Polyphenols in Liubao Tea Can Prevent CCl4-Induced Hepatic Damage in Mice through Its Antioxidant Capacities


Abstract
:1. Introduction
2. Materials and Methods
2.1. PLT Extraction
2.2. Experimental Model in Kunming (KM) Mice
2.3. Measurement of Serum Indices
2.4. Cytokine Levels in Serum
2.5. Histopathological Analysis of Liver Tissues
2.6. qPCR Analysis
2.7. Western Blot Analysis
2.8. High-Performance Liquid Chromatography (HPLC) Assay
2.9. Statistical Analysis
3. Results
3.1. Body Weight, Liver Weight, and Liver Indices of the Experimental Mice
3.2. Serum AST, ALT, and TG Levels
3.3. Serum SOD, GSH-Px, and MDA Levels
3.4. Serum IL-6, IL-12, TNF-α, and IFN-γ Cytokine Levels
3.5. Histopathological Assessment of the Liver
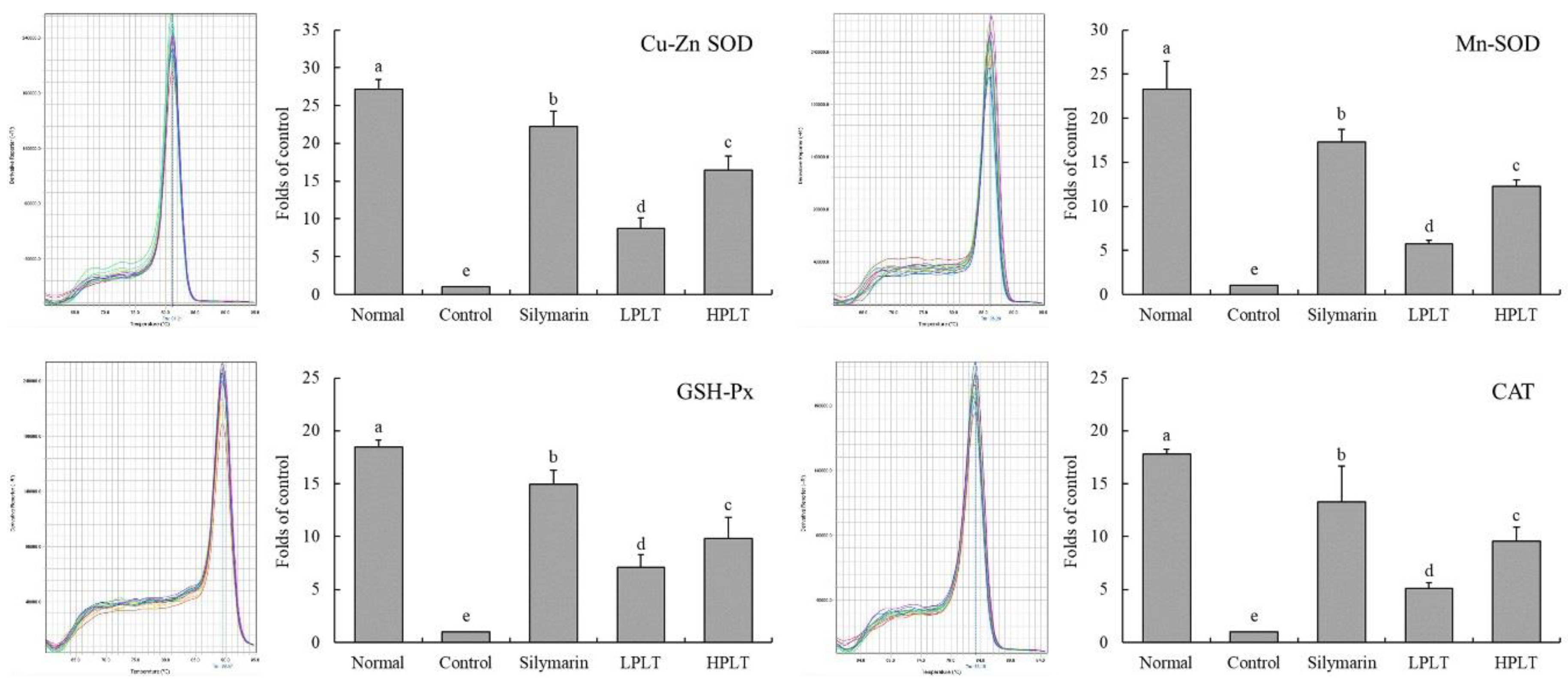
3.6. mRNA and Protein Expression of Cu/Zn-SOD, Mn-SOD, GSH-Px, and CAT in Mouse Hepatic Tissues
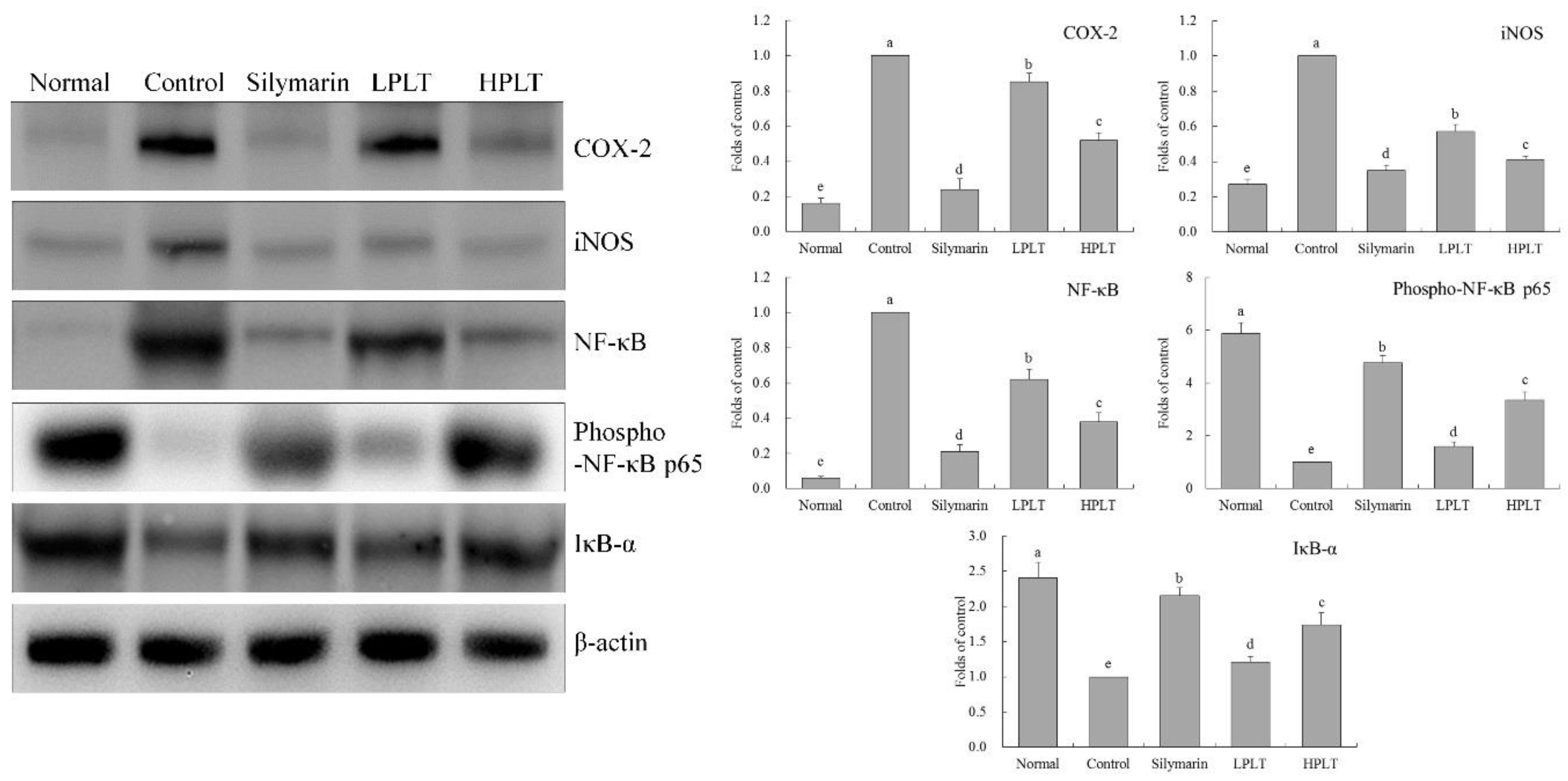
3.7. mRNA and Protein Expression of COX-2, iNOS, NF-κB, and IκB-α in Mouse Hepatic Tissues
3.8. Protein Expression of Cytochrome P450 Reductase in Mouse Hepatic Tissues
3.9. Constituents of PTL
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Liu, X.L.; Li, Y.; Jiang, Y.X.; Yu, X. Analysis on feature composition of Guangxi Liubao tea. J. Beijing Technol. Bus. Univ. (Nat. Sci. Ed.) 2012, 30, 46–50. [Google Scholar]
- Huang, L.; Peng, J.J.; Xia, N.; Teng, J.W.; Wei, B.Y. Effect on regulation of hyperlipidemia and anticoagulant for Liupu tea. Food. Sci. Technol. 2013, 38, 123–127. [Google Scholar]
- Liu, J.Q.; Shao, W.F.; Zhao, B.Q.; Zhao, B.; Wang, H.Q.; Ma, X.M.; Guo, T.; Hou, Y. Study of fermented pu-erh tea powder, dark tea powder and liupu tea on losing weight in hyperlipidemia model rats. China J. Tradi. Chin. Med. Pharm. 2014, 29, 108–112. [Google Scholar]
- Zhang, X.Y.; Huang, Y.S.; Liu, G.P.; Rao, W.Y.; Qin, L.; Deng, Y.Y. The effects of Liubao tea on blood lipid and antioxidation and hyperlipidemia mice. J. Med. Theory Pract. 2013, 26, 563–564. [Google Scholar]
- Zhao, Y.Y.; Huang, L.; Wei, B.Y.; Teng, J.W.; Xia, N. Effect of Liupao tea extract on fecal microbiota in hyperlipidemic mice. Sci. Technol. Food Ind. 2015, 36, 364–367. [Google Scholar]
- Teng, Q.Q.; Liu, Z.H.; Gong, S.J.; Peng, Y.X.; Ma, R. Effect of Liupao Tea on glucose and lipid metabolism in palmitate-induced insulin resistance 3T3-L1 adipocytes. J. Tea Sci. 2014, 34, 230–238. [Google Scholar]
- Xu, B.B.; Li, Y.L.; Wang, B.Y. Research advances in risk factors for alcoholic liver disease. Zhonghua Gan Zang Bing Za Zhi 2017, 25, 397–400. [Google Scholar] [PubMed]
- Burk, R.F.; Lane, J.M.; Patel, K. Relationship of oxygen and glutathione in protection against carbon tetrachloride-induced hepatic microsomal lipid peroxidation and covalent binding in the rat. Rationale for the use of hyperbaric oxygen to treat carbon tetrachloride ingestion. J. Clin. Investig. 1984, 74, 1996–2001. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Cai, G.M.; He, Q. Oxidative stress and hepatic injury. World Chin. J. Digestol. 2008, 16, 3310–3315. [Google Scholar] [CrossRef]
- Kayesh, M.E.H.; Ezzikouri, S.; Sanada, T.; Chi, H.; Hayashi, Y.; Rebbani, K.; Kitab, B.; Matsuu, A.; Miyoshi, N.; Hishima, T.; et al. Oxidative stress and immune responses during hepatitis C virus infection in tupaia belangeri. Sci. Rep. 2017, 7, 9848. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.; Rashid, S.; Nafees, S.; Hasan, S.K.; Shahid, A.; Majed, F.; Sultana, S. Protective effect of Chlorogenic acid against methotrexate induced oxidative stress, inflammation and apoptosis in rat liver: An experimental approach. Chem. Biol. Interact. 2017, 272, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Zhang, X.; Han, J.; Man, K.; Zhang, Y.; Chu, E.S.; Nan, Y.; Yu, J. Pro-inflammatory CXCR3 impairs mitochondrial function in experimental non-alcoholic steatohepatitis. Theranostics 2017, 7, 4192–4203. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, S.; Ghosh, S.; Mukherjee, S.; Gupta, P.; Bhattacharya, S.; Adhikary, A.; Chattopadhyay, S. Pomegranate protects against arsenic-induced p53-dependent ROS-mediated inflammation and apoptosis in liver cells. J. Nutr. Biochem. 2016, 38, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Niedzwiecki, A.; Roomi, M.W.; Kalinovsky, T.; Rath, M. Anticancer efficacy of polyphenols and their combinations. Nutrients 2016, 8, E552. [Google Scholar] [CrossRef] [PubMed]
- Megow, I.; Darvin, M.E.; Meinke, M.C.; Lademann, J. A randomized controlled trial of green tea beverages on the in vivo radical scavenging activity in human skin. Skin Pharmacol. Physiol. 2017, 30, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Yi, R.; Wang, R.; Sun, P.; Zhao, X. Antioxidant-mediated preventative effect of Dragon-pearl tea crude polyphenol extract on reserpine-induced gastric ulcers. Exp. Ther. Med. 2015, 10, 338–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, G.J.; Gong, Z.J.; Sun, X.M.; Zheng, S.H.; Li, X. Tea polyphenols inhibit expressions of iNOS and TNF-alpha and prevent lipopolysaccharide-induced liver injury in rats. Hepatobiliary Pancreat. Dis. Int. 2006, 5, 262–267. [Google Scholar] [PubMed]
- Salomone, F.; Godos, J.; Zelber-Sagi, S. Natural antioxidants for non-alcoholic fatty liver disease: Molecular targets and clinical perspectives. Liver Int. 2016, 36, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Q.; Ye, Y.; Cheng, H.; Tang, D.D.; Su, L.H. Studies on physical and chemical properties of Liubao tea. Chin. Agri. Sci. Bull. 2008, 24, 77–80. [Google Scholar]
- Zhao, X.; Qian, Y. Preventive effects of Kuding tea crude polyphenols in DSS-induced C57BL/6J mice ulcerative colitis. Sci. Technol. Food Ind. 2017, 38, 357–362. [Google Scholar]
- Zhao, X.; Qian, Y.; Li, G.J.; Tan, J. Preventive effects of the polysaccharide of Larimichthys crocea swim bladder on carbon tetrachloride (CCl4)-induced hepatic damage. Chin. J. Nat. Med. 2015, 13, 521–528. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, X.; Wang, H.; Yang, Z.; Li, J.; Suo, H. Prevent effects of Lactobacillus fermentum HY01 on dextran sulfate sodium-induced colitis in mice. Nutrients 2017, 9, E545. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Cheng, Q.; Qian, Y.; Yi, R.K.; Gu, L.J.; Wang, S.S.; Song, J.L. Insect tea attenuates hydrochloric acid and ethanol-induced mice acute gastric injury. Exp. Ther. Med. 2017, 14, 5135–5142. [Google Scholar] [CrossRef] [PubMed]
- Fan, D.M.; Fan, K.; Yu, C.P.; Lu, Y.T.; Wang, X.C. Tea polyphenols dominate the short-term tea (Camellia sinensis) leaf litter decomposition. J. Zhejiang Univ. Sci. B 2017, 18, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.L.; Lin, Y.; Liu, Z.H.; Huang, J.A.; Long, Z.R.; Teng, C.Q.; Ma, S.C.; Qiu, R.J.; Cao, Z.H. Research on acute and subacute toxicity evaluation of Liupao tea. J. Tea Sci. 2017, 37, 173–181. [Google Scholar]
- Wu, H.; Qiu, Y.; Shu, Z.; Zhang, X.; Li, R.; Liu, S.; Chen, L.; Liu, H.; Chen, N. Protective effect of Trillium tschonoskii saponin on CCl4-induced acute liver injury of rats through apoptosis inhibition. Can. J. Physiol. Pharmacol. 2016, 94, 1291–1297. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.M.; Abdelrahman, S.A.; Salama, A.E. Efficacy of gold nanoparticles against isoproterenol induced acute myocardial infarction in adult male albino rats. Ultrastruct. Pathol. 2017, 41, 168–185. [Google Scholar] [CrossRef] [PubMed]
- Maksymchuk, O.; Shysh, A.; Rosohatska, I.; Chashchyn, M. Quercetin prevents type 1 diabetic liver damage through inhibition of CYP2E1. Pharmacol. Rep. 2017, 69, 1386–1392. [Google Scholar] [CrossRef] [PubMed]
- Mahmoodzadeh, Y.; Mazani, M.; Rezagholizadeh, L. Hepatoprotective effect of methanolic Tanacetum parthenium extract on CCl4-induced liver damage in rats. Pharmacol. Rep. 2017, 4, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Su, M.; Chen, Q.; Chang, Q.; Wang, W.; Li, H. Protective effect of a polysaccharide from Anoectochilus roxburghii against carbon tetrachloride-induced acute liver injury in mice. J. Ethnopharmacol. 2017, 200, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Dreher, D.; Junod, A.F. Differential effects of superoxide, hydrogen peroxide, and hydroxyl radical on intracellular calcium in human endothelial cells. J Cell Physiol. 1995, 162, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; You, G.; Xu, Y.; Wang, C.; Wang, P.; Miao, L.; Dai, S.; Lv, B.; Yang, Y. Antioxidant enzyme activities as biomarkers of fluvial biofilm to ZnO NPs ecotoxicity and the Integrated Biomarker Responses (IBR) assessment. Ecotoxicol. Environ. Saf. 2016, 133, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.; Ma, M.; Han, X.; Wang, Z.; Li, H. Hyperin protects against cisplatin-induced liver injury in mice. Acta Cir. Bras. 2017, 32, 633–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Won, G.; Kim, T.H.; Lee, J.H. A novel Salmonella strain inactivated by a regulated autolysis system and expressing the B subunit of Shiga toxin 2e efficiently elicits immune responses and confers protection against virulent Stx2e-producing Escherichia coli. BMC Vet. Res. 2017, 13, 40. [Google Scholar] [CrossRef] [PubMed]
- Kampan, N.C.; Madondo, M.T.; McNally, O.M.; Stephens, A.N.; Quinn, M.A.; Plebanski, M. Interleukin 6 present in inflammatory ascites from advanced epithelial ovarian cancer patients promotes tumor necrosis factor receptor 2-expressing regulatory T. cells. Front. Immunol. 2017, 8, 1482. [Google Scholar] [CrossRef] [PubMed]
- Gil-Farina, I.; Di Scala, M.; Salido, E.; López-Franco, E.; Rodríguez-García, E.; Blasi, M.; Merino, J.; Aldabe, R.; Prieto, J.; Gonzalez-Aseguinolaza, G. Transient expression of transgenic IL-12 in mouse liver triggers unremitting inflammation mimicking human autoimmune hepatitis. J. Immunol. 2016, 197, 2145–2156. [Google Scholar] [CrossRef] [PubMed]
- Erkasap, S.; Erkasap, N.; Bradford, B.; Mamedova, L.; Uysal, O.; Ozkurt, M.; Ozyurt, R.; Kutlay, O.; Bayram, B. The effect of leptin and resveratrol on JAK/STAT pathways and Sirt-1 gene expression in the renal tissue of ischemia/reperfusion induced rats. Bratisl. Lek. Listy 2017, 118, 443–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Chen, M.J.; Wang, C.; Nie, H.; Huang, W.J.; Yuan, T.D.; Sun, T.; Shu, K.G.; Wang, C.F.; Gong, Q.; et al. Protective effects of hesperidin on concanavalin A-induced hepatic injury in mice. Int. Immunopharmacol. 2014, 21, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Li, Y.; Duan, H.; Sivakumar, R.; Li, X. Chronic exposure of nanomolar MC-LR caused oxidative stress and inflammatory responses in HepG2 cells. Chemosphere 2018, 192, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.X.; Han, X.M.; Jiang, X.W.; Tao, J. The Effects of two different multivitamins on aging mice. Chin. J. Physiol. 2017, 60, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Najafpour, M.M. A possible evolutionary origin for the Mn4 cluster in photosystem II: From manganese superoxide dismutase to oxygen evolving complex. Orig. Life Evol. Biosph. 2009, 32, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Suo, H.; Feng, X.; Zhu, K.; Wang, C.; Zhao, X.; Kan, J. Shuidouchi (fermented soybean) fermented in different vessels attenuates HCl/ethanol-induced gastric mucosal injury. Molecules 2015, 20, 19748–19763. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, X.J.; Feng, R.; Jiang, Y.; Zhang, D.Y.; He, C.; Li, P.; Wan, J.B. Hepatoprotective properties of Penthorum chinense Pursh against carbon tetrachloride-induced acute liver injury in mice. Chin. Med. 2017, 12, 32. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, M.H.; Wefers, H.; Sies, H. Generation of excited species catalyzed by horseradish peroxidase or hemin in the presence of reduced glutathione and H2O2. Free Radic. Biol. Med. 1987, 3, 107–110. [Google Scholar] [CrossRef]
- Ma, Q.; Liu, C.M.; Qin, Z.H.; Jiang, J.H.; Sun, Y.Z. Ganoderma applanatum terpenes protect mouse liver against benzo(α)pyren-induced oxidative stress and inflammation. Environ. Toxicol. Pharmacol. 2011, 31, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.M.; Zheng, Y.L.; Lu, J.; Zhang, Z.F.; Fan, S.H.; Wu, D.M.; Ma, J.Q. Quercetin protects rat liver against lead-induced oxidative stress and apoptosis. Environ. Toxicol. Pharmacol. 2010, 29, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Alam, M.S.; Jabbar, Z.; Javed, K.; Athar, M. Evaluation of antioxidant activity of Cassia siamea flowers. J. Ethnopharmacol. 2006, 108, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.; Liu, C.; Chiang, F.Y.; Wang, L.F.; Lee, K.W.; Chen, W.T.; Kuo, P.L.; Liang, C.H. IL-8 promotes inflammatory mediators and stimulates activation of p38 MAPK/ERK-NF-κB pathway and reduction of JNK in HNSCC. Oncotarget 2017, 8, 56375–56388. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Chen, W.N.; Li, R.; Hu, C.M.; Lei, C.; Li, C.M. Therapeutic effect of acetazolamide, an aquaporin 1 inhibitor, on adjuvant-induced arthritis in rats by inhibiting NF-κB signal pathway. Immunopharmacol. Immunotoxicol. 2018, 40, 117–125. [Google Scholar] [CrossRef] [PubMed]
- El-Gohary, A. Obestatin improves hepatic injury induced by ischemia/reperfusion in rats: Role of nitric oxide. Gen. Physiol. Biophys. 2017, 36, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, M.; Waibler, Z.; Pleli, T.; Pfeilschifter, J.; Mühl, H. Type I interferon supports inducible nitric oxide synthase in murine hepatoma cells and hepatocytes and during experimental acetaminophen-induced liver damage. Front. Immunol. 2017, 8, 890. [Google Scholar] [CrossRef] [PubMed]
- Araújo Júnior, R.F.; Garcia, V.B.; Leitão, R.F.; Brito, G.A.; Miguel Ede, C.; Guedes, P.M.; de Araújo, A.A. Carvedilol improves inflammatory response, oxidative stress and fibrosis in the alcohol-induced liver injury in rats by regulating Kuppfer cells and hepatic stellate cells. PLoS ONE 2016, 12, e0148868. [Google Scholar] [CrossRef] [PubMed]
- Turesky, R.J.; Konorev, D.; Fan, X.; Tang, Y.; Yao, L.; Ding, X.; Xie, F.; Zhu, Y.; Zhang, Q.Y. Effect of cytochrome P450 reductase deficiency on 2-amino-9H-pyrido[2,3-b]indole metabolism and DNA adduct formation in liver and extrahepatic tissues of mice. Chem. Res. Toxicol. 2015, 28, 2400–2410. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, S.C.; Wu, C.H.; Wu, C.C.; Yen, J.H.; Liu, M.C.; Hsueh, C.M.; Hsu, S.L. Gallic acid selectively induces the necrosis of activated hepatic stellate cells via a calcium-dependent calpain I. activation pathway. Life Sci. 2014, 102, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.H.; Yang, J.; Yang, Y.H. Research progress on pharmacological effects of gallic acid. Chin. Hosp. Pharm. J. 2017, 37, 94–98. [Google Scholar]
- Zhai, W.; Zheng, J.H.; Yao, X.D.; Peng, B.; Liu, M.; Huang, J.H.; Wang, G.C.; Xu, Y.F. Catechin prevents the calcium oxalate monohydrate induced renal calcium crystallization in NRK-52E cells and the ethylene glycol induced renal stone formation in rat. BMC Complem. Altern. Med. 2013, 13, 228. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Yamamoto, S.; Hirayama, C.; Takino, T.; Fujisawa, K.; Oda, T. Cianidanol therapy for HBe-antigen-positive chronic hepatitis: A multicentre, double-blind study. Liver 1986, 6, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Ramadori, G.; Sipe, J.D.; Dinarello, C.A.; Mizel, S.B.; Colten, H.R. Pretranslational modulation of acute phase hepatic protein synthesis by murine recombinant interleukin 1 (IL-1) and purified human IL-1. J. Exp. Med. 1995, 162, 930–942. [Google Scholar] [CrossRef]
- Furman, D.; Chang, J.; Lartigue, L.; Bolen, C.R.; Haddad, F.; Gaudilliere, B.; Ganio, E.A.; Fragiadakis, G.K.; Spitzer, M.H.; Douchet, I.; et al. Expression of specific inflammasome gene modules stratifies older individuals into two extreme clinical and immunological states. Nat. Med. 2017, 23, 174–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Lv, X.W.; Li, J.; Zhang, L.; Liu, H.F.; Huang, C.; Zhu, P.L. Protective effect of caffeine on alcohol-induce acute liver injury in mice. Acta Univ. Med. Anhui 2009, 44, 359–362. [Google Scholar]
- Kim, A. Mechanisms underlying beneficial health effects of tea catechins to improve insulin resistance and endothelial dysfunction. Endocr. Metab. Immune Disord. Drug Targets 2008, 8, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, R.H.; Karam, R.A.; Amer, M.G. Epicatechin attenuates doxorubicin-induced brain toxicity: Critical role of TNF-α, iNOS and NF-κB. Brain Res. Bull. 2011, 86, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Zhen, C.; Wang, X.M.; Yin, Z.Y.; Wang, Q.; Liu, P.G.; Wu, G.Y.; Yu, K.K.; Li, G.S. Effect of EGCG on expression of TGF-β1 and CTGF in rats with liver fibrosis. World Chin. J. Digestol. 2008, 16, 3828–3834. [Google Scholar] [CrossRef]
- Liu, D.M.; Wang, X.F. EGCG influences the NF-κB and ICAM-I expression of ConA inducing liver injury. Anat. Res. 2014, 36, 27–30. [Google Scholar]
- Ye, J.X.; Wang, L.; Liang, R.X.; Yang, B. Protection and its mechanism of catechin morphon on hypoxia-reoxynation induced in myocardial cells. China J. Chin. Mater. Med. 2008, 33, 801–805. [Google Scholar]
- Huang, X.; Zhong, W.B.; Huang, S.L.; Hu, Y.H.; Wang, D.D.; Sun, Y. Anti-tumor activities of catechins EGCG and ECG against human hepatocarcinoma BEL-7402 cells. J. Guangdong Pharm. Univ. 2013, 29, 435–438. [Google Scholar]
- Zhang, X.N.; Lin, Y.; Huang, J.A.; Liu, Z.H.; Liang, D.D. Inhibitory effects of tea extracts EGCG, GCG and ECG on the melanogenesis in melanoma cell B16. J. Hunan Agric. Univ. (Nat. Sci.) 2017, 43, 405–410. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grade | Portal Area and Surrounding Area | Hepatic Lobule |
|---|---|---|
| 0 | No inflammation | No inflammation |
| 1 | Portal inflammation | Degeneration and few necrotic foci |
| 2 | Mild detrital necrosis | Degeneration, focal necrosis |
| 3 | Moderate detrital necrosis | Degeneration or necrosis, or bridge necrosis |
| 4 | Severe detrital necrosis | Bridge necrosis wide range, involving multiple lobules, leaflet structure disorder |
| Gene | Forward Sequence | Reverse Sequence |
|---|---|---|
| COX-2 | 5′–GGTGCCTGGTCTGATGATG–3′ | 5′–TGCTGGTTTGGAATAGTTGCT–3′ |
| iNOS | 5′–GTTCTCAGCCCAACAATACAAGA–3′ | 5′–GTGGACGGGTCGATGTCAC–3 |
| NF-κB | 5′–ATGGCAGACGATGATCCCTAC–3′ | 5′–CGGAATCGAAATCCCCTCTGTT–3′ |
| IκB-α | 5′–TGAAGGACGAGGAGTACGAGC–3′ | 5′–TGCAGGAACGAGTCTCCGT–3′ |
| Cu/Zn-OD | 5′–AACCAGTTGTGTTGTCAGGAC–3′ | 5′–CCACCATGTTTCTTAGAGTGAGG–3′ |
| Mn-SOD | 5′–CAGACCTGCCTTACGACTATGG–3′ | 5′–CTCGGTGGCGTTGAGATTGTT–3′ |
| GSH-Px | 5′–CCACCGTGTATGCCTTCTCC–3′ | 5′–AGAGAGACGCGACATTCTCAAT–3′ |
| CAT | 5′–GGAGGCGGGAACCCAATAG–3′ | 5′–GTGTGCCATCTCGTCAGTGAA–3′ |
| GAPDH | 5′–AGGTCGGTGTGAACGGATTTG–3′ | 5′–GGGGTCGTTGATGGCAACA–3′ |
| t/min | A/% | B/% |
|---|---|---|
| 0 | 10 | 90 |
| 6.5 | 18.5 | 81.5 |
| 20 | 29.5 | 70.5 |
| Group | 1st Day Body Weight (g) | 14th Day Body Weight (g) | 15th Day Body Weight (g) | Liver Weight (g) | Liver Index |
|---|---|---|---|---|---|
| Normal | 35.17 ± 0.32 a | 42.66 ± 1.59 b | 42.12 ± 1.96 b | 1.57 ± 0.07 b | 3.73 ± 0.18 e |
| Control | 35.53 ± 0.28 a | 48.22 ± 2.62 a | 46.64 ± 1.38 a | 2.41 ± 0.18 a | 5.15 ± 0.23 a |
| Silymarin | 35.06 ± 0.22 a | 41.43 ± 0.55 b | 40.35 ± 0.84 b | 1.58 ± 0.08 b | 3.92 ± 0.19 d |
| LPLT | 35.19 ± 0.26 a | 36.62 ± 1.45 c | 35.61 ± 1.81 c | 1.61 ± 0.12 b | 4.51 ± 0.11 b |
| HPLT | 35.41 ± 0.20 a | 36.63 ± 2.41 c | 35.84 ± 3.70 c | 1.48 ± 0.15 b | 4.14 ± 0.03 c |
| Group | AST (U/L) | ALT (U/L) | TG (pg/mL) |
|---|---|---|---|
| Normal | 6.20 ± 0.43 e | 1.54 ± 0.22 e | 150.00 ± 26.15 e |
| Control | 21.13 ± 0.93 a | 17.98 ± 1.53 a | 563.75 ± 16.18 a |
| Silymarin | 12.13 ± 0.35 d | 4.09 ± 0.44 d | 208.75 ± 20.06 d |
| LPLT | 17.75 ± 0.57 b | 12.36 ± 2.23 b | 385.00 ± 57.72 b |
| HPLT | 13.85 ± 0.55 c | 8.45 ± 0.64 c | 273.75 ± 32.89 c |
| Group | SOD (U/mL) | GSH-Px (U/mL) | MDA (nmol/mL) |
|---|---|---|---|
| Normal | 121.38 ± 4.88 a | 85.92 ± 1.83 a | 2.24 ± 0.06 e |
| Control | 58.56 ± 2.42 d | 5.52 ± 1.02 e | 5.93 ± 0.45 a |
| Silymarin | 107.11 ± 1.77 b | 63.94 ± 3.20 b | 2.89 ± 0.16 d |
| LPLT | 78.44 ± 8.35 c | 25.35 ± 1.03 d | 4.44 ± 0.21 b |
| HPLT | 106.12 ± 1.37 b | 53.49 ± 2.84 c | 3.34 ± 0.26 c |
| Group | IL-6 (pg/mL) | IL-12 (pg/mL) | TNF-α (pg/mL) | IFN-γ (pg/mL) |
|---|---|---|---|---|
| Normal | 31.11 ± 1.84 d | 26.17 ± 3.06 d | 365.40 ± 16.75 e | 32.32 ± 0.59 d |
| Control | 64.33 ± 3.80 a | 56.68 ± 6.98 a | 718.76 ± 40.28 a | 77.94 ± 1.60 a |
| Silymarin | 41.02 ± 3.01 c | 34.50 ± 1.90 c | 467.22 ± 38.47 d | 39.07 ± 0.27 c |
| LPLT | 54.27 ± 6.05 b | 42.46 ± 4.92 b | 622.90 ± 50.68 b | 44.61 ± 0.79 b |
| HPLT | 45.19 ± 1.24 c | 36.69 ± 0.55 bc | 547.63 ± 26.83 c | 39.10 ± 0.56 c |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, Y.; Long, X.; Yi, R.; Zhao, X. Polyphenols in Liubao Tea Can Prevent CCl4-Induced Hepatic Damage in Mice through Its Antioxidant Capacities. Nutrients 2018, 10, 1280. https://doi.org/10.3390/nu10091280
Pan Y, Long X, Yi R, Zhao X. Polyphenols in Liubao Tea Can Prevent CCl4-Induced Hepatic Damage in Mice through Its Antioxidant Capacities. Nutrients. 2018; 10(9):1280. https://doi.org/10.3390/nu10091280
Chicago/Turabian StylePan, Yanni, Xingyao Long, Ruokun Yi, and Xin Zhao. 2018. "Polyphenols in Liubao Tea Can Prevent CCl4-Induced Hepatic Damage in Mice through Its Antioxidant Capacities" Nutrients 10, no. 9: 1280. https://doi.org/10.3390/nu10091280
APA StylePan, Y., Long, X., Yi, R., & Zhao, X. (2018). Polyphenols in Liubao Tea Can Prevent CCl4-Induced Hepatic Damage in Mice through Its Antioxidant Capacities. Nutrients, 10(9), 1280. https://doi.org/10.3390/nu10091280