Edenharder, R.; Kurz, P.; John, K.; Burgard, S.; Seeger, K. In vitro effect of vegetable and fruit juices on the mutagenicity of 2-amino-3-methylimidazo[4,5-f]quinoline, 2-amino-3,4-dimethylimidazo[4,5-f]quinoline and 2-amino-3,8-dimethylimidazo[4,5-f]quinoxaline. Food Chem. Toxicol. 1994, 32, 443–459. [Google Scholar] [CrossRef]
- Edenharder, R.; Leopold, C.; Kries, M. Modifying actions of solvent extracts from fruit and vegetable residues on 2-amino-3-methylimidazo[4,5-f]quinoline (IQ) and 2-amino-3,4-dimethylimidazo[4,5-f]quinoxaline (MeIQx) induced mutagenesis in Salmonella typhimurium TA 98. Mutat. Res. 1995, 341, 303–318. [Google Scholar] [CrossRef]
- Ikken, Y.; Cambero, I.; Marin, M.; Haza, A.; Morales, P. Antimutagenic effect of fruit and vegetable aqueous extracts against n-nitrosamines evaluated by the ames test. J. Agric. Food Chem. 1998, 46, 5194–5200. [Google Scholar] [CrossRef]
- Ikken, Y.; Morales, P.; Martinez, A.; Marin, M.; Haza, A. Antimutagenic effect of fruit and vegetable ethanolic extracts against n-nitrosamines evaluated by the ames test. J. Agric. Food Chem. 1999, 47, 3257–3264. [Google Scholar] [CrossRef] [PubMed]
- Murugan, S.S.; Balakrishnamurthy, P.; Mathew, Y.J. Antimutagenic effect of broccoli flower head by the ames salmonella reverse mutation assay. Phytother. Res. 2007, 21, 545–547. [Google Scholar] [PubMed]
- Anupama, M.; Murgan, S.S.; Murthy, P.B. Broccoli flower head extract reduces mitomycin-C induced sister chromatid exchange in cultured human lymphocytes. Food Chem. Toxicol. 2008, 46, 3351–3353. [Google Scholar] [CrossRef] [PubMed]
- Edenharder, R.; Sager, J.W.; Glatt, H.; Muckel, E.; Platt, K.L. Protection by beverages, fruits, vegetables, herbs, and flavonoids against genotoxicity of 2-acetylaminofluorene and 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP) in metabolically competent V79 cells. Mutat. Res. 2002, 521, 57–72. [Google Scholar] [CrossRef]
- Keck, A.-S.; Finley, J.W. Aqueous extracts of selenium-fertilized broccoli increase selenoprotein activity and inhibit DNA single-strand breaks, but decrease the activity of quinone reductase in Hepa 1c1c7 cells. Food Chem. Toxicol. 2006, 44, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.; Yoon, S.; Carter, O.; Bailey, G.S.; Dashwood, R.H. Antioxidant and antigenotoxic activities of Angelica keiskei, Oenanthe javanica and Brassica oleracea in the Salmonella mutagenicity assay and in HCT116 human colon cancer cells. BioFactors 2006, 26, 231–244. [Google Scholar] [CrossRef] [PubMed]
- Yeh, J.-Y.; Ou, B.-R.; Liang, Y.-C.; Burchfiel, J.; Butler, J.A.; Forsberg, N.E.; Whanger, P.D. Mechanism for proliferation inhibition by various selenium compounds and selenium-enriched broccoli extract in rat glial cells. BioMetals 2006, 19, 611–621. [Google Scholar] [CrossRef]
- Platt, K.L.; Edenharder, R.; Aderhold, S.; Muckel, E.; Glatt, H. Fruits and vegetables protect against the genotoxicity of heterocyclic aromatic amines activated by human xenobiotic-metabolizing enzymes expressed in immortal mammalian cells. Mutat. Res. 2010, 703, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Sousa, C.; Fernandes, F.; Valentão, P.; Rodrigues, A.S.; Coelho, M.; Teixeira, J.P.; Silva, S.; Ferreres, F.; de Pinho, P.G.; Andrade, P.B. Brassica oleracea L. Var. costata DC and Pieris brassicae L. aqueous extracts reduce methyl methanesulfonate-induced DNA damage in V79 hamster lung fibroblasts. J. Agric. Food Chem. 2012, 60, 5380–5387. [Google Scholar] [CrossRef] [PubMed]
- Ferrarini, L.; Pellegrini, N.; Mazzeo, T.; Miglio, C.; Galati, S.; Milano, F.; Rossi, C.; Buschini, A. Anti-proliferative activity and chemoprotective effects towards DNA oxidative damage of fresh and cooked Brassicaceae. Br. J. Nutr. 2012, 107, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Bárta, I.; Smerák, P.; Polívková, Z.; Sestáková, H.; Langová, M.; Turek, B.; Bártová, J. Current trends and perspectives in nutrition and cancer prevention. Neoplasma 2006, 53, 19–25. [Google Scholar] [PubMed]
- Mandelová, L.; Totušek, J. Chemoprotective effects of broccoli juice treated with high pressure. Czech. J. Food Sci. 2006, 24, 19–25. [Google Scholar] [CrossRef]
- Tiku, A.B.; Abraham, S.K.; Kale, R.K. Protective effect of the cruciferous vegetable mustard leaf (Brassica campestris) against in vivo chromosomal damage and oxidative stress induced by γ-radiation and genotoxic chemicals. Environ. Mol. Mutagen. 2008, 49, 335–342. [Google Scholar] [CrossRef]
- Riso, P.; Martini, D.; Visioli, F.; Martinetti, A.; Porrini, M. Effect of broccoli intake on markers related to oxidative stress and cancer risk in healthy smokers and nonsmokers. Nutr. Cancer 2009, 61, 232–237. [Google Scholar] [CrossRef]
- McKay, D.L.; Blumberg, J.B. The role of tea in human health: An update. J. Am. Coll. Nutr. 2002, 21, 1–13. [Google Scholar] [CrossRef]
- McKay, D.L.; Blumberg, J.B. A Review of the bioactivity and potential health benefits of chamomile tea (Matricaria recutita L.). Phytother. Res. 2006, 20, 519–530. [Google Scholar] [CrossRef]
- Madrigal-Santillán, E.; Madrigal-Bujaidar, E.; Cruz Jaime, S.; del Carmen Valadez-Vega, M.; Teresa, M.; Guadalupe, K.; Antonio, J. The chemoprevention of chronic degenerative disease through dietary antioxidants: Progress, promise and evidences. In Oxidative Stress and Chronic Degenerative Diseases—A Role for Antioxidants; InTech: London, UK, 2013. [Google Scholar]
- Singh, O.; Khanam, Z.; Misra, N.; Srivastava, M.K. Chamomile (Matricaria chamomilla L.): An overview. Pharmacogn. Rev. 2011, 5, 82–95. [Google Scholar] [CrossRef]
- Hernández-Ceruelos, A.; Madrigal-Bujaidar, E.; de la Cruz, C. Inhibitory effect of chamomile essential oil on the sister chromatid exchanges induced by daunorubicin and methyl methanesulfonate in mouse bone marrow. Toxicol. Lett. 2002, 135, 103–110. [Google Scholar] [CrossRef]
- Jakovlev, V.; Isaac, O.; Flaskamp, E. Pharmacologic studies on chamomile compounds. VI. Studies on the antiphlogistic effect of chamazulene and matricine. Planta Med. 1983, 49, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Viola, H.; Wasowski, C.; Levi de Stein, M.; Wolfman, C.; Silveira, R.; Dajas, F.; Medina, J.; Paladini, A. Apigenin, a component of Matricaria recutita flowers, is a central benzodiazepine receptors-ligand with anxiolytic effects. Planta Med. 1995, 61, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Zeilmann, C.A.; Dole, E.J.; Skipper, B.J.; McCabe, M.; Low Dog, T.; Rhyne, R.L. Use of herbal medicine by elderly Hispanic and non-Hispanic white patients. Pharmacotherapy 2003, 23, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Howrey, B.T.; Peek, M.K.; McKee, J.M.; Raji, M.A.; Ottenbacher, K.J.; Markides, K.S. Chamomile consumption and mortality: A prospective study of mexican origin older adults. Gerontologist 2016, 56, 1146–1152. [Google Scholar] [CrossRef] [PubMed]
- Stavric, B.; Matula, T.I.; Klassen, R.; Downie, R.H. The effect of teas on the in vitro mutagenic potential of heterocyclic aromatic amines. Food Chem. Toxicol. 1996, 34, 515–523. [Google Scholar] [CrossRef]
- Hernández-Ceruelos, A.; Madrigal-Santillán, E.; Morales-González, J.A.; Chamorro-Cevallos, G.; Cassani-Galindo, M.; Madrigal-Bujaidar, E. Antigenotoxic effect of Chamomilla recutita (L.) rauschert essential oil in mouse spermatogonial cells, and determination of its antioxidant capacity in vitro. Int. J. Mol. Sci. 2010, 11, 3793–3802. [Google Scholar] [CrossRef]
- Drummond, E.M.; Harbourne, N.; Marete, E.; Martyn, D.; Jacquier, J.; O’Riordan, D.; Gibney, E.R. Inhibition of proinflammatory biomarkers in THP1 macrophages by polyphenols derived from chamomile, meadowsweet and willow bark. Phytother. Res. 2013, 27, 588–594. [Google Scholar] [CrossRef]
- Gomes-Carneiro, M.R.; Dias, D.M.M.; De-Oliveira, A.C.A.X.; Paumgartten, F.J.R. Evaluation of mutagenic and antimutagenic activities of α-bisabolol in the Salmonella/microsome assay. Mutat. Res. 2005, 585, 105–112. [Google Scholar] [CrossRef]
- Anter, J.; Romero-Jiménez, M.; Fernández-Bedmar, Z.; Villatoro-Pulido, M.; Analla, M.; Alonso-Moraga, Á.; Muñoz-Serrano, A. Antigenotoxicity, cytotoxicity, and apoptosis induction by apigenin, bisabolol, and protocatechuic acid. J. Med. Food 2011, 14, 276–283. [Google Scholar] [CrossRef]
- Safayhi, H.; Sabieraj, J.; Sailer, E.R.; Ammon, H.P. Chamazulene: An antioxidant-type inhibitor of leukotriene B4 formation. Planta Med. 1994, 60, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Capuzzo, A.; Occhipinti, A.; Maffei, M.E. Antioxidant and radical scavenging activities of chamazulene. Nat. Prod. Res. 2014, 28, 2321–2323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusconi, M.; Conti, A. Theobroma cacao L., the food of the gods: A scientific approach beyond myths and claims. Pharmacol. Res. 2010, 61, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Aprotosoaie, A.C.; Luca, S.V.; Miron, A. Flavor chemistry of cocoa and cocoa products: An overview. Compr. Rev. Food Sci. Food Saf. 2016, 15, 73–91. [Google Scholar] [CrossRef]
- McShea, A.; Ramiro-Puig, E.; Munro, S.B.; Casadesus, G.; Castell, M.; Smith, M.A. Clinical benefit and preservation of flavonols in dark chocolate manufacturing. Nutr. Rev. 2008, 66, 630–641. [Google Scholar] [CrossRef] [PubMed]
- Latif, R. Chocolate/cocoa and human health: A review. Neth. J. Med. 2013, 71, 63–68. [Google Scholar] [PubMed]
- Ellam, S.; Williamson, G. Cocoa and human health. Annu. Rev. Nutr. 2013, 33, 105–128. [Google Scholar] [CrossRef]
- Kim, J.; Kim, J.; Shim, J.; Lee, C.Y.; Lee, K.W.; Lee, H.J. Cocoa phytochemicals: Recent advances in molecular mechanisms on health. Crit. Rev. Food Sci. Nutr. 2014, 54, 1458–1472. [Google Scholar] [CrossRef]
- Latif, R. Health benefits of cocoa. Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 669–674. [Google Scholar] [CrossRef]
- Yamagishi, M.; Osakabe, N.; Natsume, M.; Adachi, T.; Takizawa, T.; Kumon, H.; Osawa, T. Anticlastogenic activity of cacao: Inhibitory effect of cacao liquor polyphenols against mitomycin C-induced DNA damage. Food Chem. Toxicol. 2001, 39, 1279–1283. [Google Scholar] [CrossRef]
- Cajurao, E.; Revale, I. Antigenotoxicity screening of coffee (Coffea arabica Linn) and Cacao (Theobroma cacao Linn.). Asian Pac. J. Educ. Art Sci. 2016, 3, 94–97. [Google Scholar]
- Ibero-Baraibar, I.; Azqueta, A.; Lopez de Cerain, A.; Martinez, J.A.; Zulet, M.A. Assessment of DNA damage using comet assay in middle-aged overweight/obese subjects after following a hypocaloric diet supplemented with cocoa extract. Mutagenesis 2015, 30, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Fujie, K.; Aoki, T.; Ito, Y.; Maeda, S. Sister-chromatid exchanges induced by trihalomethanes in rat erythroblastic cells and their suppression by crude catechin extracted from green tea. Mutat. Res. 1993, 300, 241–246. [Google Scholar] [CrossRef]
- Tanaka, R. Protective effects of (-)-epigallocatechin gallate and (+)-catechin on paraquat-induced genotoxicity in cultured cells. J. Toxicol. Sci. 2000, 25, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Law, Y.L.; Wong, T.P.W.; Yu, K.N. Influence of catechins on bystander responses in CHO cells induced by alpha-particle irradiation. Appl. Radiat. Isot. 2010, 68, 726–729. [Google Scholar] [CrossRef]
- Delgado, M.E.; Haza, A.I.; Arranz, N.; García, A.; Morales, P. Dietary polyphenols protect against N-nitrosamines and benzo(a)pyrene-induced DNA damage (strand breaks and oxidized purines/pyrimidines) in HepG2 human hepatoma cells. Eur. J. Nutr. 2008, 47, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Delgado, M.E.; Haza, A.I.; García, A.; Morales, P. Myricetin, quercetin, (+)-catechin and (-)-epicatechin protect against N-nitrosamines-induced DNA damage in human hepatoma cells. Toxicol. In Vitro 2009, 23, 1292–1297. [Google Scholar] [CrossRef] [PubMed]
- Haza, A.I.; Morales, P. Effects of (+)-catechin and (-)-epicatechin on heterocyclic amines-induced oxidative DNA damage. J. Appl. Toxicol. 2011, 31, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Papież, M.A. The influence of curcumin and (-)-epicatechin on the genotoxicity and myelosuppression induced by etoposide in bone marrow cells of male rats. Drug Chem. Toxicol. 2013, 36, 93–101. [Google Scholar] [CrossRef]
- Zor, M.; Aydin, S.; Güner, N.D.; Başaran, N.; Başaran, A.A. Antigenotoxic properties of Paliurus spina-christi Mill fruits and their active compounds. BMC Complement. Altern. Med. 2017, 17, 229. [Google Scholar] [CrossRef]
- De Lima, R.M.T.; Dos Reis, A.C.; de Menezes, A.A.; Santos, J.V.; Filho, J.W.; Ferreira , J.R.; de Alencar, M.V.; Khan, I.N.; Islam, A.; et al. Protective and therapeutic potential of ginger (Zingiber officinale) extract and [6]-gingerol in cancer: A comprehensive review. Phytother. Res. 2018, 32, 1885–1907. [Google Scholar] [CrossRef] [PubMed]
- Singletary, K. Ginger: An overview of health benefits. Nutr. Today 2010, 45, 171–183. [Google Scholar] [CrossRef]
- Srinivasan, K. Ginger rhizomes (Zingiber officinale): A spice with multiple health beneficial potentials. PharmaNutrition 2017, 5, 18–28. [Google Scholar] [CrossRef]
- Odunola, O.A. Comparative effects of some local food condiments on sodium arsenite-induced clastogenicity. Afr. J. Med. Med. Sci. 2003, 32, 75–80. [Google Scholar] [PubMed]
- Bidinotto, L.T.; Spinardi-Barbisan, A.L.T.; Rocha, N.S.; Salvadori, D.M.F.; Barbisan, L.F. Effects of ginger (Zingiber officinale Roscoe) on DNA damage and development of urothelial tumors in a mouse bladder carcinogenesis model. Environ. Mol. Mutagen. 2006, 47, 624–630. [Google Scholar] [CrossRef] [PubMed]
- Jayakumar, R.; Kanthimathi, M.S. Dietary spices protect against hydrogen peroxide-induced DNA damage and inhibit nicotine-induced cancer cell migration. Food Chem. 2012, 134, 1580–1584. [Google Scholar] [CrossRef] [PubMed]
- Jeena, K.; Liju, V.B.; Ramanath, V.; Kuttan, R. Protection against whole body γ-irradiation induced oxidative stress and clastogenic damage in mice by ginger essential oil. Asian Pac. J. Cancer Prev. 2016, 17, 1325–1332. [Google Scholar] [CrossRef]
- Yang, G.; Zhong, L.; Jiang, L.; Geng, C.; Cao, J.; Sun, X.; Liu, X.; Chen, M.; Ma, Y. 6-gingerol prevents patulin-induced genotoxicity in HepG2 cells. Phytother. Res. 2011, 25, 1480–1485. [Google Scholar] [CrossRef]
- Meschini, R.; Berni, A.; Filippi, S.; Pepe, G.; Grossi, M.R.; Natarajan, A.T.; Palitti, F. The micronucleus assay in mammalian cells in vitro to assess health benefits of various phytochemicals. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2015, 793, 79–85. [Google Scholar] [CrossRef]
- Yang, G.; Gao, X.; Jiang, L.; Sun, X.; Liu, X.; Chen, M.; Yao, X.; Sun, Q.; Wang, S. 6-Gingerol prevents MEHP-induced DNA damage in human umbilical vein endothelia cells. Hum. Exp. Toxicol. 2017, 36, 1177–1185. [Google Scholar] [CrossRef]
- Garg, S.N.; Siddiqui, M.S.; Agarwal, S.K. New fatty acid esters and hydroxy ketones from fruits of laurus nobilis. J. Nat. Prod. 1992, 55, 1315–1319. [Google Scholar] [CrossRef]
- Demir, V.; Gunhan, T.; Yagcioglu, A.K.; Degirmencioglu, A. Mathematical modelling and the determination of some quality parameters of air-dried bay leaves. Biosyst. Eng. 2004, 88, 325–335. [Google Scholar] [CrossRef]
- Patrakar, R.; Mansuriya, M.; Patil, P. Phytochemical and pharmacological review on Laurus Nobilis. Int. J. Pharm. Chem. Sci. 2012, 1, 595–602. [Google Scholar]
- Chahal, K.; Kaur, M.; Bhardwaj, U.; Singla, N.; Kaur, A.; Kaur, M.; Bhardwaj, U.; Singla, N.; Kaur, A. A review on chemistry and biological activities of Laurus nobilis L. essential oil. J. Pharmacogn. Phytochem. 2017, 6, 1153–1161. [Google Scholar]
- Abu-Dahab, R.; Kasabri, V.; Afifi, F.U. Evaluation of the volatile oil composition and antiproliferative activity of Laurus nobilis L. (Lauraceae) on breast cancer cell line models. Rec. Nat. Prod. 2014, 82, 136–147. [Google Scholar]
- Caputo, L.; Nazzaro, F.; Souza, L.; Aliberti, L.; De Martino, L.; Fratianni, F.; Coppola, R.; De Feo, V. Laurus nobilis: Composition of essential oil and its biological activities. Molecules 2017, 22, 930. [Google Scholar] [CrossRef] [PubMed]
- Türkez, H.; Geyikoglu, F. The effect of laurel leaf extract against toxicity induced by 2,3,7,8-tetrachlorodibenzo-p-dioxin in cultured rat hepatocytes. Arh. Hig. Rada Toksikol. 2011, 62, 309–315. [Google Scholar] [CrossRef]
- Türkez, H.; Toğar, B. Aluminium phosphide-induced genetic and oxidative damages in vitro: Attenuation by Laurus nobilis L. leaf extract. Indian J. Pharmacol. 2013, 45, 71–75. [Google Scholar] [CrossRef]
- Türkez, H.; Toğar, B. Aluminum phosphide-induced genetic and oxidative damages in rats: Attenuation by Laurus nobilis leaf extract. Toxicol. Ind. Health 2013, 29, 579–583. [Google Scholar] [CrossRef]
- Snuossi, M.; Trabelsi, N.; Ben Taleb, S.; Dehmeni, A.; Flamini, G.; De Feo, V. Laurus nobilis, zingiber officinale and anethum graveolens essential oils: Composition, antioxidant and antibacterial activities against bacteria isolated from fish and shellfish. Molecules 2016, 21, 1414. [Google Scholar] [CrossRef]
- Horvathova, E.; Turcaniova, V.; Slamenova, D. Comparative study of DNA-damaging and DNA-protective effects of selected components of essential plant oils in human leukemic cells K562. Neoplasma 2007, 54, 478–483. [Google Scholar] [PubMed]
- Nikolić, B.; Mitić-Ćulafić, D.; Vuković-Gačić, B.; Knežević-Vukčević, J. Modulation of genotoxicity and DNA repair by plant monoterpenes camphor, eucalyptol and thujone in Escherichia coli and mammalian cells. Food Chem. Toxicol. 2011, 49, 2035–2045. [Google Scholar] [CrossRef] [PubMed]
- Yogalakshmi, B.; Viswanathan, P.; Anuradha, C.V. Investigation of antioxidant, anti-inflammatory and DNA-protective properties of eugenol in thioacetamide-induced liver injury in rats. Toxicology 2010, 268, 204–212. [Google Scholar] [CrossRef] [PubMed]
- De Paula Porto, M.; Da Silva, G.N.; Luperini, B.C.; Bachiega, T.F.; de Castro Marcondes, J.P.; Sforcin, J.M.; Salvadori, D.M. Citral and eugenol modulate DNA damage and pro-inflammatory mediator genes in murine peritoneal macrophages. Mol. Biol. Rep. 2014, 41, 7043–7051. [Google Scholar] [CrossRef] [PubMed]
- Tiku, A.B.; Abraham, S.K.; Kale, R.K. Eugenol as an in vivo radioprotective agent. J. Radiat. Res. 2004, 45, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Abraham, S.K. Anti-genotoxicity of trans-anethole and eugenol in mice. Food Chem. Toxicol. 2001, 39, 493–498. [Google Scholar] [CrossRef]
- Mitić-Culafić, D.; Zegura, B.; Nikolić, B.; Vuković-Gacić, B.; Knezević-Vukcević, J.; Filipic, M. Protective effect of linalool, myrcene and eucalyptol against t-butyl hydroperoxide induced genotoxicity in bacteria and cultured human cells. Food Chem. Toxicol. 2009, 47, 260–266. [Google Scholar] [CrossRef]
- Gunaseelan, S.; Balupillai, A.; Govindasamy, K.; Ramasamy, K.; Muthusamy, G.; Shanmugam, M.; Thangaiyan, R.; Robert, B.M.; Prasad Nagarajan, R.; Ponniresan, V.K.; et al. Linalool prevents oxidative stress activated protein kinases in single UVB-exposed human skin cells. PLoS ONE 2017, 12, e0176699. [Google Scholar] [CrossRef]
- Doppalapudi, R.S.; Riccio, E.S.; Rausch, L.L.; Shimon, J.A.; Lee, P.S.; Mortelmans, K.E.; Kapetanovic, I.M.; Crowell, J.A.; Mirsalis, J.C. Evaluation of chemopreventive agents for genotoxic activity. Mutat. Res. 2007, 629, 148–160. [Google Scholar] [CrossRef]
- Khalil, M.Y.; Naguib, N.Y.; Sherbeny, S.E.E. A comparative study on the productivity and chemical constituents of various sources and species of calendula plants as affected by two foliar fertilizers. J. Appl. Sci. Res. 2005, 1, 176–189. [Google Scholar]
- Nelofer, J.; Khurshid, I.A.; Riffat, J. Calendula officinalis—An important medicinal plant with potential biological properties. Proc. Indian Natl. Sci. Acad. 2017, 83, 769–787. [Google Scholar]
- Arora, D.; Rani, A.; Sharma, A. A review on phytochemistry and ethnopharmacological aspects of genus Calendula. Pharmacogn. Rev. 2013, 7, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Murakami, T.; Kishi, A.; Kageura, T.; Matsuda, H. Medicinal flowers. III. Marigold. (1): Hypoglycemic, gastric emptying inhibitory, and gastroprotective principles and new oleanane-type triterpene oligoglycosides, calendasaponins A, B, C, and D, from Egyptian Calendula officinalis. Chem. Pharm. Bull. 2001, 49, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Ukiya, M.; Akihisa, T.; Yasukawa, K.; Tokuda, H.; Suzuki, T.; Kimura, Y. Anti-inflammatory, anti-tumor-promoting, and cytotoxic activities of constituents of marigold (Calendula officinalis) flowers. J. Nat. Prod. 2006, 69, 1692–1696. [Google Scholar] [CrossRef] [PubMed]
- Rehecho, S.; Uriarte-Pueyo, I.; Calvo, J.; Vivas, L.A.; Calvo, M.I. Ethnopharmacological survey of medicinal plants in Nor-Yauyos, a part of the Landscape Reserve Nor-Yauyos-Cochas, Peru. J. Ethnopharmacol. 2011, 133, 75–85. [Google Scholar] [CrossRef]
- Safdar, W.; Majeed, H.; Naveed, I.; Kayani, W.; Ahmed, H.; Hussain, S.; Kamal, A. Pharmacognostical study of the medicinal plant Calendula officinalis l. (family compositae). Int. J. Cell Mol. Biol. 2010, 1, 108–116. [Google Scholar]
- Singh, M.K.; Sahu, P.; Nagori, K.; Dewangan, D.; Kumar Alexander, T.A.; Badwaik, H.; Tripathi, D. Organoleptic properties in-vitro and in-vivo pharmacological activities of Calendula officinalis Linn: An over review. J. Chem. Pharm. Res. 2011, 3, 655–663. [Google Scholar]
- Dimer Leffa, D.; da Rosa, R.; Pazini Munhoz, B.; de Araújo Martins Mello, A.; Dagostim Mandelli, F.; de Aguiar Amaral, P.; Erna Rossatto, Â.; Andrade, V. Genotoxic and antigenotoxic properties of calendula o fficinalis extracts in mice treated with methyl methanesulfonate. Adv. Life Sci. 2012, 2, 21–28. [Google Scholar] [CrossRef]
- Abdel-Aziem, S.H.; Hassan, A.M.; El-Denshary, E.S.; Hamzawy, M.A.; Mannaa, F.A.; Abdel-Wahhab, M.A. Ameliorative effects of thyme and calendula extracts alone or in combination against aflatoxins-induced oxidative stress and genotoxicity in rat liver. Cytotechnology 2014, 66, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Muley, B.; Khadabadi, S.; Banarase, N. Phytochemical constituents and pharmacological activities of Calendula officinalis Linn (Asteraceae): A review. Trop. J. Pharm. Res. 2009, 8, 455–465. [Google Scholar] [CrossRef]
- Wang, M.; Tsao, R.; Zhang, S.; Dong, Z.; Yang, R.; Gong, J.; Pei, Y. Antioxidant activity, mutagenicity/anti-mutagenicity, and clastogenicity/anti-clastogenicity of lutein from marigold flowers. Food Chem. Toxicol. 2006, 44, 1522–1529. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.L.; Morris, B.; Tonnis, B.; Davis, J.; Pederson, G.A. Assessment of oil content and fatty acid composition variability in two economically important Hibiscus species. J. Agric. Food Chem. 2012, 60, 6620–6626. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.; Ikram, E.; Nazri, H. Roselle (Hibiscus sabdariffa L.) seeds nutritional composition protein quality and health benefits. Food 2008, 2, 1–16. [Google Scholar]
- Da-Costa-Rocha, I.; Bonnlaender, B.; Sievers, H.; Pischel, I.; Heinrich, M. Hibiscus sabdariffa L. A phytochemical and pharmacological review. Food Chem. 2014, 165, 424–443. [Google Scholar] [CrossRef] [PubMed]
- Mahadevan, N.; Shivali, K.P. Hibiscus sabdariffa Linn.An overview. Nat. Prod. Radiance 2009, 8, 77–83. [Google Scholar]
- Riaz, G.; Chopra, R. A review on phytochemistry and therapeutic uses of Hibiscus sabdariffa L. Biomed. Pharmacother. 2018, 102, 575–586. [Google Scholar] [CrossRef] [PubMed]
- Bako, I.G.; Mabrouk, M.A.; Abubakar, A. Antioxidant effect of ethanolic seed extract of Hibiscus sabdariffa linn (Malvaceae) alleviate the toxicity induced by chronic administration of sodium nitrate on some haematological parameters in wistars rats. Adv. J. Food Sci. Technol. 2009, 1, 39–42. [Google Scholar]
- Bolade, M.K.; Oluwalana, I.B.; Ojo, O. Commercial practice of roselle (Hibiscus sabdariffa L.) beverage production: Optimization of hot water extraction and sweetness level. World J. Agric. Sci. 2009, 5, 126–131. [Google Scholar]
- Okoro, C. Production of red wine from roselle (Hibiscus sabdariffa) and pawpaw (Carica papaya) using palm-wine yeast (Saccharomyces cerevisiae). Niger. Food J. 2010, 25, 158–164. [Google Scholar] [CrossRef]
- Ali, B.H.; Wabel, N.A.; Blunden, G. Phytochemical, pharmacological and toxicological aspects ofHibiscus sabdariffa L.: A review. Phytother. Res. 2005, 19, 369–375. [Google Scholar] [CrossRef]
- Nwaiwu, N.E.; Mshelia, F.; Raufu, I.A. Antimicrobial activities of crude extracts of Moringa Oleifera, Hibiscus Sabdariffa and Hibiscus esculentus seeds against some enterobacteria. J. Appl. Phytotechnol. Environ. Sanit. 2012, 1, 11–16. [Google Scholar]
- Adetutu, A.; Odunola, O.A.; Owoade, O.A.; Adeleke, O.A.; Amuda, O.S. Anticlastogenic effects of Hibiscus sabdariffa fruits against sodium arsenite-induced micronuclei formation in erythrocytes in mouse bone marrow. Phytother. Res. 2004, 18, 862–864. [Google Scholar] [CrossRef] [PubMed]
- Farombi, E.O.; Fakoya, A. Free radical scavenging and antigenotoxic activities of natural phenolic compounds in dried flowers of Hibiscus sabdariffa L. Mol. Nutr. Food Res. 2005, 49, 1120–1128. [Google Scholar] [CrossRef] [PubMed]
- Rosa, R.M.; Moura, D.J.; Melecchi, M.I.S.; dos Santos, R.S.; Richter, M.F.; Camarão, E.B.; Henriques, J.A.P.; de Paula Ramos, A.L.L.; Saffi, J. Protective effects of Hibiscus tiliaceus L. methanolic extract to V79 cells against cytotoxicity and genotoxicity induced by hydrogen peroxide and tert-butyl-hydroperoxide. Toxicol. In Vitro 2007, 21, 1442–1452. [Google Scholar] [CrossRef] [PubMed]
- Vilela, T.C.; Leffa, D.D.; Damiani, A.P.; Damazio, D.D.C.; Manenti, A.V.; Carvalho, T.J.G.; Ramlov, F.; Amaral, P.A.; Andrade, V.M.D. Hibiscus acetosella extract protects against alkylating agent-induced DNA damage in mice. An. Acad. Bras. Cienc. 2018, 90, 3165–3174. [Google Scholar] [CrossRef] [PubMed]
- Alonso, J. Tratado De Fitofármacos Y Nutracéuticos; Corpus: Rosario, Argentina, 2004; ISBN 987-20292-3-7. [Google Scholar]
- Barni, M.V.; Fontanals, A.; Moreno, S. Estudio de la eficacia antibiótica de un extracto etanólico de Rosmarinus officinalis L. contra Staphylococcus aureus en dos modelos de infección en piel de ratón. Bol. Latinoam. Caribe Plantas Med. Aromáticas 2009, 8, 219–223. [Google Scholar]
- Sardans, J.; Rodà, F.; Peñuelas, J. Effects of water and a nutrient pulse supply on Rosmarinus officinalis growth, nutrient content and flowering in the field. Environ. Exp. Bot. 2005, 53, 1–11. [Google Scholar] [CrossRef]
- Avila-Sosa, R.; Navarro-Cruz, A.R.; Vera-López, O.; Dávila-Márquez, R.M.; Melgoza-Palma, N.; Meza-Pluma, R. Romero (Rosmarinus officinalis L.): Una revisión de sus usos no culinarios. Cienc. Mar 2011, 15, 23–36. [Google Scholar]
- Begum, A.; Sandhya, S.; Shaffath Ali, S.; Vinod, K.R.; Reddy, S.; Banji, D. An in-depth review on the medicinal flora Rosmarinus officinalis (Lamiaceae). Acta Sci. Pol. Technol. Aliment. 2013, 12, 61–73. [Google Scholar]
- Ozcan, M.M.; Chalchat, J.-C. Chemical composition and antifungal activity of rosemary (Rosmarinus officinalis L.) oil from Turkey. Int. J. Food Sci. Nutr. 2008, 59, 691–698. [Google Scholar] [CrossRef]
- Miguel, M.G.; Guerrero, C.; Rodrigues, H.; Brito, J.; Duarte, F.; Venâncio, F.; Tavares, R. Essential oils of Rosmarinus officinalis L., effect of harvesting dates, growing media and fertilizers. Greece 2007, 24, 65–67. [Google Scholar]
- Lo, A.-H.; Liang, Y.-C.; Lin-Shiau, S.-Y.; Ho, C.-T.; Lin, J.-K. Carnosol, an antioxidant in rosemary, suppresses inducible nitric oxide synthase through down-regulating nuclear factor-kappaB in mouse macrophages. Carcinogenesis 2002, 23, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Sanders, C.; Diego, M.; Fernandez, M.; Field, T.; Hernandez-Reif, M.; Roca, A. EEG asymmetry responses to lavender and rosemary aromas in adults and infants. Int. J. Neurosci. 2002, 112, 1305–1320. [Google Scholar] [CrossRef] [PubMed]
- Del Baño, M.J.; Lorente, J.; Castillo, J.; Benavente-García, O.; del Río, J.A.; Ortuño, A.; Quirin, K.-W.; Gerard, D. Phenolic diterpenes, flavones, and rosmarinic acid distribution during the development of leaves, flowers, stems, and roots of Rosmarinus officinalis. Antioxidant activity. J. Agric. Food Chem. 2003, 51, 4247–4253. [Google Scholar] [CrossRef] [PubMed]
- Tschiggerl, C.; Bucar, F. Investigation of the volatile fraction of rosemary infusion extracts. Sci. Pharm. 2010, 78, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Slamenova, D.; Kuboskova, K.; Horvathova, E.; Robichova, S. Rosemary-stimulated reduction of DNA strand breaks and FPG-sensitive sites in mammalian cells treated with H2O2 or visible light-excited Methylene Blue. Cancer Lett. 2002, 177, 145–153. [Google Scholar] [CrossRef]
- Aherne, S.A.; Kerry, J.P.; O’Brien, N.M. Effects of plant extracts on antioxidant status and oxidant-induced stress in Caco-2 cells. Br. J. Nutr. 2007, 97, 321–328. [Google Scholar] [CrossRef]
- Zegura, B.; Dobnik, D.; Niderl, M.H.; Filipič, M. Antioxidant and antigenotoxic effects of rosemary (Rosmarinus officinalis L.) extracts in Salmonella typhimurium TA98 and HepG2 cells. Environ. Toxicol. Pharmacol. 2011, 32, 296–305. [Google Scholar] [CrossRef]
- Razavi-Azarkhiavi, K.; Behravan, J.; Mosaffa, F.; Sehatbakhsh, S.; Shirani, K.; Karimi, G. Protective effects of aqueous and ethanol extracts of rosemary on H2O2-induced oxidative DNA damage in human lymphocytes by comet assay. J. Complement. Integr. Med. 2014, 11, 27–33. [Google Scholar] [CrossRef]
- Pérez-Sánchez, A.; Barrajón-Catalán, E.; Caturla, N.; Castillo, J.; Benavente-García, O.; Alcaraz, M.; Micol, V. Protective effects of citrus and rosemary extracts on UV-induced damage in skin cell model and human volunteers. J. Photochem. Photobiol. B 2014, 136, 12–18. [Google Scholar] [CrossRef]
- Mezzoug, N.; Abrini, J.; Serano, A.M.; Alonso-Moraga, A.; Idaomar, M. Study on antigenotoxic effects of Moroccan medicinal plants and spices using the white/white + somatic assay in Drosophila. Afr. J. Tradit. Complement. Altern. Med. 2006, 3, 22–31. [Google Scholar]
- Manoharan, S.; Balakrishnan, S.; Vinothkumar, V.; Silvan, S. Anti-clastogenic potential of carnosic acid against 7,12-dimethylbenz(a)anthracene(DMBA)-induced clastogenesis. Pharmacol. Rep. 2010, 62, 1170–1177. [Google Scholar] [CrossRef]
- Furtado, R.A.; de Araújo, F.R.R.; Resende, F.A.; Cunha, W.R.; Tavares, D.C. Protective effect of rosmarinic acid on V79 cells evaluated by the micronucleus and comet assays. J. Appl. Toxicol. 2010, 30, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Ghisalberti, E.L. Propolis: A review. Bee World 1979, 60, 59–84. [Google Scholar] [CrossRef]
- De Groot, A.C. Propolis: A review of properties, applications, chemical composition, contact allergy, and other adverse effects. Dermat. Contact Atopic Occup. Drug 2013, 24, 263–282. [Google Scholar] [CrossRef] [PubMed]
- García-Viguera, C.; Greenaway, W.; Whatley, F.R. Composition of propolis from two different Spanish regions. Z. Für Naturforschung C 1992, 47, 634–637. [Google Scholar] [CrossRef]
- Brumfitt, W.; Hamilton-Miller, J.M.; Franklin, I. Antibiotic activity of natural products: 1. Propolis. Microbios 1990, 62, 19–22. [Google Scholar]
- Marcucci, M.C. Propolis: Chemical composition, biological properties and therapeutic activity. Apidologie 1995, 26, 83–99. [Google Scholar] [CrossRef]
- Castaldo, S.; Capasso, F. Propolis, an old remedy used in modern medicine. Fitoterapia 2002, 73, S1–S6. [Google Scholar] [CrossRef]
- Denisow, B.; Denisow-Pietrzyk, M. Biological and therapeutic properties of bee pollen: A review. J. Sci. Food Agric. 2016, 96, 4303–4309. [Google Scholar] [CrossRef]
- Banskota, A.H.; Tezuka, Y.; Kadota, S. Recent progress in pharmacological research of propolis. Phytother. Res. 2001, 15, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Farooqui, T.; Farooqui, A.A. Beneficial effects of propolis on human health and neurological diseases. Front. Biosci. 2012, 4, 779–793. [Google Scholar] [CrossRef]
- De Lima, R.O.A.; Bazo, A.P.; Said, R.A.; Sforcin, J.M.; Bankova, V.; Darros, B.R.; Salvadori, D.M.F. Modifying effect of propolis on dimethylhydrazine-induced DNA damage but not colonic aberrant crypt foci in rats. Environ. Mol. Mutagen. 2005, 45, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Aliyazicioglu, Y.; Demir, S.; Turan, I.; Cakiroglu, T.N.; Akalin, I.; Deger, O.; Bedir, A. Preventive and protective effects of Turkish propolis on H2O2-induced DNA damage in foreskin fibroblast cell lines. Acta Biol. Hung. 2011, 62, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Yalcin, C.O.; Aliyazicioglu, Y.; Demir, S.; Turan, I.; Bahat, Z.; Misir, S.; Deger, O. Evaluation of the radioprotective effect of Turkish propolis on foreskin fibroblast cells. J. Cancer Res. Ther. 2016, 12, 990–994. [Google Scholar] [PubMed]
- Benković, V.; Kopjar, N.; Horvat Knezevic, A.; Dikić, D.; Basić, I.; Ramić, S.; Viculin, T.; Knezević, F.; Orolić, N. Evaluation of radioprotective effects of propolis and quercetin on human white blood cells in vitro. Biol. Pharm. Bull. 2008, 31, 1778–1785. [Google Scholar] [CrossRef] [PubMed]
- Benković, V.; Orsolić, N.; Knezević, A.H.; Ramić, S.; Dikić, D.; Basić, I.; Kopjar, N. Evaluation of the radioprotective effects of propolis and flavonoids in gamma-irradiated mice: The alkaline comet assay study. Biol. Pharm. Bull. 2008, 31, 167–172. [Google Scholar] [CrossRef]
- Benkovic, V.; Knezevic, A.H.; Dikic, D.; Lisicic, D.; Orsolic, N.; Basic, I.; Kosalec, I.; Kopjar, N. Radioprotective effects of propolis and quercetin in gamma-irradiated mice evaluated by the alkaline comet assay. Phytomedicine 2008, 15, 851–858. [Google Scholar] [CrossRef]
- Benković, V.; Knezević, A.H.; Dikić, D.; Lisicić, D.; Orsolić, N.; Basić, I.; Kopjar, N. Radioprotective effects of quercetin and ethanolic extract of propolis in gamma-irradiated mice. Arh. Hig. Rada Toksikol. 2009, 60, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Benkovic, V.; Knezevic, A.H.; Orsolic, N.; Basic, I.; Ramic, S.; Viculin, T.; Knezevic, F.; Kopjar, N. Evaluation of radioprotective effects of propolis and its flavonoid constituents: In vitro study on human white blood cells. Phytother. Res. 2009, 23, 1159–1168. [Google Scholar] [CrossRef]
- Oršolić, N.; Car, N.; Lisičić, D.; Benković, V.; Knežević, A.H.; Dikić, D.; Petrik, J. Synergism between propolis and hyperthermal intraperitoneal chemotherapy with cisplatin on ehrlich ascites tumor in mice. J. Pharm. Sci. 2013, 102, 4395–4405. [Google Scholar] [CrossRef] [PubMed]
- Oršolić, N.; Skurić, J.; Dikić, D.; Stanić, G. Inhibitory effect of a propolis on di-n-propyl disulfide or n-hexyl salycilate-induced skin irritation, oxidative stress and inflammatory responses in mice. Fitoterapia 2014, 93, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Oršolić, N.; Benković, V.; Lisičić, D.; Dikić, D.; Erhardt, J.; Knežević, A.H. Protective effects of propolis and related polyphenolic/flavonoid compounds against toxicity induced by irinotecan. Med. Oncol. 2010, 27, 1346–1358. [Google Scholar] [CrossRef] [PubMed]
- Türkez, H.; Yousef, M.I.; Geyikoglu, F. Propolis prevents aluminium-induced genetic and hepatic damages in rat liver. Food Chem. Toxicol. 2010, 48, 2741–2746. [Google Scholar] [CrossRef] [PubMed]
- Türkez, H.; Yousef, M.I.; Geyikoglu, F. Propolis protects against 2,3,7,8-tetrachlorodibenzo-p-dioxin-induced toxicity in rat hepatocytes. Food Chem. Toxicol. 2012, 50, 2142–2148. [Google Scholar] [CrossRef] [PubMed]
- Francielli de Oliveira, P.; Lima, I.M.; Monteiro Neto, M.D.; Bastos, J.K.; da Silva Filho, A.A.; Tavares, D.C. Evaluation of genotoxicity and antigenotoxicity of artepillin C in V79 cells by the comet and micronucleus assays. Nutr. Cancer 2013, 65, 1098–1103. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, P.F.; Leandro, L.F.; Montanheiro, G.; Bastos, J.K.; da Silva Filho, A.A.; Tavares, D.C. Baccharin prevents genotoxic effects induced by methyl methanesulfonate and hydrogen peroxide in V79 cells. J. Food Sci. 2012, 77, T138–T142. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, P.F.; Monteiro Neto, M.A.B.; Leandro, L.F.; Bastos, J.K.; da Silva Filho, A.A.; Tavares, D.C. In vivo antigenotoxicity of baccharin, an important constituent of Baccharis dracunculifolia DC (Asteraceae). Basic Clin. Pharmacol. Toxicol. 2011, 109, 35–41. [Google Scholar] [CrossRef] [PubMed]
- De Azevedo Bentes Monteiro Neto, M.; de Souza Lima, I.M.; Furtado, R.A.; Bastos, J.K.; da Silva Filho, A.A.; Tavares, D.C. Antigenotoxicity of artepillin C in vivo evaluated by the micronucleus and comet assays. J. Appl. Toxicol. 2011, 31, 714–719. [Google Scholar] [CrossRef] [PubMed]
- Manzolli, E.; Serpeloni, J.; Grotto, D.; Bastos, J.; Antunes, L.; Barbosa, J.; Barcelos, G. Protective effects of the flavonoid chrysin against methylmercury-induced genotoxicity and alterations of antioxidant status, in vivo. Oxid. Med. Cell. Longev. 2015, 7. [Google Scholar] [CrossRef] [PubMed]
- Roberto, M.M.; Matsumoto, S.T.; Jamal, C.M.; Malaspina, O.; Marin-Morales, M.A. Evaluation of the genotoxicity/mutagenicity and antigenotoxicity/antimutagenicity induced by propolis and Baccharis dracunculifolia, by in vitro study with HTC cells. Toxicol. In Vitro 2016, 33, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Dornelas, C.A.; Cavalcanti, B.C.; Magalhães, H.I.F.; Jamacaru, F.V.F.; Furtado, F.N.N.; Juanes, C.D.; Melo, N.D.; Moraes, M.O. Potential chemoprotective effects of green propolis, L-lysine and celecoxib on bone marrow cells and peripheral blood lymphocytes of Wistar rats subjected to bladder chemical carcinogenesis. Acta Cir. Bras. 2014, 29, 423–428. [Google Scholar] [CrossRef] [Green Version]
- Santos, G.S.; Tsutsumi, S.; Vieira, D.P.; Bartolini, P.; Okazaki, K. Effect of Brazilian propolis (AF-08) on genotoxicity, cytotoxicity and clonogenic death of Chinese hamster ovary (CHO-K1) cells irradiated with (60)Co gamma-radiation. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2014, 762, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Shelef, O.; Weisberg, P.J.; Provenza, F.D. The Value of native plants and local production in an era of global agriculture. Front. Plant Sci. 2017, 8, 2068. [Google Scholar] [CrossRef] [PubMed]
- Summya, R. Cancer and Chemoprevention: An Overview; Springer: New York, NY, USA, 2017. [Google Scholar]
- Kang, S.H.; Kwon, J.Y.; Lee, J.K.; Seo, Y.R. Recent advances in in vivo genotoxicity testing: Prediction of carcinogenic potential using comet and micronucleus assay in animal models. J. Cancer Prev. 2013, 18, 277–288. [Google Scholar] [CrossRef]
- Sasaki, Y.F.; Sekihashi, K.; Izumiyama, F.; Nishidate, E.; Saga, A.; Ishida, K.; Tsuda, S. The comet assay with multiple mouse organs: Comparison of comet assay results and carcinogenicity with 208 chemicals selected from the IARC monographs and U.S. NTP Carcinogenicity Database. Crit. Rev. Toxicol. 2000, 30, 629–799. [Google Scholar] [CrossRef]
- Bhagat, J. Combinations of genotoxic tests for the evaluation of group 1 IARC carcinogens. J. Appl. Toxicol. 2018, 38, 81–99. [Google Scholar] [CrossRef]
- Kirkland, D.; Aardema, M.; Henderson, L.; Müller, L. Evaluation of the ability of a battery of three in vitro genotoxicity tests to discriminate rodent carcinogens and non-carcinogens I. Sensitivity, specificity and relative predictivity. Mutat. Res. 2005, 584, 1–256. [Google Scholar] [CrossRef]
- Amin, A.R.M.R.; Kucuk, O.; Khuri, F.R.; Shin, D.M. Perspectives for cancer prevention with natural compounds. J. Clin. Oncol. 2009, 27, 2712–2725. [Google Scholar] [CrossRef]
- Mohsenzadegan, M.; Seif, F.; Farajollahi, M.M.; Khoshmirsafa, M. Anti-oxidants as chemopreventive agents in prostate cancer: A gap between preclinical and clinical studies. Recent Patents Anticancer Drug Discov. 2018, 13, 224–239. [Google Scholar] [CrossRef]
- Paller, C.J.; Denmeade, S.R.; Carducci, M.A. Challenges of conducting clinical trials of natural products to combat cancer. Clin. Adv. Hematol. Oncol. 2016, 14, 447–455. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Types of Mechanisms | Examples of Dietary Antimutagens |
|---|---|
| EXTRACELLULAR | |
| 1. Inhibition of mutagen uptake | Dietary fibres, probiotics, grapefruit (naringenin). |
| 2. Inhibition of endogenous formation a) Inhibition of nitrosation b) Modification of the intestinal flora | Vitamins (ascorbic acid), sulphur compounds (cysteine, glutathione). Prebiotics, probiotics. |
| 3. Complexation and/or deactivation | Dietary fibres, chlorophyllin. |
| 4. Favouring absorption of protective agents | Vitamin D3. |
| INTRACELLULAR | |
| 5. Blocking or competition a) Scavenging of reactive oxygen species b) Protection of DNA nucleophilic sites | Mango (polyphenols), guava (gallocatechin) vitamins (β-carotene,α-tocopherol, ascorbic acid), pineapple, blueberries (anthocyanins). Ellagic acid, retinoids, polyamines. |
| 6. Stimulation of trapping and detoxification in non-target cells | N-Acetyl cysteine. |
| 7. Modification of transmembrane transport | Short chain fatty acids (caproate), dietary calcium. |
| 8. Modulation of xenobiotic metabolising enzymes a) Inhibition of promutagen activation b) Induction of detoxification pathways c) Inhibition of metabolic enzymes | Isothiocyanates, monocyclic monoterpenoids (limonene, methol), flavonoids, wheat bran. Polyphenols, indoles, diterpene esters. Grapefruit (naringin, naringenin). |
| 9. Modulation of DNA metabolism and repair | Cinnamaldehyde, vanillin. |
| 10. Regulation of signaling pathways | Pomegranate (polyphenols), β-glucans. |
| 11. Enhancement of apoptosis | Retinoids, flavonoids. |
| 12. Maintenance of genomic stability | Vitamins (folic acid, B12), minerals (selenium, zinc), polyphenols. |
| Prokaryote and Eukaryote Models Germinal Cell | ||
|---|---|---|
| In Vitro In Vivo | ||
| I. GENE MUTATIONS | ||
| Bacteria (Ames assay, SOS chromotest) Yeast/Fungus (S. cerevisiae assay, A. nidulans assay) | Mouse spot test Somatic mutations and recombination test (SMART) DNA microarrays Serial analysis of gene expression (SAGE) Specific genes targeting | Recessive lethal Specific locus test Abnormalities of the sperm |
| II. CHROMOSOME CHANGES | ||
| Fibroblast culture Lymphocyte culture Mouse lymphoma assay | Micronucleus assay (MN) | Dominant lethal Heritable translocations Cytogenetic sperm Aneuploidy |
| III. INDICATORS BIOLOGICAL DAMAGE | ||
| Gene recombination Unscheduled DNA synthesis (UDS) assay | Comet assay Sister chromatid exchange (SCE) MN | Comet assay MN, SCE UDS assay |
| Year | Main Objective and Conclusion | Type of Study | Assay Employed | Ref |
|---|---|---|---|---|
| Forms of broccoli evaluated [fresh, juice, dialysate, extracts (water, ethanol, acetone), deep-frozen commercial broccoli, steamed before being consumed, and boiled or cooked with microwaves] | ||||
| 1980 | Determination of the inhibitory potential of aqueous and acetone extractions from some common vegetables (including broccoli) against the mutagenicity induced by 3-methylcholanthrene and benzo[a]pyrene. The results showed that the extracts were antimutagenic in TA 100 strain and that the said capacity was proportional to the chlorophyllin content. | In vitro | AT | [52] |
| 1988 | Evaluation of the effect of aqueous dialysates of 16 vegetables and fruits on the mutagenicity of some carcinogens [3-amino-l-methyl-5H-pyrido (4,3-b) nature (Trp-P-2), benzo[a]pyrene, (BaP), aflatoxin Bl (AFB1), acrylamide, and N-methyl-N’-nitroso-N-nitrosoguanidine] in Salmonella typhimurium strain TA 100. All dialysates (including broccoli) inhibited the mutagenicity of Trp-P-2 and this antimutagenicity was maintained even after heating the dialysates at 100 °C for 20 min. The results also indicated that the apple dialysate was better at inhibiting the mutagenicity of BaP, AFB1, acrylamide, and N-methyl-N’-nitroso-N-nitrosoguanidine. | In vitro | AT | [53] |
| 1994 1995 | In summary, the researchers evaluated the antimutagenic potential of various vegetables [both in fresh form, as juices and extracts (n-hexane, dichloromethane, acetone, and 2-propanol)] against the toxic effect of 2-amino-3-methyl [4,5-f] -quinoline (IQ), 2-amino-3,4-dimethylimidazo [4,5-f] quinoline (MeIQ), and 2-amino-3,8-dimethylimidazo [4,5-f] quinoxaline (MeIQx) in strains, TA98 and TA100, of Salmonella typhimurium. They observed that both fresh vegetables and juices showed different levels, from weak to strong, of antimutagenic activity, unlike extracts where there was a greater antimutagenic effect, especially with n-hexane (96%), dichloromethane (64%), and acetone (44%). In both studies, they concluded that the peroxidase enzyme and the chlorophyll pigment present in broccoli and in other green vegetables, strongly contribute to this protective activity. | In vitro | AT | [54] [55] |
| 1998 1999 | Evaluation of the inhibitory effect of nine aqueous and ethanolic extracts from fruits and vegetables against the mutagenicity of N-nitrosodimethylamine (NDMA), N-nitrosopyrrolidine (NPYR), N-nitrosodibutylamine (NDBA), and N-nitrosopiperidine (NPIP). Both types of extracts showed an antimutagenic effect in the range of 50 microg/plate–2000 microg/plate, observing that broccoli was classified between the second and third places of effectiveness against all mutants. | In vitro | AT | [56] [57] |
| 2007 | The antimutagenic effect of broccoli flower head against mitomycin C (MMC) by the Ames Salmonella reverse mutation assay. Three strains (TA 98, TA102, and TA 1535) were challenged with the broccoli flower head ethanol extract (BFHEE) at concentrations of 23 mg/plate and 46 mg/plate. The conclusion of the experiment was that the BFHEE was not cytotoxic; on the contrary, the highest concentration showed a significant antimutagenic potential against MMC. | In vitro | AT | [58] |
| 2008 | This study is a continuation of the experiment conducted by Murugan SS et al. (2007). The investigation was on the effect of the BFHEE on the sister chromatid exchange (SCE) induced by MMC on cultured human peripheral blood lymphocytes. The enumeration of SCE in second division mitotic cells indicated that the BFHEE significantly reduced the SCE induced by MMC in both concentrations tested (200 microg/mL and 400 microg/mL). | In vitro | SCE | [59] |
| 2002 | Protective effect of broccoli (raw and cooked) against genotoxicity of 2-acetylaminofluorene (AAF) and 2-amino-1-methyl-6-phenylimidazo [4, 5-b] pyridine (PhIP) in V79 cells. The genotoxic activity of AAF and PhIP was strongly reduced in a dose-related manner by broccoli, demonstrating that protection of vegetables against genotoxicity of heterocyclic aromatic amines may take place within metabolically competent mammalian cells. | In vitro | CA | [60] |
| 2006 | The objective of the study was to determine whether selenized broccoli extracts decrease the hydrogen peroxide-induced DNA single-strand breaks measured in mouse hepatoma cells (Hepa 1c1c7 cells). The results showed that previous fertilization of broccoli with selenium was significantly effective (94%) to reduce the single-strand breaks to the genetic material. The data suggest that selenium is an important component for decreasing oxidative stress, but maximizing its content in the cultivation of broccoli might also stimulate its capacity to induce phase II detoxification proteins. | In vitro | CA | [61] |
| 2006 | This study examined the antigenotoxic and antioxidant properties of chlorophyll-rich methanol extracts of Brassica oleracea. The extract was highly effective in assays that measured ferric reducing/antioxidant power, oxygen radical absorbance property, and Fe2+/H2O2-DNA mediated nicking. Subsequently, by means of the “comet’ assay”, the methanol extract protected against H2O2-induced genotoxic damage in human HCT116 colon cancer cells. These findings provide support for the antigenotoxic and antioxidant properties of chlorophyll-rich extracts of B. oleracea through mechanisms that include inhibition of carcinogen activation and scavenging of reactive oxygen species. | In vitro | CA | [62] |
| 2006 | The objective was to investigate the differential effects of various selenium (Se) compounds and Se-enriched broccoli extracts on cell proliferation and the possible mechanism responsible for the Se-induced growth inhibition. C6 rat glial cells were incubated with graded concentrations up to 1000 nM of selenite, selenate, selenomethionine (SeM), Se-methyl-selenocysteine (SeMCys), high-Se broccoli (H-SeB) extract, or low-Se broccoli (L-SeB) extract for 24 h and 48 h. The comet assay indicated that there was no significant DNA single-strand break found for all Se treatments in C6 cells. In addition, the Se-induced proliferation inhibition may involve an H2O2-dependent mechanism with elevated cellular glutathione peroxidase (cGPX) activity. Both H-SeB and L-SeB inhibited C6 cell proliferation, but H-SeB was less inhibitory than L-SeB. | In vitro | CA | [63] |
| 2010 | This study was a continuation of the experiment conducted by Edenharder R. et al. (2002). Chinese hamster lung fibroblasts, genetically engineered for the expression of rat cytochrome P450 dependent monooxygenase 1A2 and rat sulfotransferase 1C1 (V79-rCYP1A2-rSULT1C1 cells), were utilized to detect protective effects of plant-based beverages against the genotoxicity induced by 2-acetylaminofluorene (AAF) or 2-amino-1-methyl-6-phenylimidazo [4,5-b]pyridine (PhIP). The genotoxic activity of PhIP was strongly reduced in a dose-related manner by broccoli (raw and cooked). Similarly, the genotoxicity of AAF showed a reduction, although in a lower potency by cooked broccoli. These results are suggestive for enzyme inhibition (possibly involves CYP1A2) as a protection mechanism induced by the complex chemical mixtures present in the plant-based beverages. | In vitro | CA | [64] |
| 2012 | B. oleracea L. var. costata leaves and Pieris brassicae L. larvae aqueous extracts were assayed for their potential characteristics to prevent/induce DNA damage. Using the comet assay, none of the extracts revealed to be genotoxic by itself, and both showed protection, more advisable on larvae extracts, against MMS-induced genotoxicity. As for the genotoxic/antigenotoxic effects of Brassica vegetables, they are commonly attributed to isothiocyanates. The extracts were screened in search of these compounds by means of a headspace-solid-phase microextraction/gas chromatography-mass spectrometry. These findings demonstrate that both extracts could be useful against damage caused by genotoxic compounds; with the larvae extract being the most promising. | In vitro | CA | [65] |
| 2012 | Because cruciferous species are usually processed before eating and the real impact of cooking practices on their bioactive properties is not fully understood, Ferrarini L. et al., evaluated the effect of common cooking practices (boiling, microwaving, and steaming) on the biological activities of broccoli, cauliflower, and Brussels sprouts. The objective was to determine and compare the chemoprotective capacity of fresh and cooked vegetable extracts against oxidative DNA damage induced by 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium in HT-29 human colon carcinoma cells. The results indicated that both types of vegetables showed a protective activity comparable to vitamin C. In conclusion, the cooking methods applied did not alter the antioxidant effect of raw vegetables. | In vitro | CA | [66] |
| 2006 | The purpose of this study was to examine the antimutagenic and antigenotoxic effect of vegetable homogenates (broccoli, cauliflower, red cabbage, onion, garlic) on the damage produced by AFB1, 2-amino-3-methylimidazo [4,5, -f] quinolin (IQ), and N-nitroso-N-methylurea (MNU). The Ames test, in vivo micronuclei, and the comet assay showed that all homogenates contained clear antigenotoxic activities. Only, in the Ames test, the effect of some phytochemicals against the direct mutagen, MNU, was lower in comparison to the indirect mutagens, AFB1 and IQ. | In vivo | CA & MN | [67] |
| 2006 | The analysis was on the chemoprotective effect of broccoli juice treated under high pressure (500 MPa during 10 min) against the damage produced by N-nitroso-N-methylurea (MNU) in BALB/C mice. After administering broccoli juice (0.2 mL/10 g) for 14 days and a unique application of the mutagen, a statistically significant decrease in the number of micronuclei induced by the MNU was obtained. As a conclusion, substances that favor chemoprotective capacity, such as vitamins, polyphenolic componds, glucosinates, etc., can be preserved in broccoli juice treated at high pressure. | In vivo | MN | [68] |
| 2008 | These researchers evaluated the protective effect of mustard leaf (Brassica campestris), a popular Indian cruciferous vegetable, against chromosomal damage and oxidative stress induced by gamma-radiation, cyclophosphamide (CP), and urethane (URE) through the in vivo bone marrow micronucleus test. A pretreatment with 50 mg/kg-250 mg/kg of mustard leaf extract (MLE) for seven days significantly reduced the frequencies of micronuclei induced by gamma-radiation, CPH, and URE. The protective effect against chromosomal damage was associated with the modulation of lipid peroxidation and with an increase in GSH and the GSH-dependent enzyme, glutathione S-transferase (GST). | In vivo | MN | [69] |
| 2009 | The objective was to evaluate the protective effect of broccoli intake in smokers and non-smokers. 20 healthy young people (10 smokers and 10 nonsmokers) were randomized in a crossover design and received a 200 g portion of brocolli or maintained a controlled diet for 10 days each. Blood samples were collected at 0, 10, 30, and 40 days to evaluate the DNA damage. The ex vivo protection of DNA damage induced by H2O2 and damage of endogenous DNA were evaluated in lymphocytes by the comet assay. Strand breaks decreased significantly after the broccoli diet, both in smokers and nonsmokers. | Clinical study | CA | [70] |
| Year | Main Objective and Conclusion | Type of Study | Assay Employed | Ref |
|---|---|---|---|---|
| Main phytochemicals [sulforaphane (SUL), phenethyl isothiocyanate (PEITC), allyl isothiocyanate (AITC), and indole-3-carbinol (I3C)] | ||||
| 1996 | The study tested the sulforaphane (SUL) capacity to inhibit the genotoxicity of N-nitrosodimethylamine (NDMA) in the TA100 strain of Salmonella typhimurium. The results showed that SUL reduces the mutagenicity of NDMA at concentrations of 0.8 microM and suggest that its mechanism of action involves the inhibition of cytochrome P450 isoenzyme 2E1 (CYP2E1). | In vitro | AT | [41] |
| 2003 | The aim was to determine the antimutagenic potential of sulforaphane (SUL) against different heterocyclic amines present in cooked foods. The use of strains, TA98 and TA100, of Salmonella typhimurium demonstrated that SUL is a potent inhibitor. Approximately 60% of the mutagenicity was induced by 2-amino-3-methylimidazo [4,5-f] quinoline (IQ), 2-amino -3,4-dimethylimidazo [4,5-f] quinoline (MeIQ), and 2-amino-3,8-dimethylimidazo [4,5-f] quinoxaline (MeIQx). | In vitro | AT | [42] |
| 2009 | This study was a continuation of the experiment conducted by Shishu and Kaur (2003). In this case, the antimutagenic potential of SUL (extracted from broccoli) and sulforaphane (found in the radish) was compared and evaluated against the same heterocyclic amines present in cooked foods. The use of the same strains of S. typhimurium demonstrated that both isothiocyanates were potent inhibitors of the mutagenicity induced by all the mutagens tested. However, sulforaphane showed a greater chemoprotective activity than SUL did. | In vitro | AT | [43] |
| 1998 | The study demonstrated that in human liver cells, T5-2E1 and T5-1A2, the N-nitrosodimethylamine (NDMA) and 2-amino-3-methylimidazo [4,5-f] quinoline (IQ) generated DNA strand breaks. The comet assay showed that SUL markedly reduced the DNA damage induced by these mutagens; likewise, the results suggest that the inhibition of the said genotoxicity was mediated by the action of CYP2E1 and CYP1A2 isoenzymes, which contributes to a relevant chemopreventive activity. | In vitro | CA | [44] |
| 2001 | The study showed that a pretreatment with SUL in LS-174 cells before exposing them to BaP for 24 h reduced the number of single-strand DNA breaks (approximately 30%) generated by the carcinogen. This suggests that indoles and isothiocyanates (ITCs) from some cruciferous vegetables (including broccoli) can stimulate apoptosis in human colon adenocarcinoma and induce DNA protection. | In vitro | CA | [45] |
| 2004 | The aim was to evaluate the ability of indole-3-carbinol (I3C) to reduce DNA damage generated by N-methyl-N’-nitro-N-nitrosoguanidine (MNNG) in cultured Chinese hamster lung fibroblast cells (CH V-79). The result was that MNNG produced DNA single strand breaks in a dose- and time-dependent manner, as was determined by the increase in the tail moment of the comet. In contrast, when the cells were pretreated with I3C, a significant protection was induced in the DNA. Thus, concluding that I3C can be considered a relevant chemopreventive agent capable of inducing the DNA repair. | In vitro | CA | [46] |
| 2006 | The evaluation was on the protective effect of the three most common isothiocyanates (ITCs) of broccoli (phenethyl isothiocyanate (PEITC), allyl isothiocyanate (AITC), and indole-3-carbinol (I3C)) towards DNA damage induced by N-nitrosamines in HepG2 cells. The results showed that none of the concentrations used of the ITCs caused DNA damage per se. The three ITCs showed a genoprotective effect of oxidative damage induced by N-nitrosopyrrolidine (NPYR) and/or N-nitrosodimethylamine (NDMA); however, the most important protection corresponded to I3C and PEITC against NPYR. | In vitro | CA | [47] |
| 2008 | The study was on the protective effect of isothiocyanates (ITCs) alone or in combination with vitamin C towards oxidative DNA damage induced by N-nitrosodibutylamine (NDBA) and/or N-nitrosopiperidine (NPIP) in HepG2 cells. The conclusions were that both the PEITC and the I3C showed a weak protective effect (27%) against the NDBA and NPIP. In contrast, HepG2 cells treated with the combination of vitamin C and each of the ITCs presented a stronger genoprotection of oxidative damage induced by these carcinogens. This evidence suggests that a possible mechanism of action of ITCs (alone or in combination with vitamins) could be regulating the bioactivation of NDBA and NPIP through cytochrome P450. | In vitro | CA | [48] |
| 2009 | Due to the evidence that exposure to certain pesticides represents a potential risk to human health, the antigenotoxic capacity of SUL against endosulfan, chlorpyrifos, and thiram in human lymphocytes was assessed by the comet assay. The mixture of pesticides at an environmentally relevant concentration (5 microM each) produced DNA damage in the lymphocytes while with a pre-incubation and co-incubation of SUL, a significant genoprotection was observed in a concentration dependent manner(between 10 microg/mL–20 microg/mL). These data suggest that the exposure to low levels of these pesticide mixtures can induce DNA damage, and the presence of SUL in the diet can reduce the incidence of this genetic damage, especially in farm workers. | In vitro | CA | [49] |
| 2013 | Protecting the eye lens against oxidative stress is of great importance to delay the onset of cataracts. Sulforaphane may be a good strategy to provide cytoprotection against oxidative stress. Therefore, the ability of SUL to perform this function on the lens cells was evaluated and its potential to delay the onset of the cataract was established. The comet assay determined the level of DNA strand breaks in human lens epithelial cell line, FHL124. The exposure of 30 μM of H2O2 to these cells caused a reduction in cell viability and an increase in cytotoxicity; whereas a pre-treatment with SFN inhibited these effects and significantly reduced the DNA strand breaks induced by H2O2. | In vitro | CA | [50] |
| 1997 | Using the micronucleus assay, the mutagenicity of the pesticide, propoxur, and its inhibition was determined by the administration of indole-3-carbinol (I3C) in Swiss mice. Intraperitoneal administration of propoxur (25 mg/kg) induced MN formation in bone marrow cells after a 48h exposure. In contrast, the application of I3C (500 mg/kg body weight) significantly inhibited the genotoxicity. | In vivo | MN | [51] |
| Component | RT2 | Area in the Plant (%) |
|---|---|---|
| (E)-β- farnesene | 38.46 | 28.17 |
| Germacrene-D | 39.23 | 2.19 |
| Unidentified sesquiterpene | 40.07 | 1.40 |
| Unidentified sesquiterpene | 41.17 | 0.78 |
| (Z,E)-α- farnesene | 41.35 | 1.59 |
| Unidentified sesquiterpene | 48.52 | 0.71 |
| α-bisabolol oxide A | 54.46 | 41.77 |
| α-bisabolol oxide B | 49.28 | 4.31 |
| α-bisabolol oxide | 50.65 | 5.30 |
| α-bisabolol | 51.18 | 2.31 |
| Chamazulene | 52.80 | 2.39 |
| 1,6-dioxaspiro [4,4] non-3-ene,2-(2,4 hexadyn-1-ylidene) | 60.73 | 2.19 |
| Hexatriacontane | 67.49 | 0.50 |
| Year | Main objective and Conclusion | Type of Study | Type of Procyanidin | Ref |
|---|---|---|---|---|
| 2008 2009 | The aim was to determine the protective effect of CAT and EPI against N-nitrosamines and benzo[a]pyrene-induced DNA damage (strand breaks and oxidized purines/pyrimidines) in HepG2 human hepatoma cells. Both polyphenols in concentrations of 10 microM decreased the DNA strand breaks (approximately 30%) generated by the action of mutagens. | In vitro | CAT & EPI | [100] [101] |
| 2011 | This study analized the effect of procyanidins on the oxidative DNA damage induced by some heterocyclic amines in human hepatoma cells (HepG2). The three amines (8-MeIQx, 4,8-diMeIQx, PhIP) increased the purines and pyrimidines oxidized; and, consequently, the number of breaks of DNA strands. In contrast, the action of procyanidins reduced the effect. | In vitro | CAT & EPI | [102] |
| 2013 | This in vivo study evaluated the influence of EPI on the genotoxicity induced by etoposide in bone marrow cells of male rats. The results showed that EPI significantly reduced the DNA strand break produced by this drug. | In vivo | EPI | [103] |
| 2017 | The objective was to isolate and evaluate the antigenotoxic capacity of the main phytochemicals of Paliurus spina-christi Mill in a Chinese hamster lung fibroblasts (V79) cell line. The result was that the methanol extract of this typical Turkish fruit did not induce DNA damage. In contrast, there was a significant reduction of isolated procyanidins on DNA damage induced by H2O2. | In vitro | CAT & GALL | [104] |
| Year | Main objective and Conclusion | Assay Employed | Type of Study | Ref |
|---|---|---|---|---|
| Eucalyptol (EUC) | ||||
| 2007 | This study examined the possible protective effect of EUC against the DNA damage induced by H2O2 in human leukemic K562 cells. The results were not fully conclusive. No significant decrease in the level of breaks of single strands of DNA was observed. | CA | In vitro | [125] |
| 2011 | The aim was to examine the protective potential of plant monoterpenes against 4-Nitroquinoline 1-oxide (4NQO)-induced genotoxicity in the Vero cell line. Incubation of 4NQO-pretreated Vero cells with EUC resulted in a significant reduction of the tail moment. However, higher concentrations of monoterpenes induced DNA strand breaks. As in the previous study, the results are not fully conclusive and suggest that EUC can stimulate error-free DNA repair processes and may act as bioantimutagen. | CA | In vitro | [126] |
| Eugenol (EUG) | ||||
| 2010 | This study investigated the preventive effect of EUG on thioacetamide (TA) -induced liver injury. The pretreatment of EUG decreased the elevated expression of the COX-2 gene and DNA strand breaks induced by TA. These findings suggest that EUG reduces the genotoxic effects of TA in the liver. | CA | In vivo | [127] |
| 2014 | The evaluation was on the modulating effect of EUG (0.31 μg/mL, 0.62 μg/mL, 1.24 μg/mL, and 2.48 μg/mL) on DNA damage induced by doxorubicin (DXR) in mouse peritoneal macrophages. The data were confusing because EUG showed both genotoxic and antigenotoxic potential. These results suggest that EUG can modulate the DNA damage induced by DXR, but it should be cautiously used and investigations should be extended to confirm whether they could induce a primary DNA damage. | CA | In vitro | [128] |
| 2004 | Three doses of EUG (75 mg/kg,150 mg/kg, and 300 mg/kg) were administered to Swiss albino mice before being exposed to 1.5 Gy of gamma radiation. The results showed a significant reduction in the frequency of micronucleated polychromatic erythrocytes (MNPE) with the three doses used; in addition, this decrease was sustained up to 72 h after the irradiation. These data revealed that the EUG exerted a significant protection against oxidative stress and suggest a radioprotective potential. | MN | In vivo | [129] |
| 2001 | The aim was to determine the antigenotoxic potential of EUG against the damage produced by cyclophosphamide (CP), procarbazine (PCB), N-methyl-N’-nitro-N-nitrosoguanidine (MNNG), and urethane (URE). The oral administration of EUG (50 mg/kg–500 mg/kg) before injecting the genotoxins showed a dose-dependent protective effect. It also confirmed that the EUG is not a micronucleus-inducing agent in mouse bone marrow. | MN | In vivo | [130] |
| Linalool (LIN) | ||||
| 2009 | The results of this study suggest that LIN can be considered an important antioxidant agent, because it is a natural monoterpene capable of reducing DNA damage induced by t-butyl hydroperoxide in two types of cell lines (human hepatoma (HepG2) and human B lymphoid cells (NC-NC)) by approximately 50%. | CA | In vitro | [131] |
| 2017 | This study analyzed the preventive effect of LIN against the oxidative imbalance induced by ultraviolet-B radiation (UVB) in human skin cells (HDFa). LIN significantly reduced the formation of 8-deoxy guanosine mediated by UVB. This result suggests that this natural monoterpene can be considered a photoprotective agent for preventing the formation of reactive oxygen species (ROS). | CA | In vitro | [132] |
| Geraniol (GER) | ||||
| 2007 | There is no concrete evidence on the antigenotoxic capacity of GER. The results of this study suggest that it is not a clastogenic agent; because it does not increase the frequency of MN in mouse bone marrow. | MN | In vitro | [133] |
| Chemical Groups | Compounds |
|---|---|
| Alcohols | Benzyl alcohol, cinnamyl alcohol, glycerol, α-glycerol phosphate, hydroquinone, isobutenol, phenethyl alcohol |
| Aldehydes | Benzaldehyde, caproic aldehyde, p-hydroxybenzaldehyde, isovanillin, vanillin |
| Aliphatic acids and aliphatic esters | Acetic acid, angelic acid, butyric acid, crotonic acid, fumaric acid, isobutyric acid, methylbutyric acid, isobutyl acetate, isopentyl acetate |
| Amino acids | Alanine, β-alanine, α-aminobutyric acid, γ-aminobutyric acid, arginine, asparagine, aspartic acid, cysteine, glutamic acid, glycine, histidine, isoleucine, leucine, lysine, methionine, ornithine, phenylalanine, proline, serine, threonine, tryptophan, tyrosine, valine |
| Aromatic acids | Benzoic acid, caffeic acid, cinnamic, coumaric, acid, ferulic acid, gallic acid, gentisic acid, hydroxycinnamic acid, p-hydroxybenzoic acid, isoferulic acid, 4-methoxy cinnamic acid, salicylic acid, vanillic acid |
| Aromatic esters | Benzyl acetate, benzyl benzoate, benzyl caffeate, benzyl coumarate, benzyl ferulate, benzyl isoferulate, benzyl salicylate, butenyl caffeate, butyl caffeate, cinnamyl benzoate, cinnamyl caffeate, ethyl benzoate |
| Chalcones | Alpinetin chalcone, naringenin chalcone, pinobanksin chalcone, pinocembrin chalcone, sakuranetin chalcone |
| Flavanones | Naringenin, pinobanksin, pinobanksin-3-acetate, pinobanksin-3-butyrate, pinobanksin-3-hexanoate, pinobanksin-3-methyl ether, pinobanksin-3-pentanoate |
| Flavones and flavonols | Acacetin, apigenin, apigenin-7-methyl ether, galagin, galagin-3-methyl ether, izalpinin, isorhamnetin, kaempferol, quercetin, ramnetin, ramnocitrin |
| Waxy acids | Arachid acid, behenic acid, cerotic acid, lauric acid, linoleic acid, lignoceric acid, montanic acid, myristic acid, oleic acid, palmitic acid, stearic acid |
| Ketones | Acetophenone, dihydroxy-acetophenone, methylacetophenone |
| Terpenoids and other compounds | α-acetoxybetulenol, β-bisabolol, 1,8-cineole, α-copaene, cymene, limonene, styrene, xanthorreol, naphthalene, sesquiterpene alcohol, sesquiterpene diol |
| Steroids | Calinasterol acetate, b-dihydrofucosterol acetate, ucosterol acetate, stigmasterol acetate |
| Sugars | Fructofuranose, α-D-glucopyranose, β-D-glucopyranose |
| Year | Authors | Main Objective, Results, and Conclusion | Ref |
|---|---|---|---|
| Comet assay in vivo | |||
| 2005 | de Lima et al. | Evaluation of the modifying effect of an aqueous extract of propolis (AEP) on the formation of aberrant crypt foci (ACF) and DNA damage induced by 1,2-dimethylhydrazine (DMH) in the colon of Wistar rats. The AEP was orally administered at approximate doses of 12 mg/kg, 34 mg/kg, 108 mg/kg, and 336 mg/kg of body weight/day. Subsequently, the animals received different injections of DMH. The results showed that the simultaneous administration of both compounds reduced the induction of DNA damage in the colon. However, AEP had no effect on the formation of ACF. | [188] |
| 2008 | Benkovic et al. | The protective effect of a water-soluble derivate of propolis (WSDP) and some flavonoids (including caffeic acid, chrysin, and naringin) against damage caused by two doses of gamma irradiation was analyzed in this study. Like the previous study, the data indicated that all compounds administered before irradiation protect animals from lethal effects of whole-body irradiation and diminish primary DNA damage in their white blood cells. The WSDP showed the best radioprotective effect. | [192] |
| 2008 | Benkovic et al. | The objective was to analyze the radioprotective effects of the ethanolic extract of propolis (EEP) and quercetin on the white blood cells of mice irradiated with gamma rays. Irradiation was performed using a gamma-ray source [(60) Co], and the absorbed dose was 9 Gy. The efficacy of the compounds was evaluated intraperitoneally at a dose of 100 mg/kg for three consecutive days before and/or after irradiation. The results showed that propolis and quercetin protected these cells from the lethal effect of irradiation and decreased primary DNA damage. | [193] |
| 2009 | Benkovic et al. | The aim of this study was to assess radioprotective effects of quercetin and the ethanolic extract of propolis (EEP) in CBA mice exposed to a single radiation dose (4 Gy) of gamma radiation. Similar to the two previous studies, the mice were treated daily with 100 mg/kg of quercetin or EEP for three consecutive days before or after gamma irradiation. The leukocyte count was determined in blood drawn from the tail vein, and DNA in these cells was assessed using the comet assay. Animals pretreated with the compounds were less sensitive to irradiation. Those that received the therapy after irradiation showed a slight, but not significant, increase in the total leukocyte count compared to the negative control. EEP and quercetin were confirmed not to be genotoxic for non-irradiated mice. | [194] |
| Comet assay in vitro | |||
| 2011 | Aliyazicioglu et al. | The analysis was on the antigenotoxic potential of propolis extracts in foreskin fibroblast cells against oxidative damage induced by hydrogen peroxide. The results showed a significant decrease in DNA damage induced by H2O2 in the cultures pretreated with the extract. These data suggest that the antigenotoxicity of propolis may occur under different mechanisms, including the antioxidant activity of the phenolic components present in the extract. | [189] |
| 2016 | Yalcinet al. | The objective was to evaluate the protective effect of ethanolic extract of propolis (EEP) against γ-ray-induced DNA damage on fibroblast cells. Initially, the cells were pretreated for 15 min and 30 min with three different concentrations of EEP (100 μg/mL, 200 μg/mL, and 300 μg/mL); subsequently, they were exposed to 3 Gyγ-rays. The results showed a significant decrease in γ-ray-induced DNA damage on cells treated with EEP. It was concluded that EEP might have radioprotective activity. | [190] |
| Micronucleus, chromosome aberrations, andcomet assayin vitro | |||
| 2008 | Benkovic et al. | Using three different cytogenetic tests, the protective effect of propolis and quercetin against damage to the genetic material induced by gamma radiation in a culture of human leukocytes was evaluated. The results suggested that both compounds have a low toxicity profile in in vitro conditions. Likewise, it was observed that they could be considered radioprotective agents, because they reduced the levels of DNA damage in leukocytes irradiated with gamma rays in the three types of tests evaluated. It was confirmed that propoleo was the most effective and that it can have different mechanisms of protective action. | [191] |
| 2009 | Benkovic et al. | This in vitro study aimed to evaluate the possible radioprotective effects of water-soluble derivate of propolis (WSDP), caffeic acid, chryrin, and naringin on gamma-irradiated human white blood cells. Using the same three tests as in the previous study, it was confirmed that none of the tested compounds induced significant genotoxicity; on the contrary, they offered a measurable protection against DNA damage. It was observed that the WSDP was the most effective in reducing levels of cytogenetic DNA damage in white blood cells. | [195] |
| Micronucleus and comet assay in vitro | |||
| 2016 | Roberto et al. | In this work, the genotoxic and antigenotoxic potential of the ethanolic extracts of Brazilian green propoleo and B. dracunculifolia in mammalian cells (HTC cells) was evaluated. After evaluating the exposure of the cells to each extract, individually and in combination with the MMS, the results showed that no extract was genotoxic; on the contrary, they exerted a significant reduction in DNA damage. The experiment carried out with a pre-incubation period was more effective than without the incubation test, showing that the tested extracts were able to inactivate the mutagen before it could react with the DNA. | [206] |
| Micronucleus and comet assay in vivo | |||
| 2013 | Oršolić et al. | The study was on the antitumor, chemopreventive, and immunostimulative effects of local chemoimmunotherapy and hyperthermal intraperitoneal chemotherapy (HIPEC) in a mouse-bearing Ehrlich ascites tumor (EAT). In this case, mice were treated with WSDP at a dose of 50 mg/kg, seven and three days before implantation of EAT cells, whereas the cisplatin (CIS) was injected three days after implantation of EAT cells. The combination of WSDP plus CIS resulted in tumor growth inhibition and increased the survival of mice at approximately 115%. The study also confirmed that WSDP reduced the cisplatin toxic and genotoxic effect to normal cells without affecting cisplatin cytotoxicity on EAT cells. | [196] |
| Micronucleus assay in vivo | |||
| 2010 | Türkez et al. | With the purpose of determining the effectiveness of propolis in the modulation of the genotoxicity induced by aluminum chloride (AlCl (3)) and the hepatotoxicity in the liver of rats, in a group of animals, a dose of the natural resin (50 mg/kg) plus AlCl (3) was administered orally daily for 30 days. At the end of the experiment, the histological alterations in the liver were investigated and their hepatocytes (HEP) were isolated to count the number of micronucleated hepatocytes (MNHEPs). This simultaneous treatment significantly modulated the pathological damages generated by aluminum chloride (sinusoidal dilatation, congestion of the central vein, lipid accumulation, and infiltration of lymphocytes) and reduced the frequency of MNHEPs. In conclusion, propolis may have an anticlastogenic effect and might antagonize the toxicity of AlCl (3). | [199] |
| 2010 | Oršolić et al. | Considering previous studies, where WSDP, quercetin, and naringin showed their effectiveness to reduce the DNA damage induced by gamma irradiation in white blood cells, the same group of researchers evaluated this property against the toxicity induced by irinotecan (antineoplastic derived from camptothecin and extracted from the Chinese tree, Camptotheca acuminata). On this occasion, they determined that propolis and some flavonoids could reduce the cyto and genotoxicity induced by irinotecan in cells of mice with Ehrlich ascites tumors (EAT); and at the same time, protect normal blood, liver, and kidney cells. The pre-treatment with propolis and/or flavonoides resulted in a substantial inhibition of the growth of EAT cells and significantly reduced the frequency of micronuclei. These results suggest that propolis have a significant immunomodulatory effect and can decrease the toxicity of the antineoplastic in normal cells. | [198] |
| 2014 | Dornelas et al. | The objective of this study was to evaluate the anticlastogenic effect of green propolis and L-lysine on the damage produced by the carcinogen, BBN [n-butyl-n (4-hydroxybutyl) nitrosamine], in erythroblasts (bone marrow) and leukocytes (peripheral blood). After 40 weeks of treatment with the evaluation compounds, the animals were anesthetized and subjected to femoral bone marrow aspiration and blood collection from the aorta to quantify the frequency of MN. The study confirmed that both propolis (150 mg/kg) and L-lysine (300 mg/kg), alone or in combination, were not genotoxic by themselves toward the cells evaluated; on the contrary, the dosage and timing were effective in protecting against the genotoxicity of BBN. | [207] |
| 2014 | Oršolić et al. | The analysis was on the inhibitory effect of an ethanolic extract of propolis (EEP) on the skin irritation, oxidative stress, and inflammatory response induced by n-Hexyl salycilate (HXS) or Di-n-propyl disulfide (PPD) in mice. The inflammation process was monitored by histopatological assessment of the skin, total number of inflammatory cells in the peritoneal cavity, macrophage-spreading index, and frequencies of micronucleated reticulocytes, lipid peroxidation, and glutathione assay in the skin. The topical application of EEP reduced the lipid peroxidation, the total number of inflammatory cells in the peritoneal cavity, and functional activity of macrophages. A significant decrease in the number of micronucleated reticulocytes was observed. These results demonstrate that a topical application of EEP may improve psoriatic-like skin lesions by suppressing the functional activity of macrophages and ROS production. | [197] |
| Micronucelus assay in vitro | |||
| 2012 | Türkez et al. | With the goal to determine the effectiveness of this natural resin in alleviating the 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)-induced toxicity in rat hepatocytes, three concentrations of propolis (25 μM, 50 μM, and 100 μM) were added in combination with TCDD to a primary culture of hepatocytes. The cell viability, total antioxidant capacity (TAC), levels of oxidative stress (TOS), and DNA damage were further quantified. The evidence indicated that in cultures treated with TCDD only, the cell viability and the level of TAC decreased, while the frequency of micronucleated hepatocytes (MNHEPs) increased. A contrary situation was present in cultures with a combination of TCDD and propolis, in which the resin modulated its toxic effects and significantly reduced the number of MNHEPs. | [200] |
| 2014 | Santos et al. | Brazilian researchers evaluated the effect of propolis (type AF-08) on the genotoxicity, cytotoxicity, and clonogenic death of Chinese hamster ovary (CHO-K1) cells irradiated with (60) Co gamma-radiation. The MN test revealed that AF-08 alone (5 μg/mL–100μg/mL) was not genotoxic and that a low concentration of it (30μg/mL) reduced the damage induced by radiation in the DNA. On the other hand, analysis of cytotoxicity showed that a concentration of 50μg/mL presented a significant proliferative effect when associated with radiation, decreasing the percentage of necrotic cells. Concerning the clonogenic capacity, AF-08 also evidenced a significant stimulating effect on cell proliferation. Together, these data suggest that propolis AF-08 might be potentially radioprotective. | [208] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Romero, D.; Izquierdo-Vega, J.A.; Morales-González, J.A.; Madrigal-Bujaidar, E.; Chamorro-Cevallos, G.; Sánchez-Gutiérrez, M.; Betanzos-Cabrera, G.; Alvarez-Gonzalez, I.; Morales-González, Á.; Madrigal-Santillán, E. Evidence of Some Natural Products with Antigenotoxic Effects. Part 2: Plants, Vegetables, and Natural Resin. Nutrients 2018, 10, 1954. https://doi.org/10.3390/nu10121954
López-Romero D, Izquierdo-Vega JA, Morales-González JA, Madrigal-Bujaidar E, Chamorro-Cevallos G, Sánchez-Gutiérrez M, Betanzos-Cabrera G, Alvarez-Gonzalez I, Morales-González Á, Madrigal-Santillán E. Evidence of Some Natural Products with Antigenotoxic Effects. Part 2: Plants, Vegetables, and Natural Resin. Nutrients. 2018; 10(12):1954. https://doi.org/10.3390/nu10121954
Chicago/Turabian StyleLópez-Romero, David, Jeannett A. Izquierdo-Vega, José Antonio Morales-González, Eduardo Madrigal-Bujaidar, Germán Chamorro-Cevallos, Manuel Sánchez-Gutiérrez, Gabriel Betanzos-Cabrera, Isela Alvarez-Gonzalez, Ángel Morales-González, and Eduardo Madrigal-Santillán. 2018. "Evidence of Some Natural Products with Antigenotoxic Effects. Part 2: Plants, Vegetables, and Natural Resin" Nutrients 10, no. 12: 1954. https://doi.org/10.3390/nu10121954
APA StyleLópez-Romero, D., Izquierdo-Vega, J. A., Morales-González, J. A., Madrigal-Bujaidar, E., Chamorro-Cevallos, G., Sánchez-Gutiérrez, M., Betanzos-Cabrera, G., Alvarez-Gonzalez, I., Morales-González, Á., & Madrigal-Santillán, E. (2018). Evidence of Some Natural Products with Antigenotoxic Effects. Part 2: Plants, Vegetables, and Natural Resin. Nutrients, 10(12), 1954. https://doi.org/10.3390/nu10121954