Elemental Metabolomics and Pregnancy Outcomes


Abstract
:1. Introduction
2. Maternal Nutrition
3. Micronutrition during Gestation and Lactation
3.1. Potassium
3.2. Calcium
3.3. Magnesium
3.4. Manganese
3.5. Iron
3.6. Copper
3.7. Zinc
3.8. Iodine
3.9. Selenium
4. Micronutrition in Gestational Complications
4.1. Gestational Diabetes Mellitus
4.2. Pre-eclampsia
4.3. Intrauterine Growth Restriction
4.4. Preterm Birth
5. Profiling Micronutrients in Disease
Elemental Metabolomics
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mertz, W. The essential trace elements. Science 1981, 213, 1332–1338. [Google Scholar] [CrossRef]
- Australian Bureau of Statistics: A.B.O. National Health Survey: First Results, 2014-15. Cat. no. 4364.0. 55.001. Available online: http://www.abs.gov.au/AUSSTATS/[email protected]/DetailsPage/4364.0.55.0012014-15?OpenDocument (accessed on 18 October 2015).
- Abu-Saad, K.; Fraser, D. Maternal nutrition and birth outcomes. Epidemiol. Rev. 2010, 32, 5–25. [Google Scholar] [CrossRef] [PubMed]
- Blumfield, M.L.; Hure, A.J.; Macdonald-Wicks, L.; Smith, R.; Collins, C.E. A systematic review and meta-analysis of micronutrient intakes during pregnancy in developed countries. Nutr. Rev. 2013, 71, 118–132. [Google Scholar] [CrossRef] [PubMed]
- Darnton-Hill, I.; Mkparu, U.C. Micronutrients in pregnancy in low- and middle-income countries. Nutrients 2015, 7, 1744–1768. [Google Scholar] [CrossRef] [PubMed]
- WHO/FAO. Vitamin and Mineral Requirements in Human Nutrition, 2nd ed.; World Health Organization and Food and Agriculture Organization of the United Nations: Geneva, Switzerland, 2004. [Google Scholar]
- Ahmed, T.; Hossain, M.; Sanin, K.I. Global burden of maternal and child undernutrition and micronutrient deficiencies. Ann. Nutr. Metab. 2012, 61, 8–17. [Google Scholar] [CrossRef] [PubMed]
- New Zealand Ministry of Health. Nutrient Reference Values for Australia and New Zealand. Available online: https://www.health.govt.nz/publication/nutrient-reference-values-australia-and-new-zealand (accessed on 21 October 2015).
- Vanderlelie, J.; Scott, R.; Shibl, R.; Lewkowicz, J.; Perkins, A.; Scuffham, P.A. First trimester multivitamin/mineral use is associated with reduced risk of pre-eclampsia among overweight and obese women. Matern. Child Nutr. 2016, 12, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Gernand, A.D.; Schulze, K.J.; Stewart, C.P.; West, K.P., Jr.; Christian, P. Micronutrient deficiencies in pregnancy worldwide: Health effects and prevention. Nat. Rev. Endocrinol. 2016, 12, 274–289. [Google Scholar] [CrossRef] [PubMed]
- Cheong, J.N.; Wlodek, M.E.; Moritz, K.M.; Cuffe, J.S. Programming of maternal and offspring disease: Impact of growth restriction, fetal sex and transmission across generations. J. Physiol. 2016, 594, 4727–4740. [Google Scholar] [CrossRef] [Green Version]
- Paglia, G.; Miedico, O.; Cristofano, A.; Vitale, M.; Angiolillo, A.; Chiaravalle, A.E.; Corso, G.; Di Costanzo, A. Distinctive pattern of serum elements during the progression of Alzheimer’s disease. Sci. Rep. 2016, 6, 22769. [Google Scholar] [CrossRef]
- Zhao, H.-W.; Lin, J.; Wang, X.-B.; Cheng, X.; Wang, J.-Y.; Hu, B.-L.; Zhang, Y.; Zhang, X.; Zhu, J.-H. Assessing plasma levels of selenium, copper, iron and zinc in patients of Parkinson’s disease. PLoS ONE 2013, 8, e83060. [Google Scholar] [CrossRef]
- Liu, G.; Sun, L.; Pan, A.; Zhu, M.; Li, Z.; Wang, Z.; Liu, X.; Ye, X.; Li, H.; Zheng, H. Nickel exposure is associated with the prevalence of type 2 diabetes in Chinese adults. Int. J. Epidemiol. 2014, 44, 240–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golasik, M.; Jawień, W.; Przybyłowicz, A.; Szyfter, W.; Herman, M.; Golusiński, W.; Florek, E.; Piekoszewski, W. Classification models based on the level of metals in hair and nails of laryngeal cancer patients: Diagnosis support or rather speculation? Metallomics 2015, 7, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Gluckman, P.; Hanson, M.; Seng, C.Y.; Bardsley, A. Nutrition and Lifestyle for Pregnancy and Breastfeeding; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Fall, C.H.; Fisher, D.J.; Osmond, C.; Margetts, B.M. Multiple micronutrient supplementation during pregnancy in low-income countries: A meta-analysis of effects on birth size and length of gestation. Food Nutr. Bull. 2009, 30, S533–S546. [Google Scholar] [CrossRef] [PubMed]
- Gaccioli, F.; Lager, S. Placental nutrient transport and intrauterine growth restriction. Front. Physiol. 2016, 7, 40. [Google Scholar] [CrossRef] [PubMed]
- Antony, A.C. Folate receptors. Annu.Rev. Nutr. 1996, 16, 501–521. [Google Scholar] [CrossRef] [PubMed]
- Solanky, N.; Jimenez, A.R.; D’Souza, S.; Sibley, C.; Glazier, J. Expression of folate transporters in human placenta and implications for homocysteine metabolism. Placenta 2010, 31, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J.; Osmond, C. Infant mortality, childhood nutrition, and ischaemic heart disease in England and Wales. Lancet 1986, 327, 1077–1081. [Google Scholar] [CrossRef]
- Thornburg, K.L.; Shannon, J.; Thuillier, P.; Turker, M.S. In utero life and epigenetic predisposition for disease. Adv. Genet. 2010, 71, 57. [Google Scholar]
- Roseboom, T.J.; Van Der Meulen, J.H.; Ravelli, A.C.; Osmond, C.; Barker, D.J.; Bleker, O.P. Effects of prenatal exposure to the Dutch famine on adult disease in later life: An overview. Mol. Cell. Endocrinol. 2001, 185, 93–98. [Google Scholar] [CrossRef]
- Grieger, J.A.; Clifton, V.L. A review of the impact of dietary intakes in human pregnancy on infant birthweight. Nutrients 2014, 7, 153–178. [Google Scholar] [CrossRef]
- Lewicka, I.; Kocyłowski, R.; Grzesiak, M.; Gaj, Z.; Oszukowski, P.; Suliburska, J. Selected trace elements concentrations in pregnancy and their possible role—Literature review. Ginekol. Pol. 2017, 88, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Burtis, C.A.; Ashwood, E.R.; Bruns, D.E. Tietz Textbook of Clinical Chemistry and Molecular Diagnostics; Elsevier Saunders: Philadelphia, PA, USA, 2006. [Google Scholar]
- Matsunami, K.; Imai, A.; Tamaya, T. Hypokalemia in a pregnant woman with long-term heavy cola consumption. Int. J. Gynecol. Obstet. 1994, 44, 283–284. [Google Scholar] [CrossRef]
- Appel, C.C.; Myles, T.D. Caffeine-induced hypokalemic paralysis in pregnancy. Obstet. Gynecol. 2001, 97, 805–807. [Google Scholar] [PubMed]
- Khaw, K.; Barrett-Connor, E. The association between blood pressure, age, and dietary sodium and potassium: A population study. Circulation 1988, 77, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Hofstee, P.; McKeating, D.R.; Perkins, A.V.; Cuffe, J.S. Placental adaptations to micronutrient dysregulation in the programming of chronic disease. Clin. Exp. Pharmacol. Physiol. 2018, 45, 871–884. [Google Scholar] [CrossRef] [PubMed]
- Wolak, T.; Shoham-Vardi, I.; Sergienko, R.; Sheiner, E. High potassium level during pregnancy is associated with future cardiovascular morbidity. J. Matern. Fetal Neonatal Med. 2016, 29, 1021–1024. [Google Scholar] [CrossRef]
- Farinde, A. Lab Values, Normal Adult: Laboratory Reference Ranges in Healthy Adults. Available online: https://emedicine.medscape.com/article/2172316-overview (accessed on 16 April 2018).
- FSANZ. The 23rd Australian Total Diet Study; Food Standards Australia New Zealand: Barton, ACT, Australia, 2011.
- Jantzen, C.; Jørgensen, H.L.; Duus, B.R.; Sporring, S.L.; Lauritzen, J.B. Chromium and cobalt ion concentrations in blood and serum following various types of metal-on-metal hip arthroplasties: A literature overview. Acta Orthop. 2013, 84, 229–236. [Google Scholar] [CrossRef]
- Galinier, A.; Périquet, B.; Lambert, W.; Garcia, J.; Assouline, C.; Rolland, M.; Thouvenot, J.-P. Reference range for micronutrients and nutritional marker proteins in cord blood of neonates appropriated for gestational ages. Early Human Dev. 2005, 81, 583–593. [Google Scholar] [CrossRef]
- Mariath, A.B.; Bergamaschi, D.P.; Rondó, P.H.; Ana, C.A.T.; de Fragas Hinnig, P.; Abbade, J.F.; Diniz, S.G. The possible role of selenium status in adverse pregnancy outcomes. Br. J. Nutr. 2011, 105, 1418–1428. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.M. Vitamin D and the Calcium-Sensing Receptor. Vitamin D, 3rd ed.; Elsevier: New York, NY, USA, 2011; pp. 425–456. [Google Scholar]
- Brini, M.; Ottolini, D.; Calì, T.; Carafoli, E. Calcium in health and disease. In Interrelations between Essential Metal Ions and Human Diseases; Sigel, A., Sigel, H., Sigel, R., Eds.; Springer: Berlin, Germany, 2013; pp. 81–137. [Google Scholar]
- Jorde, R.; Sundsfjord, J.; Haug, E.; Bønaa, K.H. Relation between low calcium intake, parathyroid hormone, and blood pressure. Hypertension 2000, 35, 1154–1159. [Google Scholar] [CrossRef]
- Lumbers, E.R.; Pringle, K.G. Roles of the circulating renin-angiotensin-aldosterone system in human pregnancy. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 306, R91–R101. [Google Scholar] [CrossRef] [PubMed]
- Imdad, A.; Bhutta, Z.A. Effects of calcium supplementation during pregnancy on maternal, fetal and birth outcomes. Paediatr. Perinat. Epidemiol. 2012, 26, 138–152. [Google Scholar] [CrossRef] [PubMed]
- Fatemi, S.; Ryzen, E.; Flores, J.; Endres, D.B.; Rude, R.K. Effect of experimental human magnesium depletion on parathyroid hormone secretion and 1,25-dihydroxyvitamin D metabolism. J. Clin. Endocrinol. Metab. 1991, 73, 1067–1072. [Google Scholar] [CrossRef] [PubMed]
- Arnaud, M.J. Update on the assessment of magnesium status. Br. J. Nutr. 2008, 99, S24–S36. [Google Scholar] [CrossRef] [PubMed]
- Rude, R.K. Magnesium deficiency: A cause of heterogenous disease in humans. J. Bone Miner. Res. 1998, 13, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Spencer, B.; Vanderlelie, J.; Perkins, A. Essentiality of trace element micronutrition in human pregnancy: A systematic review. J. Pregnancy Child Health 2015, 2, 2. [Google Scholar] [CrossRef]
- Williams, M.; Todd, G.; Roney, N.; Crawford, J.; Coles, C.; McClure, P.; Garey, J.; Zaccaria, K.; Citra, M. Toxicological Profile for Manganese; Agency for Toxic Substances and Disease Registry: Atlanta, GA, USA, 2012.
- Fitsanakis, V.A.; Zhang, N.; Garcia, S.; Aschner, M. Manganese (Mn) and iron (Fe): Interdependency of transport and regulation. Neurotox. Res. 2010, 18, 124–131. [Google Scholar] [CrossRef]
- Greger, J. Dietary standards for manganese: Overlap between nutritional and toxicological studies. J. Nutr. 1998, 128, 368S–371S. [Google Scholar] [CrossRef]
- Freeland-Graves, J.H.; Lin, P.-H. Plasma uptake of manganese as affected by oral loads of manganese, calcium, milk, phosphorus, copper, and zinc. J. Am. Coll. Nutr. 1991, 10, 38–43. [Google Scholar] [CrossRef]
- Law, N.A.; Caudle, M.T.; Pecoraro, V.L. Manganese redox enzymes and model systems: Properties, structures, and reactivity. Adv. Inorg. Chem. 1998, 46, 305–440. [Google Scholar]
- Davis, C.D.; Greger, J. Longitudinal changes of manganese-dependent superoxide dismutase and other indexes of manganese and iron status in women. Am. J. Clin. Nutr. 1992, 55, 747–752. [Google Scholar] [CrossRef] [Green Version]
- Friedman, B.; Freeland-Graves, J.H.; Bales, C.W.; Behmardi, F.; Shorey-Kutschke, R.L.; Willis, R.A.; Crosby, J.B.; Trickett, P.C.; Houston, S.D. Manganese balance and clinical observations in young men fed a manganese-deficient diet. J. Nutr. 1987, 117, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Tholin, K.; Sandström, B.; Palm, R.; Hallmans, G. Changes in blood manganese levels during pregnancy in iron supplemented and non supplemented women. J. Trace Elem. Med. Biol. 1995, 9, 13–17. [Google Scholar] [CrossRef]
- Spencer, A. Whole blood manganese levels in pregnancy and the neonate. Nutrition 1999, 15, 731–734. [Google Scholar] [CrossRef]
- Henn, B.C.; Bellinger, D.C.; Hopkins, M.R.; Coull, B.A.; Ettinger, A.S.; Jim, R.; Hatley, E.; Christiani, D.C.; Wright, R.O. Maternal and cord blood manganese concentrations and early childhood neurodevelopment among residents near a mining-impacted superfund site. Environ. Health Perspect. 2017, 125. [Google Scholar] [CrossRef]
- Institute of Medicine Dietary. Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc; Institute of Medicine National Academy Press: Washington, WA, USA, 2001. [Google Scholar]
- Hulten, L.; Gramatkovski, E.; Gleerup, A.; Hallberg, L. Iron absorption from the whole diet. Relation to meal composition, iron requirements and iron stores. Eur. J. Clin. Nutr. 1995, 49, 794–808. [Google Scholar] [PubMed]
- Hider, R.C.; Kong, X. Iron: Effect of overload and deficiency. Met. Ions Life Sci. 2013, 13, 229–294. [Google Scholar] [PubMed]
- Henderson, B.; Kühn, L. Interaction between iron-regulatory proteins and their RNA target sequences, iron-responsive elements. Prog. Mol. Subcell. Biol. 1997, 18, 17–139. [Google Scholar]
- Koulaouzidis, A.; Said, E.; Cottier, R.; Saeed, A.A. Soluble transferrin receptors and iron deficiency, a step beyond ferritin. A systematic review. J. Gastrointest. Liver Dis. 2009, 18, 345–352. [Google Scholar]
- Dandona, P.; Hussain, M.; Varghese, Z.; Politis, D.; Flynn, D.; Hoffbrand, A. Insulin resistance and iron overload. Ann. Clin. Biochem. 1983, 20, 77–79. [Google Scholar] [CrossRef] [PubMed]
- Peña, M.M.; Lee, J.; Thiele, D.J. A delicate balance: Homeostatic control of copper uptake and distribution. J. Nutr. 1999, 129, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- McKeown, N.M. Whole grain intake and insulin sensitivity: Evidence from observational studies. Nutr. Rev. 2004, 62, 286. [Google Scholar] [PubMed]
- Trumbo, P.; Schlicker, S.; Yates, A.A.; Poos, M. Dietary reference intakes for energy, carbohydrate, fiber, fat, fatty acids, cholesterol, protein and amino acids. J. Am. Diet. Assoc. 2002, 102, 1621–1630. [Google Scholar] [CrossRef]
- Drehmer, M.; Camey, S.A.; Nunes, M.A.; Duncan, B.B.; Lacerda, M.; Pinheiro, A.P.; Schmidt, M.I. Fibre intake and evolution of BMI: From pre-pregnancy to postpartum. Public Health Nutr. 2013, 16, 1403–1413. [Google Scholar] [CrossRef] [PubMed]
- Klevay, L.M. Is the Western diet adequate in copper? J. Trace Elem. Med. Biol. 2011, 25, 204–212. [Google Scholar] [CrossRef]
- Al-Rashid, R.A.; Spangler, J. Neonatal copper deficiency. N. Engl. J. Med. 1971, 285, 841–843. [Google Scholar] [CrossRef]
- Prasad, A.S. Impact of the discovery of human zinc deficiency on health. J. Trace Elem. Med. Biol. 2014, 28, 357–363. [Google Scholar] [CrossRef]
- Andreini, C.; Banci, L.; Bertini, I.; Rosato, A. Counting the zinc-proteins encoded in the human genome. J. Proteome Res. 2006, 5, 196–201. [Google Scholar] [CrossRef]
- Foster, M.; Herulah, U.N.; Prasad, A.; Petocz, P.; Samman, S. Zinc status of vegetarians during pregnancy: A systematic review of observational studies and meta-analysis of zinc intake. Nutrients 2015, 7, 4512–4525. [Google Scholar] [CrossRef]
- O’Dell, B.L. Role of zinc in plasma membrane function. J. Nutr. 2000, 130, 1432S–1436S. [Google Scholar] [CrossRef]
- Mossad, S.B.; Macknin, M.L.; Mendendorp, S.V.; Mason, P. Zinc gluconate lozenges for treating the common cold: A randomized, double-blind, placebo-controlled study. Ann. Intern. Med. 1996, 125, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.; De Benoist, B.; Darnton-Hill, I.; Delange, F.M.; Organization, W.H.; UNICEF. Iodine Deficiency in Europe: A Continuing Public Health Problem; WHO: Geneva, Switzerland, 2007. [Google Scholar]
- Hetzel, B.S. Iodine and neuropsychological development. J. Nutr. 2000, 130, 493S–495S. [Google Scholar] [CrossRef] [PubMed]
- Vejbjerg, P.; Knudsen, N.; Perrild, H.; Laurberg, P.; Andersen, S.; Rasmussen, L.B.; Ovesen, L.; Jørgensen, T. Estimation of iodine intake from various urinary iodine measurements in population studies. Thyroid 2009, 19, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Richard, K.; Holland, O.; Landers, K.; Vanderlelie, J.J.; Hofstee, P.; Cuffe, J.S.; Perkins, A.V. Review: Effects of maternal micronutrient supplementation on placental function. Placenta 2017, 54, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Vermiglio, F.; Lo Presti, V.P.; Moleti, M.; Sidoti, M.; Tortorella, G.; Scaffidi, G.; Castagna, M.G.; Mattina, F.; Violi, M.A.; Crisa, A.; et al. Attention deficit and hyperactivity disorders in the offspring of mothers exposed to mild-moderate iodine deficiency: A possible novel iodine deficiency disorder in developed countries. J. Clin. Endocrinol. Metab. 2004, 89, 6054–6060. [Google Scholar] [CrossRef] [PubMed]
- Sang, Z.; Wei, W.; Zhao, N.; Zhang, G.; Chen, W.; Liu, H.; Shen, J.; Liu, J.; Yan, Y.; Zhang, W. Thyroid dysfunction during late gestation is associated with excessive iodine intake in pregnant women. J. Clin. Endocrinol. Metab. 2012, 97, E1363–E1369. [Google Scholar] [CrossRef]
- Connelly, K.J.; Boston, B.A.; Pearce, E.N.; Sesser, D.; Snyder, D.; Braverman, L.E.; Pino, S.; LaFranchi, S.H. Congenital hypothyroidism caused by excess prenatal maternal iodine ingestion. J. Pediatr. 2012, 161, 760–762. [Google Scholar] [CrossRef] [PubMed]
- Besser, J.M.; Canfield, T.J.; La Point, T.W. Bioaccumulation of organic and inorganic selenium in a laboratory food chain. Environ. Toxicol. Chem. 1993, 12, 57–72. [Google Scholar] [CrossRef]
- Institute of Medicine, Food and Nutrition Board. Dietary Reference Intakes: Vitamin C, Vitamin E, Selenium, and Carotenoids; National Academy Press: Washington, WA, USA, 2000. [Google Scholar]
- Liu, X.; Zhao, Z.; Duan, B.; Hu, C.; Zhao, X.; Guo, Z. Effect of applied sulphur on the uptake by wheat of selenium applied as selenite. Plant Soil 2015, 386, 35–45. [Google Scholar] [CrossRef]
- Navarro-Alarcon, M.; Cabrera-Vique, C. Selenium in food and the human body: A review. Sci. Total Environ. 2008, 400, 115–141. [Google Scholar] [CrossRef]
- Dumont, E.; De Pauw, L.; Vanhaecke, F.; Cornelis, R. Speciation of se in Bertholletia excelsa (Brazil nut): A hard nut to crack? Food Chem. 2006, 95, 684–692. [Google Scholar] [CrossRef]
- Dumont, E.; Vanhaecke, F.; Cornelis, R. Selenium speciation from food source to metabolites: A critical review. Anal. Bioanal. Chem. 2006, 385, 1304–1323. [Google Scholar] [CrossRef] [PubMed]
- Korotkov, K.V.; Novoselov, S.V.; Hatfield, D.L.; Gladyshev, V.N. Mammalian selenoprotein in which selenocysteine (Sec) incorporation is supported by a new form of Sec insertion sequence element. Mol. Cell. Biol. 2002, 22, 1402–1411. [Google Scholar] [CrossRef] [PubMed]
- Papp, L.V.; Lu, J.; Holmgren, A.; Khanna, K.K. From selenium to selenoproteins: Synthesis, identity, and their role in human health. Antioxid. Redox Signal. 2007, 9, 775–806. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.B.; Köhrle, J. The impact of iron and selenium deficiencies on iodine and thyroid metabolism: Biochemistry and relevance to public health. Thyroid 2002, 12, 867–878. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, J.M.; Contreras, R.; Chen, W.; Sacks, D.A. Trends in the prevalence of preexisting diabetes and gestational diabetes mellitus among a racially/ethnically diverse population of pregnant women, 1999–2005. Diabetes Care 2008, 31, 899–904. [Google Scholar] [CrossRef]
- Dabelea, D.; Snell-Bergeon, J.K.; Hartsfield, C.L.; Bischoff, K.J.; Hamman, R.F.; McDuffie, R.S. Increasing prevalence of gestational diabetes mellitus (GDM) over time and by birth cohort: Kaiser permanente of Colorado GDM screening program. Diabetes Care 2005, 28, 579–584. [Google Scholar] [CrossRef]
- Jameson, J.L.; De Groot, L.J. Endocrinology: Adult and Pediatric, 7th ed.; Elsevier Saunders: Philadelphia, PA, USA, 2015; pp. 788–804. [Google Scholar]
- Chatterjee, R.; Yeh, H.-C.; Edelman, D.; Brancati, F. Potassium and risk of type 2 diabetes. Expert Rev. Endocrinol. Metab. 2011, 6, 665–672. [Google Scholar] [CrossRef]
- Vambergue, A.; Fajardy, I. Consequences of gestational and pregestational diabetes on placental function and birth weight. World J. Diabetes 2011, 2, 196. [Google Scholar] [CrossRef]
- Basaki, M.; Saeb, M.; Nazifi, S.; Shamsaei, H. Zinc, copper, iron, and chromium concentrations in young patients with type 2 diabetes mellitus. Biol. Trace Elem. Res. 2012, 148, 161–164. [Google Scholar] [CrossRef]
- El-Yazigi, A.; Hannan, N.; Raines, D.A. Effect of diabetic state and related disorders on the urinary excretion of magnesium and zinc in patients. Diabetes Research Edinb. Scotl. 1993, 22, 67–75. [Google Scholar]
- Jansen, J.; Rosenkranz, E.; Overbeck, S.; Warmuth, S.; Mocchegiani, E.; Giacconi, R.; Weiskirchen, R.; Karges, W.; Rink, L. Disturbed zinc homeostasis in diabetic patients by in vitro and in vivo analysis of insulinomimetic activity of zinc. J. Nutr. Biochem. 2012, 23, 1458–1466. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, R.S. The role of zinc in growth and cell proliferation. J. Nutr. 2000, 130, 1500S–1508S. [Google Scholar] [CrossRef]
- Zhang, C.; Rawal, S. Dietary iron intake, iron status, and gestational diabetes. Am. J. Clin. Nutr. 2017, 106, 1672S–1680S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, J.M. Preeclampsia: What we know and what we do not know. Semin. Perinatol. 2000, 24, 24–28. [Google Scholar] [CrossRef]
- McDonald, S.D.; Han, Z.; Walsh, M.W.; Gerstein, H.C.; Devereaux, P.J. Kidney disease after preeclampsia: A systematic review and meta-analysis. Am. J. Kidney Dis. 2010, 55, 1026–1039. [Google Scholar] [CrossRef]
- Ghulmiyyah, L.; Sibai, B. Maternal mortality from preeclampsia/eclampsia. Semin. Perinatol. 2012, 36, 56–59. [Google Scholar] [CrossRef]
- Morris, C.D.; Jacobson, S.-L.; Anand, R.; Ewell, M.G.; Hauth, J.C.; Curet, L.B.; Catalano, P.M.; Sibai, B.M.; Levine, R.J. Nutrient intake and hypertensive disorders of pregnancy: Evidence from a large prospective cohort. Am. J. Obstet. Gynecol. 2001, 184, 643–651. [Google Scholar] [CrossRef]
- Hofmeyr, G.J.; Lawrie, T.A.; Atallah, A.N.; Duley, L. Calcium supplementation during pregnancy for preventing hypertensive disorders and related problems. Cochrane Database Syst. Rev. 2010. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.; Sharma, P.; Kulshreshtha, S.; Mohan, G.; Singh, S. The role of calcium, magnesium, and zinc in pre-eclampsia. Biol. Trace Elem. Res. 2010, 133, 162–170. [Google Scholar] [CrossRef]
- Mistry, H.D.; Pipkin, F.B.; Redman, C.W.; Poston, L. Selenium in reproductive health. Am. J. Obstet. Gynecol. 2012, 206, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ergaz, Z.; Avgil, M.; Ornoy, A. Intrauterine growth restriction—Etiology and consequences: What do we know about the human situation and experimental animal models? Reprod. Toxicol. 2005, 20, 301–322. [Google Scholar] [CrossRef] [PubMed]
- Herrera, E.A.; Alegría, R.; Farias, M.; Díaz-López, F.; Hernández, C.; Uauy, R.; Regnault, T.R.; Casanello, P.; Krause, B.J. Assessment of in vivo fetal growth and placental vascular function in a novel intrauterine growth restriction model of progressive uterine artery occlusion in guinea pigs. J. Physiol. 2016, 594, 1553–1561. [Google Scholar] [CrossRef] [PubMed]
- Nardozza, L.M.M.; Araujo Júnior, E.; Barbosa, M.M.; Caetano, A.C.R.; Lee, D.J.R.; Moron, A.F. Fetal growth restriction: Current knowledge to the general Obs/Gyn. Arch. Gynecol. Obstet. 2012, 286, 1–13. [Google Scholar] [CrossRef]
- Figueras, F.; Gratacos, E. Stage-based approach to the management of fetal growth restriction. Prenat. Diagn. 2014, 34, 655–659. [Google Scholar] [CrossRef]
- Henriksen, T.; Clausen, T. The fetal origins hypothesis: Placental insufficiency and inheritance versus maternal malnutrition in well-nourished populations. Acta Obstet. Gynecol. Scand. 2002, 81, 112–114. [Google Scholar] [CrossRef] [PubMed]
- Coan, P.M.; Vaughan, O.R.; Sekita, Y.; Finn, S.L.; Burton, G.J.; Constancia, M.; Fowden, A.L. Adaptations in placental phenotype support fetal growth during undernutrition of pregnant mice. J. Physiol. 2010, 588, 527–538. [Google Scholar] [CrossRef] [Green Version]
- Sandovici, I.; Hoelle, K.; Angiolini, E.; Constância, M. Placental adaptations to the maternal–fetal environment: Implications for fetal growth and developmental programming. Reprod. Biomed. Online 2012, 25, 68–89. [Google Scholar] [CrossRef]
- Moore, K.L.; Persaud, T.V.N.; Torchia, M. The developing human: Clinically Oriented Embryology, 10th ed.; Elsevier: Philadelphia, PA, USA, 2015. [Google Scholar]
- Wood, R.J. Manganese and birth outcome. Nutr. Rev. 2009, 67, 416–420. [Google Scholar] [CrossRef]
- Wang, H.; Hu, Y.-F.; Hao, J.-H.; Chen, Y.-H.; Su, P.-Y.; Wang, Y.; Yu, Z.; Fu, L.; Xu, Y.-Y.; Zhang, C. Maternal zinc deficiency during pregnancy elevates the risks of fetal growth restriction: A population-based birth cohort study. Sci. Rep. 2015, 5, 11262. [Google Scholar] [CrossRef]
- Cao, X.-Y.; Jiang, X.-M.; Dou, Z.-H.; Rakeman, M.A.; Zhang, M.-L.; O’Donnell, K.; Ma, T.; Amette, K.; DeLong, N.; DeLong, G.R. Timing of vulnerability of the brain to iodine deficiency in endemic cretinism. N. Engl. J. Med. 1994, 331, 1739–1744. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, R.L.; Culhane, J.F.; Iams, J.D.; Romero, R. Epidemiology and causes of preterm birth. Lancet 2008, 371, 75–84. [Google Scholar] [CrossRef]
- Schieve, L.A.; Tian, L.H.; Rankin, K.; Kogan, M.D.; Yeargin-Allsopp, M.; Visser, S.; Rosenberg, D. Population impact of preterm birth and low birth weight on developmental disabilities in US children. Ann. Epidemiol. 2016, 26, 267–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crump, C.; Sundquist, K.; Sundquist, J. Adult outcomes of preterm birth. Prev. Med. 2016, 91, 400–401. [Google Scholar] [CrossRef]
- Villar, J.; Abdel-Aleem, H.; Merialdi, M.; Mathai, M.; Ali, M.M.; Zavaleta, N.; Purwar, M.; Hofmeyr, J.; Campódonico, L.; Landoulsi, S. World Health Organization randomized trial of calcium supplementation among low calcium intake pregnant women. Am. J. Obstet. Gynecol. 2006, 194, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, E.; Ryan, K.J.; Monson, R.R.; Schoenbaum, S.C. Association of maternal hematocrit with premature labor. Am. J. Obstet. Gynecol. 1988, 159, 107–114. [Google Scholar] [CrossRef]
- Scholl, T.O.; Hediger, M.L.; Fischer, R.L.; Shearer, J.W. Anemia vs iron deficiency: Increased risk of preterm delivery in a prospective study. Am. J. Clin. Nutr. 1992, 55, 985–988. [Google Scholar] [CrossRef] [PubMed]
- Dobrzynski, W.; Szymanski, W.; Zachara, B.A.; Trafikowska, U.; Trafikowska, A.; Pilecki, A. Decreased selenium concentration in maternal and cord blood in preterm compared with term delivery. Analyst 1998, 123, 93–97. [Google Scholar] [CrossRef]
- Iranpour, R.; Zandian, A.; Mohammadizadeh, M.; Mohammadzadeh, A.; Balali-Mood, M.; Hajiheydari, M. Comparison of maternal and umbilical cord blood selenium levels in term and preterm infants. Chin. J. Contemp. Pediatr. 2009, 11, 513–516. [Google Scholar]
- Baxter, I. Ionomics: The functional genomics of elements. Brief. Funct. Genomics 2010, 9, 149–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Georgiou, C.A.; Brusic, V. Elemental metabolomics. Brief. Bioinforma. 2017, 19, 524–536. [Google Scholar] [CrossRef]
- Zhang, Y. Trace Elements and Healthcare: A Bioinformatics Perspective. In Translational Informatics in Smart Healthcare; Shen, B., Ed.; Springer: Berlin, Germany, 2017; pp. 63–98. [Google Scholar]
- Yu, D.; Danku, J.M.; Baxter, I.; Kim, S.; Vatamaniuk, O.K.; Vitek, O.; Ouzzani, M.; Salt, D.E. High-resolution genome-wide scan of genes, gene-networks and cellular systems impacting the yeast ionome. BMC Genomics 2012, 13, 623. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.-Y.; Salt, D.E. Plant ionomics: From elemental profiling to environmental adaptation. Mol. Plant 2016, 9, 787–797. [Google Scholar] [CrossRef]
- Ma, S.; Lee, S.-G.; Kim, E.B.; Park, T.J.; Seluanov, A.; Gorbunova, V.; Buffenstein, R.; Seravalli, J.; Gladyshev, V.N. Organization of the mammalian ionome according to organ origin, lineage specialization, and longevity. Cell Rep. 2015, 13, 1319–1326. [Google Scholar] [CrossRef]
- Malinouski, M.; Hasan, N.M.; Zhang, Y.; Seravalli, J.; Lin, J.; Avanesov, A.; Lutsenko, S.; Gladyshev, V.N. Genome-wide RNAi ionomics screen reveals new genes and regulation of human trace element metabolism. Nat. Commun. 2014, 5, 3301. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Yu, Y.; Huang, T.; An, P.; Yu, D.; Yu, Z.; Li, H.; Sheng, H.; Cai, L.; Xue, J. Associations between ionomic profile and metabolic abnormalities in human population. PLoS ONE 2012, 7, e38845. [Google Scholar] [CrossRef] [PubMed]
- Herman, M.; Golasik, M.; Piekoszewski, W.; Walas, S.; Napierala, M.; Wyganowska-Swiatkowska, M.; Kurhanska-Flisykowska, A.; Wozniak, A.; Florek, E. Essential and Toxic Metals in Oral Fluid–a Potential Role in the Diagnosis of Periodontal Diseases. Biol. Trace Elem. Res. 2016, 173, 275–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]


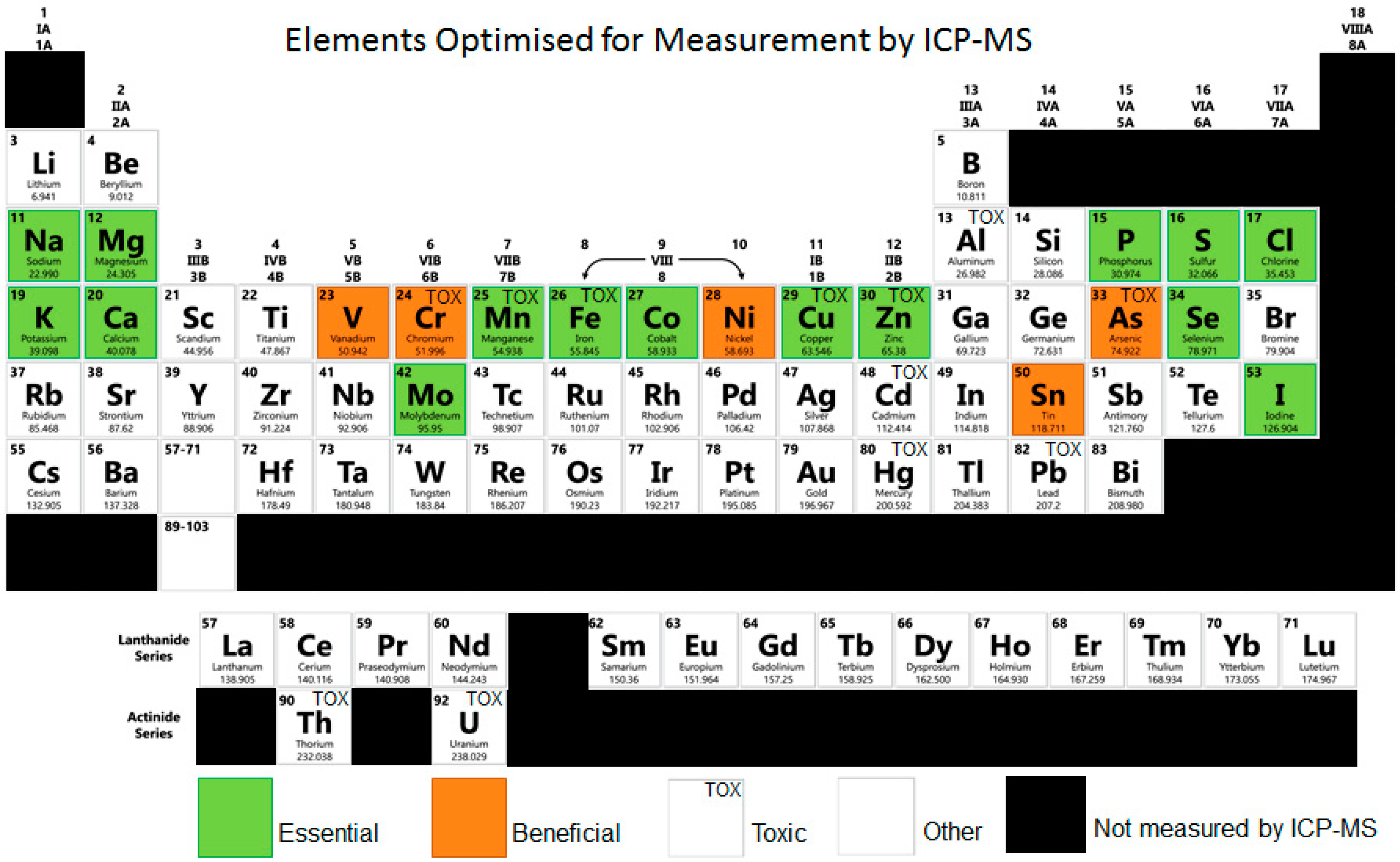{kind=link}
| RDI | Blood Reference Range | Serum/Plasma Reference Range | Urine Reference Range | Cord Reference Range | |
|---|---|---|---|---|---|
| Na (Sodium) | 460–920 mg [6] | 310–335 mg/dL [36] | - | 45 mg/dL [36] | 290–380 mg/dL [36] |
| Mg (Magnesium) | 350 mg [6] | 3.6–6.1 mg/dL [36] | 1.7–2.2 mg/dL [16] | 145–245 mg/day [36] | - |
| P (Phosphorus) | 1000 mg [6] | 6–9.2 mg/dL [36] | 2.7–4.4 mg/dL [36] | 0.4–1.3 g/day [36] | - |
| S (Sulfur) | 900 mg [6] | - | - | - | - |
| K (Potassium) | 2800 mg [6] | 14.1–20.3 mg/dL [36] | 13.3–17.2 mg/dL [36] | 97.7–490 mg/dL [36] | 21.9–46.92 mg/dL [36] |
| Ca (Calcium) | 1000 mg [6] | 8.6–10.2 mg/dL [36] | 8.6–10 mg/dL [36] | 100–300mg/day [36] | - |
| V (Vanadium) | <1.8 mg [32] | - | <1 µg/L [36] | - | - |
| Cr (Chromium) | 30 µg [6] | 0.5–2.5 µg/L [33] | 0.8–5.1 µg/mL [33] | - | - |
| Mn (Manganese) | 5 mg [6] | 4–15 µg/L [36] | 0.4–0.85 µg/L [36] | 1–8 µg/L [36] | - |
| Fe (Iron) | 27 mg [6] | - | 50–170 µg/dL [36] | - | - |
| Co (Cobalt) | - | 0.7–3.4 µg/L [33] | 0.3–7.5 µg/L [33] | - | - |
| Ni (Nickel) | - | - | <2 µg/L [36] | - | - |
| Cu (Copper) | 1.3 mg [6] | 70–140 µg/dL [36] | 80–155 µg/dL [36] | 3–35 µg/day [36] | 4.6–8.8 µmol/L [34] |
| Zn (Zinc) | 11 mg [6] | 4.5–6.5 mg/L [36] | 0.66–1.10 µg/mL [36] | - | 15.8–22 µmol/L [34] |
| As (arsenic) | - | 0.2–2.3 µg/dL [36] | - | 5–50 µg/day [36] | - |
| Se (Selenium) | 65 µg [6] | - | 70–150 µg/L [36] | 15–50 µg/L [36] | 0.5–0.7 µmol/L [34] |
| Mo (Molybdenum) | 50 µg [16] | 0.6–4 µg/L [36] | 0.3–2.0 µg/L [36] | - | - |
| I (Iodine) | 220 µg [6] | - | 40–92 µg/L [36] | 150–249 µg/L [36] | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McKeating, D.R.; Fisher, J.J.; Perkins, A.V. Elemental Metabolomics and Pregnancy Outcomes. Nutrients 2019, 11, 73. https://doi.org/10.3390/nu11010073
McKeating DR, Fisher JJ, Perkins AV. Elemental Metabolomics and Pregnancy Outcomes. Nutrients. 2019; 11(1):73. https://doi.org/10.3390/nu11010073
Chicago/Turabian StyleMcKeating, Daniel R., Joshua J. Fisher, and Anthony V. Perkins. 2019. "Elemental Metabolomics and Pregnancy Outcomes" Nutrients 11, no. 1: 73. https://doi.org/10.3390/nu11010073
APA StyleMcKeating, D. R., Fisher, J. J., & Perkins, A. V. (2019). Elemental Metabolomics and Pregnancy Outcomes. Nutrients, 11(1), 73. https://doi.org/10.3390/nu11010073