Release of Major Peanut Allergens from Their Matrix under Various pH and Simulated Saliva Conditions—Ara h2 and Ara h6 Are Readily Bio-Accessible

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Peanut Materials
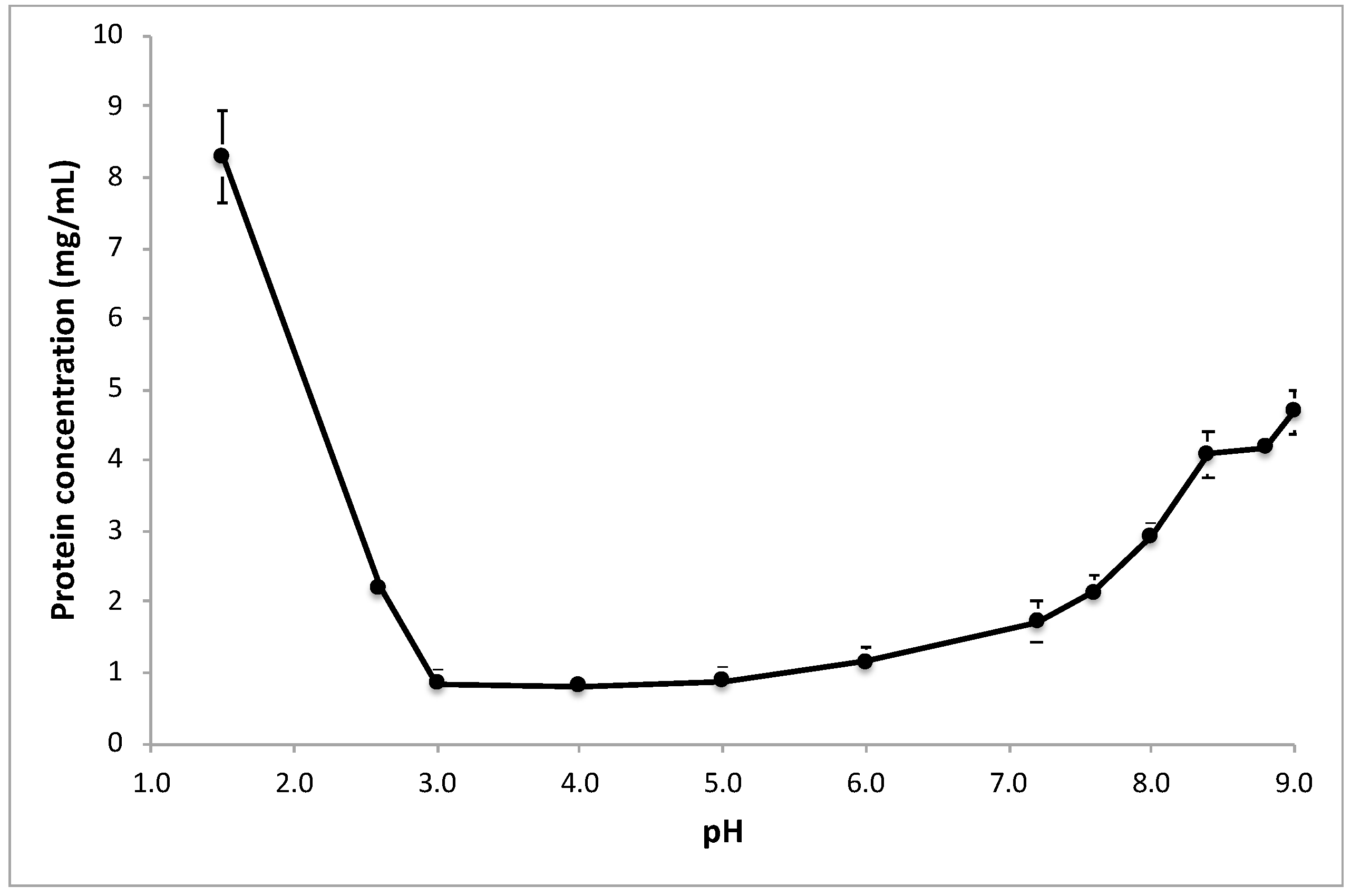
2.2. Screening Peanut Protein Solubility in a Wide pH Range (pH 1.5 to 9.0)
2.3. Testing Peanut Protein Solubility in Saliva Conditions
3. Results and Discussion
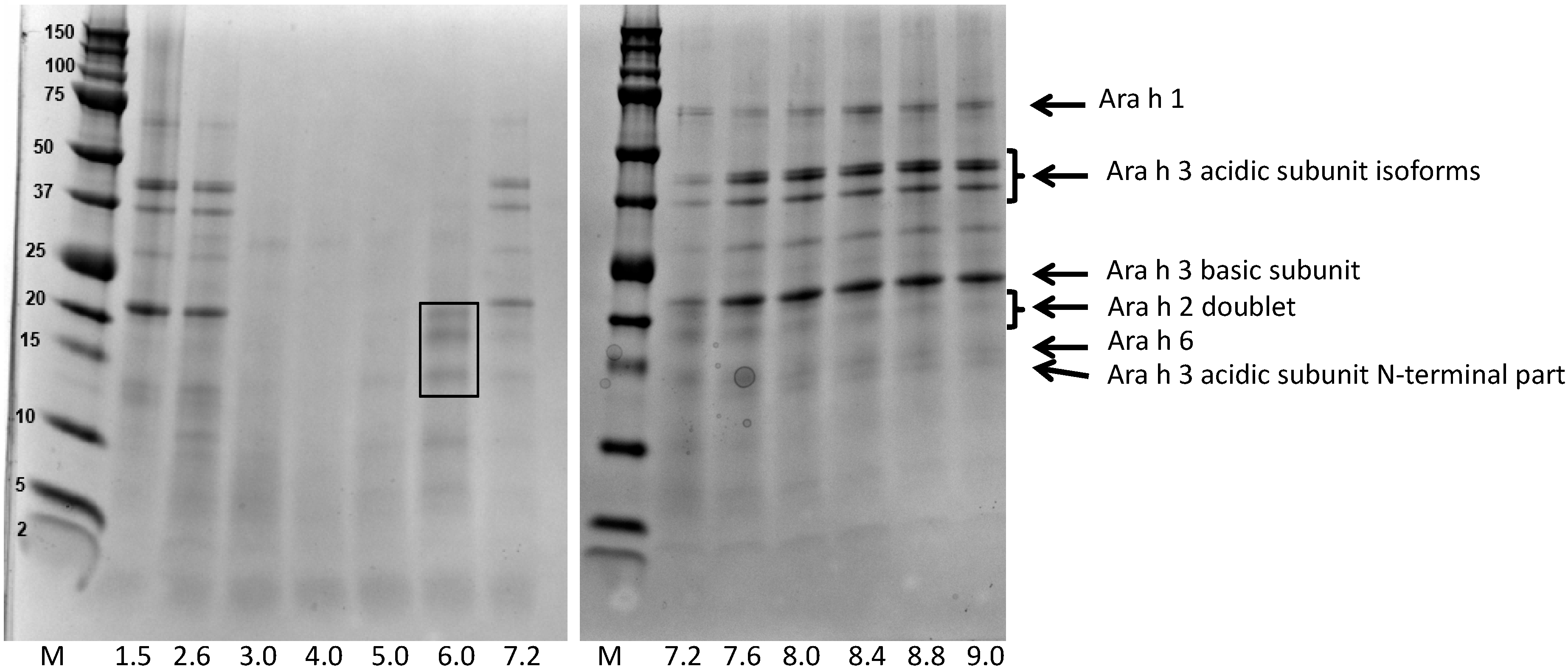
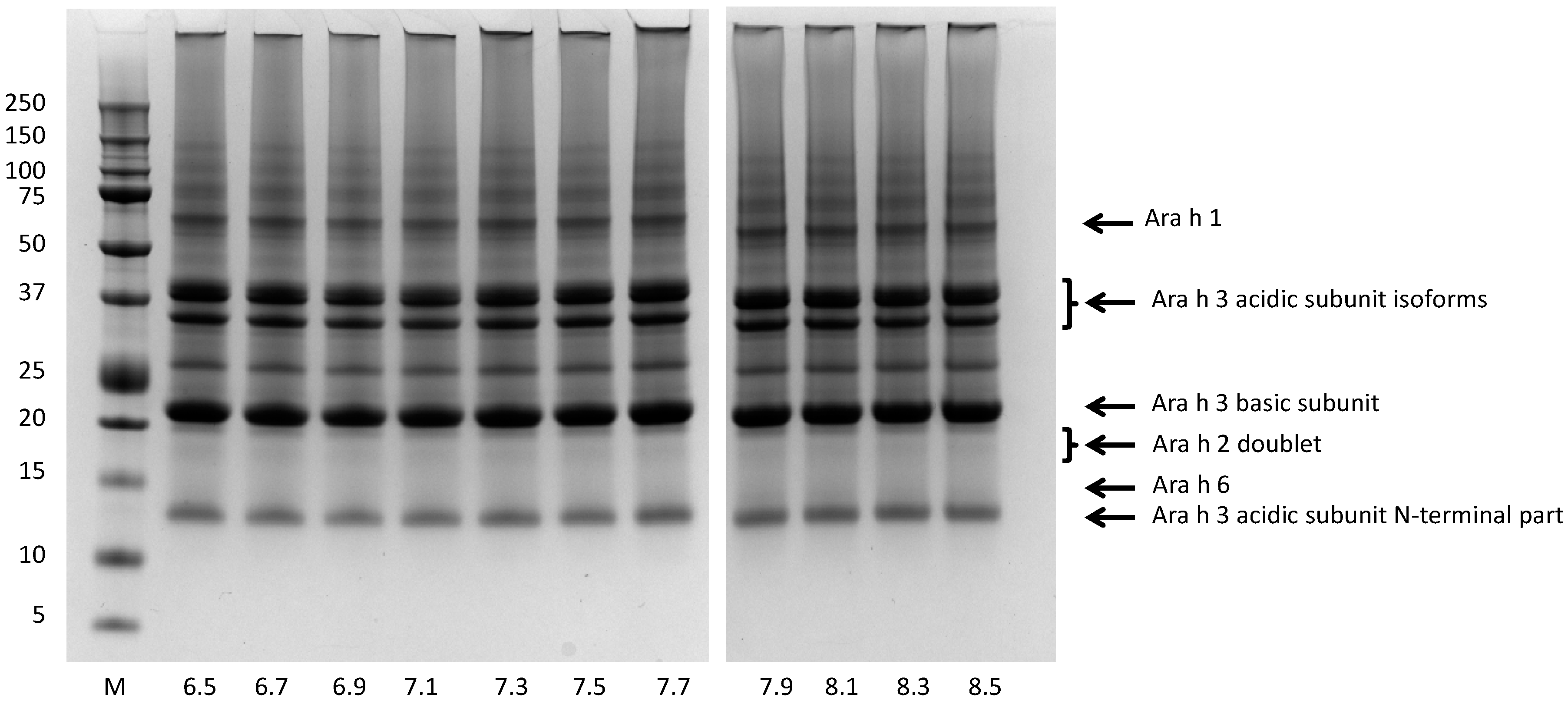
3.1. Screening the pH Effect on the Efficiency of Peanut Protein Extraction
3.2. Bio-Accessibility of Peanut Allergens in Artificial Saliva
3.3. The pH of the Extraction Medium Can Be Influenced by Peanut Flour
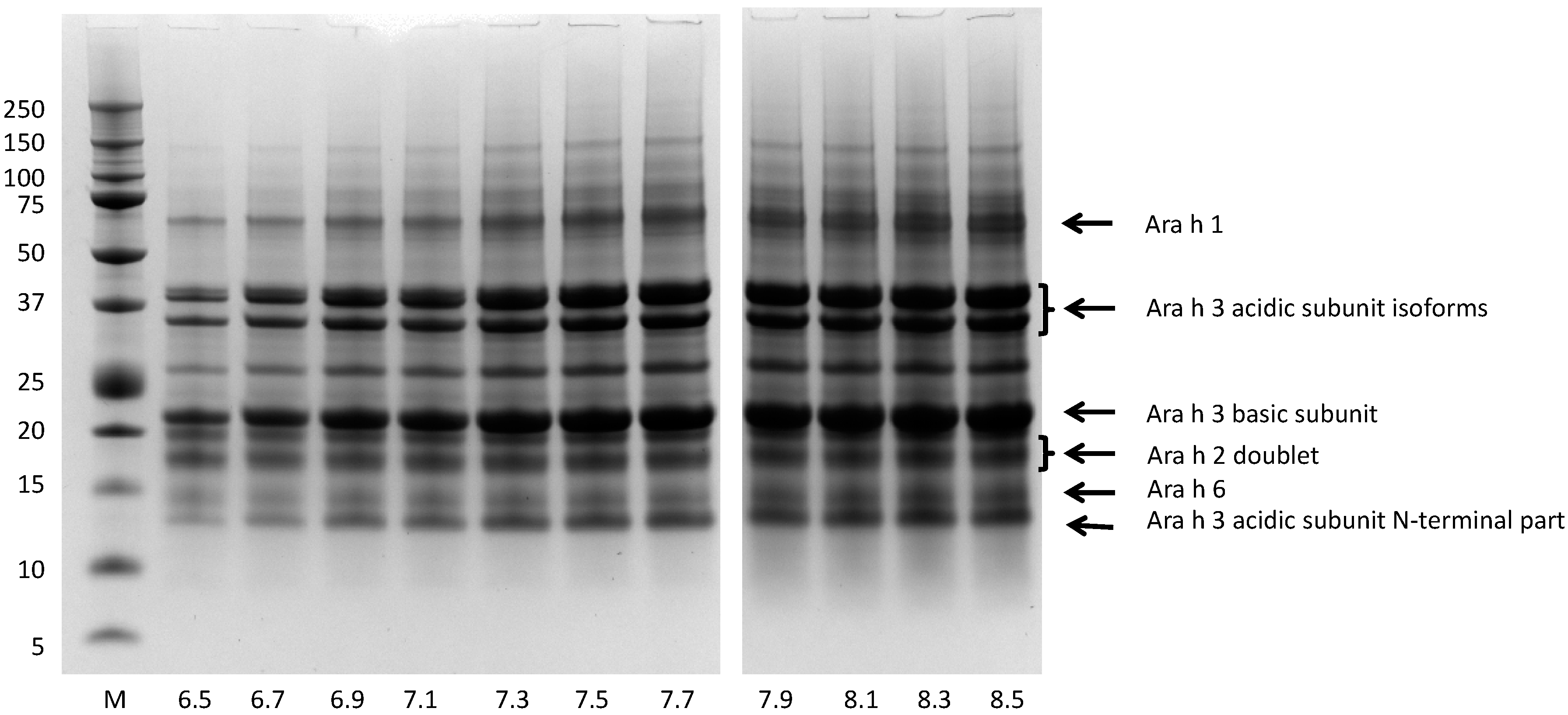
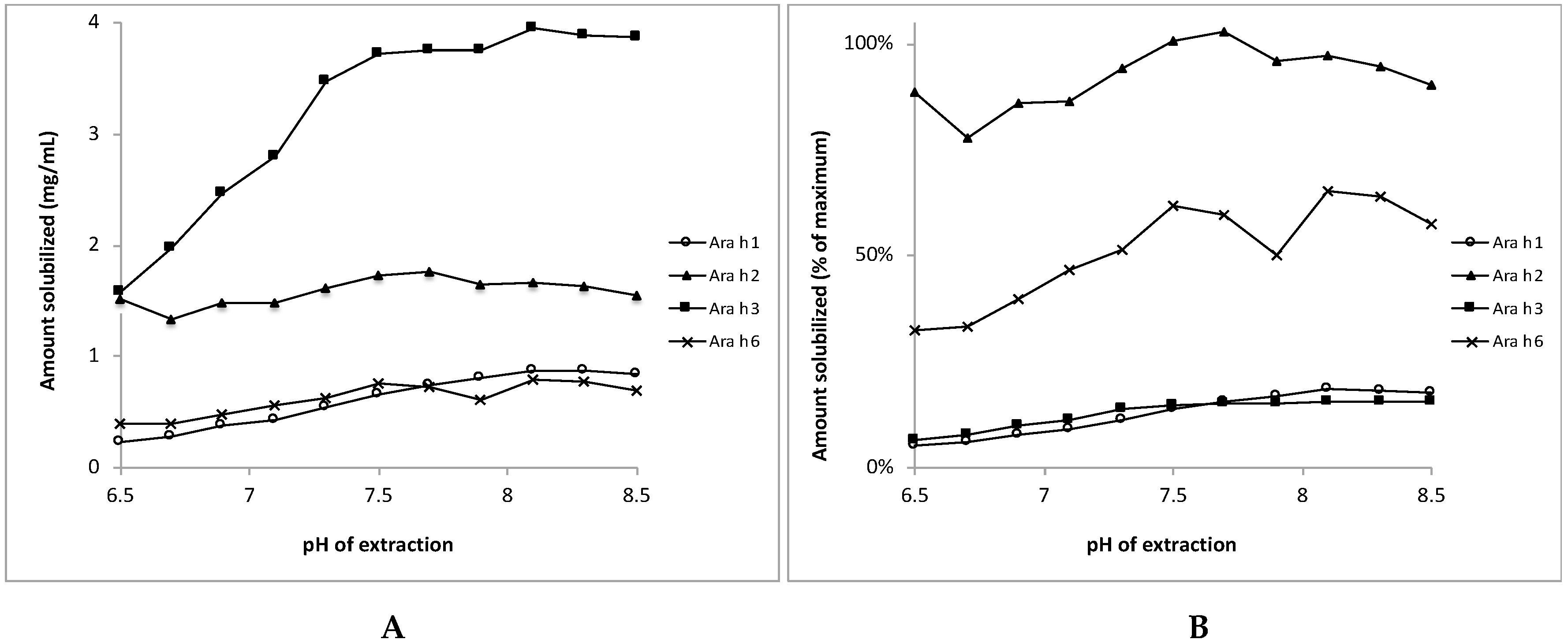
3.4. Bio-Accessibility of Peanut Allergens at the Normal pH Range of Saliva
3.5. Implications for Peanut Allergy Research
4. Conclusions
Author Contributions
Funding
Acknowledgment
Conflicts of Interest
References
- Archer, P. Overview of the Peanut Industry Supply Chain. In Peanuts: Genetics, Processing, and Utilization; Stalker, H.T., Wilson, R.F., Eds.; Academic Press and AOCS Press: London, UK, 2016. [Google Scholar]
- United States Department of Agriculture-Agricultural Marketing Servic. Available online: https://www.ams.usda.gov/grades-standards (accessed on 25 July 2017).
- Nwaru, B.I.; Hickstein, L.; Panesar, S.S.; Roberts, G.; Muraro, A.; Sheikh, A. Prevalence of common food allergies in Europe: A systematic review and meta-analysis. Allergy 2014, 69, 992–1007. [Google Scholar] [CrossRef] [PubMed]
- Sampson, H.A.; Aceves, S.; Bock, S.A.; James, J.; Jones, S.; Lang, D.; Nadeau, K.; Nowak-Wegrzyn, A.; Oppenheimer, J.; Perry, T.T.; et al. Food allergy: A practice parameter update—2014. J. Allergy Clin. Immunol. 2014, 134, 1016–1025. [Google Scholar] [CrossRef] [PubMed]
- Hourihane, J.O. Peanut allergy. Pediatr. Clin. N. Am. 2011, 58, 445–458. [Google Scholar] [CrossRef] [PubMed]
- Brough, H.A.; Makinson, K.; Penagos, M.; Maleki, S.J.; Cheng, H.; Douiri, A.; Stephens, A.C.; Turcanu, V.; Lack, G. Distribution of peanut protein in the home environment. J. Allergy Clin. Immunol. 2013, 132, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Baumert, J.L.; Bush, R.K.; Levy, M.B.; Koppelman, S.J.; Nordlee, J.A.; Hefle, S.L.; Taylor, S.L. Distribution of intact peanut protein and digestion-resistant Ara h2 peptide in human serum and saliva. J. Allergy Clin. Immunol. 2009, 123, S268. [Google Scholar] [CrossRef]
- Maloney, J.M.; Chapman, M.D.; Sicherer, S.H. Peanut allergen exposure through saliva: Assessment and interventions to reduce exposure. J. Allergy Clin. Immunol. 2006, 118, 719–724. [Google Scholar] [CrossRef] [PubMed]
- Nolan, R.C.; de Leon, M.P.; Rolland, J.M.; Loh, R.K.; O’Hehir, R.E. What’s in a kiss: Peanut allergen transmission as a sensitizer? J. Allergy Clin. Immunol. 2007, 119, 755. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, G.; Roberts, G.; Sayre, P.H.; Bahnson, H.T.; Radulovic, S.; Santos, A.F.; Brough, H.A.; Phippard, D.; Basting, M.; Feeney, M.; et al. Randomized trial of peanut consumption in infants at risk for peanut allergy. N. Engl. J. Med. 2015, 372, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Sicherer, S.H. Timing of food introduction and atopy prevention. Clin. Dermatol. 2017, 35, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, G.; Sayre, P.H.; Roberts, G.; Sever, M.L.; Lawson, K.; Bahnson, H.T.; Brough, H.A.; Santos, A.F.; Harris, K.M.; Radulovic, S.; et al. Effect of Avoidance on Peanut Allergy after Early Peanut Consumption. N. Engl. J. Med. 2016, 374, 1435–1443. [Google Scholar] [CrossRef] [PubMed]
- Bellach, J.; Schwarz, V.; Ahrens, B.; Trendelenburg, V.; Aksünger, Ö.; Kalb, B.; Niggemann, B.; Keil, T.; Beyer, K. Randomized placebo-controlled trial of hen’s egg consumption for primary prevention in infants. J. Allergy Clin. Immunol. 2017, 139, 1591–1599. [Google Scholar] [CrossRef] [PubMed]
- United States Department of Agriculture National Nutrient Database. Available online: https://ndb.nal.usda.gov/ndb/foods (accessed on 25 July 2017).
- Piersma, S.R.; Gaspari, M.; Hefle, S.L.; Koppelman, S.J. Proteolytic processing of the peanut allergen Ara h3. Mol. Nutr. Food Res. 2005, 49, 744–755. [Google Scholar] [CrossRef] [PubMed]
- Koppelman, S.J.; Jayasena, S.; Luykx, D.; Schepens, E.; Apostolovic, D.; de Jong, G.A.; Isleib, T.G.; Nordlee, J.; Baumert, J.; Taylor, S.L.; et al. Allergenicity attributes of different peanut market types. Food Chem. Toxicol. 2016, 91, 82–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oerise, N.L.; Lau, H.A.; Ritchey, S.J.; Murphy, E.W. Yield, proximate composition and mineral element content of three cultivars of raw and roasted peanuts. J. Food Sci. 1974, 39, 264–266. [Google Scholar] [CrossRef]
- Crea, F.; De Stefano, C.; Milea, D.; Pettignano, A.; Sammartano, S. SALMO and S3M: A Saliva Model and a Single Saliva Salt Model for Equilibrium Studies. Bioinorg. Chem. Appl. 2015, 2015, 267985:1–267985:12. [Google Scholar] [CrossRef] [PubMed]
- Clymer, J. Phosphate buffer Calculator. Available online: http://clymer.altervista.org/buffers/phos2.html (accessed on 25 July 2017).
- Chel-Guerrero, L.; Pérez-Flores, V.; Betancur-Ancona, D.; Dávila-Ortiz, G. Functional properties of flours and protein isolates from Phaseolus lunatus and Canavalia ensiformis seeds. J. Agric. Food Chem. 2002, 50, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Rangel, A.; Domont, G.B.; Pedrosa, C.; Ferreira, S.T. Functional properties of purified vicilins from cowpea (Vigna unguiculata) and pea (Pisum sativum) and cowpea protein isolate. J. Agric. Food Chem. 2003, 51, 5792–5797. [Google Scholar] [CrossRef] [PubMed]
- Petenate, A.M.; Glatz, C.E. Isoelectric precipitation of soy protein: I. Factors affecting particle size distribution. Biotechnol. Bioeng. 1983, 25, 3049–3058. [Google Scholar] [CrossRef] [PubMed]
- Petenate, A.M.; Glatz, C.E. Isoelectric precipitation of soy protein. II. Kinetics of protein aggregate growth and breakage. Biotechnol. Bioeng. 1983, 25, 3059–3078. [Google Scholar] [CrossRef] [PubMed]
- Kinsella, J.E. Functional properties of soy proteins. J. Am. Oil Chem. Soc. 1979, 56, 242–258. [Google Scholar] [CrossRef]
- Food and Agriculture Organization (FAO) Website on Isolated Soybean Protein. Available online: http://www.fao.org (accessed on 25 July 2017).
- Yu, J.; Goktepe, I.; Ahmedna, M. Peanut protein concentrate: Production and functional properties as affected by processing. Food Chem. 2007, 103, 121–129. [Google Scholar] [CrossRef]
- Fu, T.J.; Maks, N. Impact of thermal processing on ELISA detection of peanut allergens. J. Agric. Food Chem. 2013, 61, 5649–5658. [Google Scholar] [CrossRef] [PubMed]
- Poms, R.E.; Capelletti, C.; Anklam, E. Effect of roasting history and buffer composition on peanut protein extraction efficiency. Mol. Nutr. Food Res. 2004, 48, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Jayasena, S.; Smits, M.; Fiechter, D.; de Jong, A.; Nordlee, J.; Baumert, J.; Taylor, S.L.; Pieters, R.H.; Koppelman, S.J. Comparison of six commercial ELISA kits for their specificity and sensitivity in detecting different major peanut allergens. J. Agric. Food Chem. 2015, 63, 1849–1855. [Google Scholar] [CrossRef] [PubMed]
- Sathe, S.K.; Venkatachalam, M.; Sharma, G.M.; Kshirsagar, H.H.; Teuber, S.S.; Roux, K.H. Solubilization and electrophoretic characterization of select edible nut seed proteins. J. Agric. Food Chem. 2009, 57, 7846–7856. [Google Scholar] [CrossRef] [PubMed]
- Walczyk, N.E.; Smith, P.M.; Tovey, E.R.; Roberts, T.H. Peanut protein extraction conditions strongly influence yield of allergens Ara h1and 2 and sensitivity of immunoassays. Food Chem. 2017, 221, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Proctor, G.B. The physiology of salivary secretion. Periodontology 2000, 70, 11–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, P.D.V.; Grégio, A.M.; Machado, M.A.; de Lima, A.A.; Azevedo, L.R. Saliva composition and functions: A comprehensive review. J. Contemp. Dent. Pract. 2008, 9, 72–80. [Google Scholar]
- Ballmer-Weber, B.K.; Lidholm, J.; Fernández-Rivas, M.; Seneviratne, S.; Hanschmann, K.M.; Vogel, L.; Bures, P.; Fritsche, P.; Summers, C.; Knulst, A.C. IgE recognition patterns in peanut allergy are age dependent: Perspectives of the EuroPrevall study. Allergy 2015, 70, 391–407. [Google Scholar] [CrossRef] [PubMed]
- Berglund, J.P.; Szczepanski, N.; Penumarti, A.; Beavers, A.; Kesselring, J.; Orgel, K.; Burnett, B.; Burks, A.W.; Kulis, M. Preparation and Analysis of Peanut Flour Used in Oral Immunotherapy Clinical Trials. J. Allergy Clin. Immunol. Pract. 2017, 5, 1098–1104. [Google Scholar] [CrossRef] [PubMed]
- Koppelman, S.J.; de Jong, G.A.; Laaper-Ertmann, M.; Peeters, K.A.B.M.; Knulst, A.C.; Hefle, S.L.; Knol, E.F. Purification and immunoglobulin E-binding properties of peanut allergen Ara h6: Evidence for cross-reactivity with Ara h2. Clin. Exp. Allergy 2005, 35, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Breiteneder, H.; Mills, E.N. Molecular properties of food allergens. J. Allergy Clin. Immunol. 2005, 115, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Maleki, S.J.; Kopper, R.A.; Shin, D.S.; Park, C.W.; Compadre, C.M.; Sampson, H.; Burks, A.W.; Bannon, G.A. Structure of the major peanut allergen Ara h1may protect IgE-binding epitopes from degradation. J. Immunol. 2000, 164, 5844–5849. [Google Scholar] [CrossRef] [PubMed]
- Koppelman, S.J.; Knol, E.F.; Vlooswijk, R.A.; Wensing, M.; Knulst, A.C.; Hefle, S.L.; Gruppen, H.; Piersma, S. Peanut allergen Ara h3: Isolation from peanuts and biochemical characterization. Allergy 2003, 58, 1144–1151. [Google Scholar] [CrossRef] [PubMed]
- Van Boxtel, E.L.; van Beers, M.M.; Koppelman, S.J.; van den Broek, L.A.; Gruppen, H. Allergen Ara h1occurs in peanuts as a large oligomer rather than as a trimer. J. Agric. Food Chem. 2006, 54, 7180–7186. [Google Scholar] [CrossRef] [PubMed]
- Vissers, Y.M.; Iwan, M.; Adel-Patient, K.; Stahl-Skov, P.; Rigby, N.M.; Johnson, P.E.; Mandrup Müller, P.; Przybylski-Nicaise, L.; Schaap, M.; Ruinemans-Koerts, J.; et al. Effect of roasting on the allergenicity of major peanut allergens Ara h1and Ara h2/6: The necessity of degranulation assays. Clin. Exp. Allergy 2011, 41, 1631–1642. [Google Scholar] [CrossRef] [PubMed]
- Kulis, M.; Chen, X.; Lew, J.; Wang, Q.; Patel, O.P.; Zhuang, Y.; Murray, K.S.; Duncan, M.W.; Porterfield, H.S.; Burks, A.W.; et al. The 2S albumin allergens of Arachis hypogaea, Ara h2 and Ara h6, are the major elicitors of anaphylaxis and can effectively desensitize peanut-allergic mice. Clin. Exp. Allergy 2012, 42, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Peeters, K.A.; Koppelman, S.J.; van Hoffen, E.; van der Tas, C.W.; den Hartog-Jager, C.F.; Penninks, A.H.; Hefle, S.L.; Bruijnzeel-Koomen, C.A.; Knol, E.F.; Knulst, A.C. Does skin prick test reactivity to purified allergens correlate with clinical severity of peanut allergy? Clin. Exp. Allergy 2007, 37, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Porterfield, H.S.; Murray, K.S.; Schlichting, D.G.; Chen, X.; Hansen, K.C.; Duncan, M.W.; Dreskin, S.C. Effector activity of peanut allergens: A critical role for Ara h2, Ara h6, and their variants. Clin. Exp. Allergy 2009, 39, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Klemans, R.J.; Liu, X.; Knulst, A.C.; Knol, M.J.; Gmelig-Meyling, F.; Borst, E.; Pasmans, S.G.; Knol, E.F. IgE binding to peanut components by four different techniques: Ara h2 is the most relevant in peanut allergic children and adults. Clin. Exp. Allergy 2013, 43, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Klemans, R.J.; Knol, E.F.; Bruijnzeel-Koomen, C.A.; Knulst, A.C. The diagnostic accuracy of specific IgE to Ara h6 in adults is as good as Ara h2. Allergy 2014, 69, 1112–1114. [Google Scholar] [CrossRef] [PubMed]
- Sen, M.; Kopper, R.; Pons, L.; Abraham, E.C.; Burks, A.W.; Bannon, G.A. Protein structure plays a critical role in peanut allergen stability and may determine immunodominant IgE-binding epitopes. J. Immunol. 2002, 169, 882–887. [Google Scholar] [CrossRef] [PubMed]
- Koppelman, S.J.; Hefle, S.; Taylor, S.L.; de Jong, G.A. Digestion of peanut allergens Ara h1, Ara h2, Ara h3, and Ara h6: A comparative in vitro study and partial characterization of digestion-resistant peptides. Mol. Nutr. Food Res. 2010, 54, 1711–1721. [Google Scholar] [CrossRef] [PubMed]
- Apostolovic, D.; Stanic-Vucinic, D.; de Jongh, H.H.; de Jong, G.A.; Mihailovic, J.; Radosavljevic, J.; Radibratovic, M.; Nordlee, J.A.; Baumert, J.L.; Milcic, M.; et al. Conformational stability of digestion-resistant peptides of peanut conglutins reveals the molecular basis of their allergenicity. Sci. Rep. 2016, 6, 29249. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Theoretical pH | Measured pH of Buffer | pH after Peanut Flour Addition * | OH− Equivalent Added (mM) | pH Immediately after Adjusting | pH 2 min after Adjusting |
|---|---|---|---|---|---|
| 6.5 | 6.49 | 6.42 | 2.5 | 6.53 | 6.50 |
| 6.7 | 6.69 | 6.57 | 3.0 | 6.69 | 6.69 |
| 6.9 | 6.88 | 6.76 | 3.0 | 6.88 | 6.87 |
| 7.1 | 7.08 | 6.95 | 4.0 | 7.09 | 7.08 |
| 7.3 | 7.28 | 7.10 | 4.8 | 7.28 | 7.28 |
| 7.5 | 7.48 | 7.23 | 4.8 | 7.52 | 7.53 |
| 7.7 | 7.67 | 7.33 | 5.5 | 7.70 | 7.69 |
| 7.9 | 7.87 | 7.48 | 5.5 | 7.90 | 7.87 |
| 8.1 | 8.09 | 7.55 | 6.0 | 8.11 | 8.10 |
| 8.3 | 8.29 | 7.65 | 6.3 | 8.30 | 8.29 |
| 8.5 | 8.49 | 7.69 | 6.5 | 8.49 | 8.48 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koppelman, S.J.; Smits, M.; Tomassen, M.; De Jong, G.A.H.; Baumert, J.; Taylor, S.L.; Witkamp, R.; Veldman, R.J.; Pieters, R.; Wichers, H. Release of Major Peanut Allergens from Their Matrix under Various pH and Simulated Saliva Conditions—Ara h2 and Ara h6 Are Readily Bio-Accessible. Nutrients 2018, 10, 1281. https://doi.org/10.3390/nu10091281
Koppelman SJ, Smits M, Tomassen M, De Jong GAH, Baumert J, Taylor SL, Witkamp R, Veldman RJ, Pieters R, Wichers H. Release of Major Peanut Allergens from Their Matrix under Various pH and Simulated Saliva Conditions—Ara h2 and Ara h6 Are Readily Bio-Accessible. Nutrients. 2018; 10(9):1281. https://doi.org/10.3390/nu10091281
Chicago/Turabian StyleKoppelman, Stef J., Mieke Smits, Monic Tomassen, Govardus A.H. De Jong, Joe Baumert, Steve L. Taylor, Renger Witkamp, Robert Jan Veldman, Raymond Pieters, and Harry Wichers. 2018. "Release of Major Peanut Allergens from Their Matrix under Various pH and Simulated Saliva Conditions—Ara h2 and Ara h6 Are Readily Bio-Accessible" Nutrients 10, no. 9: 1281. https://doi.org/10.3390/nu10091281
APA StyleKoppelman, S. J., Smits, M., Tomassen, M., De Jong, G. A. H., Baumert, J., Taylor, S. L., Witkamp, R., Veldman, R. J., Pieters, R., & Wichers, H. (2018). Release of Major Peanut Allergens from Their Matrix under Various pH and Simulated Saliva Conditions—Ara h2 and Ara h6 Are Readily Bio-Accessible. Nutrients, 10(9), 1281. https://doi.org/10.3390/nu10091281