Meat Proteins as Dipeptidyl Peptidase IV Inhibitors and Glucose Uptake Stimulating Peptides for the Management of a Type 2 Diabetes Mellitus In Silico Study



Abstract
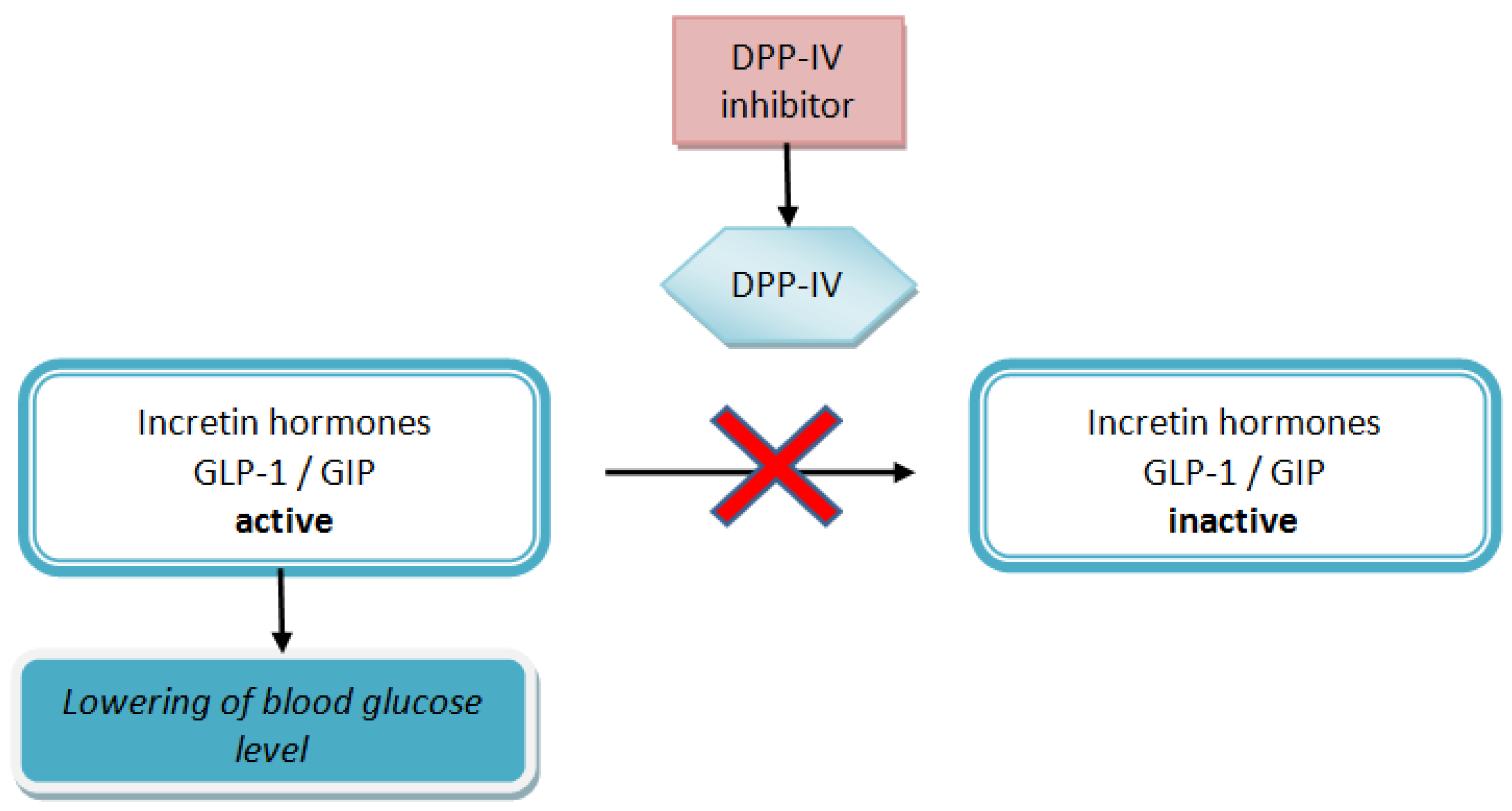
:1. Introduction
2. Materials and Methods
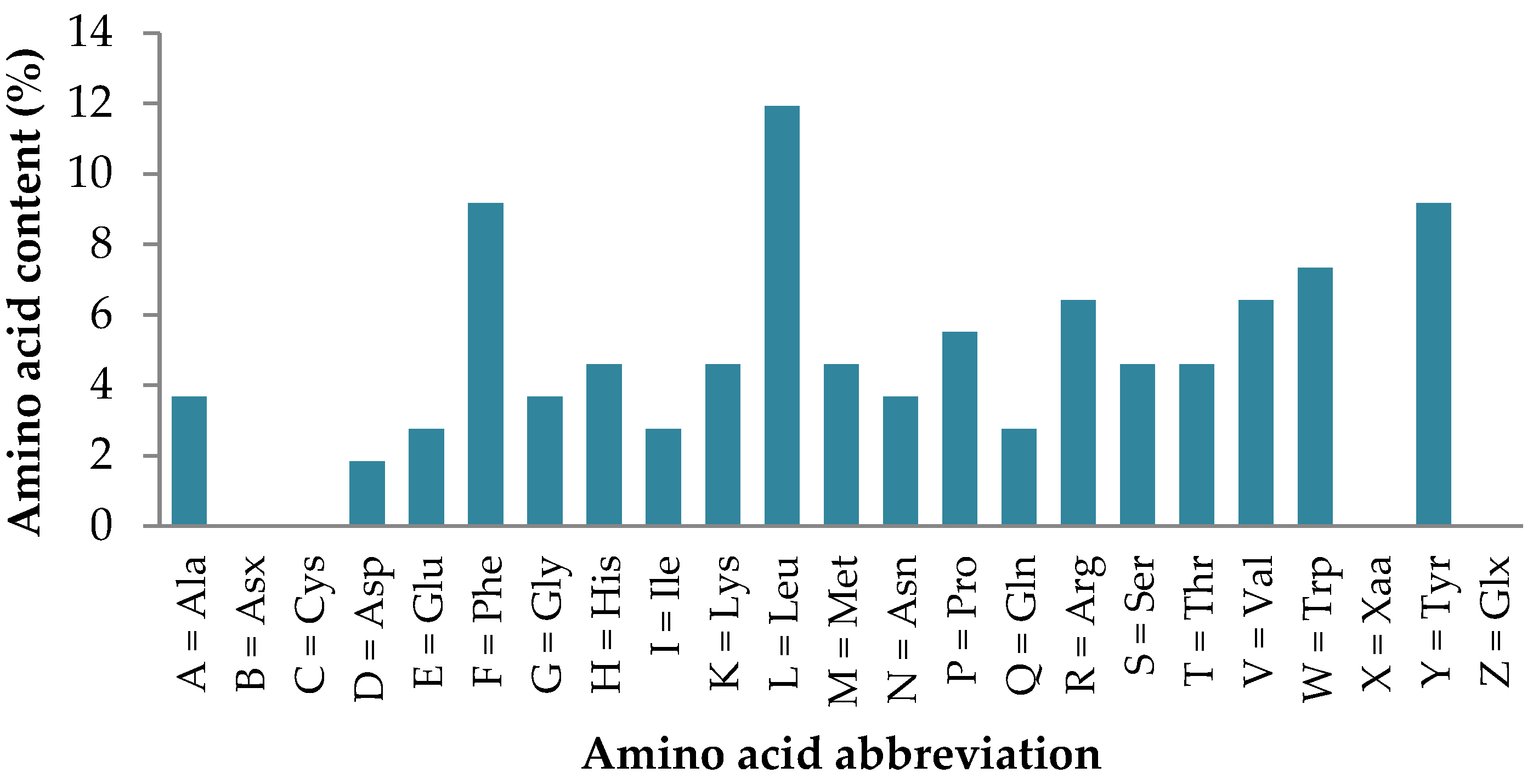
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization (WHO). Available online: http://www.who.int/diabetes/publications/grd-2016/en/ (accessed on 2 September 2018).
- Patil, P.; Mandal, S.; Tomar, S.K.; Anand, S. Food protein-derived bioactive peptides in management of type 2 diabetes. Eur. J. Nutr. 2015, 54, 863–880. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, I.M.; Li-Chan, E.C. Overview of food products and dietary constituents with antidiabetic properties and their putative mechanisms of action: A natural approach to complement pharmacotherapy in the management of diabetes. Mol. Nutr. Food Res. 2014, 58, 61–78. [Google Scholar] [CrossRef] [PubMed]
- Deacon, C.F.; Hughes, T.E.; Holst, J.J. Dipeptidyl peptidase IV inhibition potentiates the insulinotropic effect of glucagon-like peptide 1 in the anesthetized pig. Diabetes 1998, 47, 764–769. [Google Scholar] [CrossRef] [PubMed]
- Drucker, D.J. Enhancing the action of incretin hormones: A new whey forward. Endocrinology 2006, 147, 3171–3172. [Google Scholar] [CrossRef]
- Power, O.; Nongonierma, A.B.; Jakeman, P.; FitzGerald, R.J. Food protein hydrolysates as a source of dipeptidyl peptidase IV inhibitory peptides for the management of type 2 diabetes. Proc. Nutr. Soc. 2014, 73, 34–46. [Google Scholar] [CrossRef]
- Demuth, H.U.; McIntosh, C.H.; Pederson, R.A. Type 2 diabetes-therapy with dipeptidyl peptidase IV inhibitors. BBA Proteins Proteom. 2005, 1751, 33–44. [Google Scholar] [CrossRef]
- Jakubowicz, D.; Froy, O.; Ahren, B.; Boaz, M.; Landau, Z.; Bar-Dayan, Y.; Ganz, T.; Barnea, M.; Wainstein, J. Incretin, insulinotropic and glucose-lowering effects of whey protein pre-load in type 2 diabetes: A randomised clinical trial. Diabetologia 2014, 57, 1807–1811. [Google Scholar] [CrossRef]
- Lacroix, I.M.; Li-Chan, E.C. Evaluation of the potential of dietary proteins as precursors of dipeptidyl peptidase (DPP)-IV inhibitors by an in silico approach. J. Funct. Foods 2012, 4, 403–422. [Google Scholar] [CrossRef]
- Gallego, M.; Aristoy, M.C.; Toldra, F. Dipeptidyl peptidase IV inhibitory peptides generated in Spanish dry-cured ham. Meat Sci. 2014, 96, 757–761. [Google Scholar] [CrossRef] [Green Version]
- Stadnik, J.; Kęska, P. Meat and fermented meat products as a source of bioactive peptides. Acta Sci. Pol. Technol. Aliment. 2015, 14, 181–190. [Google Scholar] [CrossRef]
- Kęska, P.; Stadnik, J. Porcine myofibrillar proteins as potential precursors of bioactive peptides-an in silico study. Food Funct. 2016, 7, 2878–2885. [Google Scholar] [CrossRef] [PubMed]
- Kęska, P.; Stadnik, J. Stability of Antiradical Activity of Protein Extracts and Hydrolysates from Dry-Cured Pork Loins with Probiotic Strains of LAB. Nutrients 2018, 10, 521. [Google Scholar] [CrossRef] [PubMed]
- Keska, P.; Stadnik, J. Ageing-Time Dependent Changes of Angiotensin I-Converting Enzyme-Inhibiting Activity of Protein Hydrolysates Obtained from Dry-Cured Pork Loins Inoculated with Probiotic Lactic Acid Bacteria. Int. J. Pept. Res. Ther. 2018, 25, 1173–1185. [Google Scholar] [CrossRef] [Green Version]
- Kęska, P.; Stadnik, J. Taste-active peptides and amino acids of pork meat as components of dry-cured meat products: An in-silico study. J. Sens. Stud. 2017, 32, e12301. [Google Scholar] [CrossRef]
- Keska, P.; Stadnik, J. Antimicrobial peptides of meat origin-an in silico and in vitro analysis. Protein Pept. Lett. 2017, 24, 165–173. [Google Scholar] [CrossRef]
- Keska, P.; Wojciak, K.M.; Stadnik, J. Bioactive peptides from beef products fermented by acid whey—In vitro and in silico study. Sci. Agric. 2019, 76, 311–320. [Google Scholar] [CrossRef]
- Sayd, T.; Dufour, C.; Chambon, C.; Buffiere, C.; Remond, D.; Sante-Lhoutellier, V. Combined in vivo and in silico approaches for predicting the release of bioactive peptides from meat digestion. Food Chem. 2018, 249, 111–118. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Prospects for the management of type 2 diabetes using food protein-derived peptides with dipeptidyl peptidase IV (DPP-IV) inhibitory activity. Curr. Opin. Food Sci. 2016, 8, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Nongonierma, A.B.; FitzGerald, R.J. Structure activity relationship modelling of milk protein-derived peptides with dipeptidyl peptidase IV (DPP-IV) inhibitory activity. Peptides 2016, 79, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Scott, D.O. Structure-based drug metabolism predictions for drug design. Chem. Biol. Drug Des. 2010, 75, 3–17. [Google Scholar] [CrossRef]
- Bielenica, A.; Kossakowski, J. Zastosowanie metod obliczeniowych do wyznaczania budowy modeli farmakoforowych receptorów 5-HT1A, 5-HT2A oraz 5-HT7. Biul. Wydz. Farm. WUM 2010, 2, 13–21. [Google Scholar]
- Al-Sehemi, A.G.; Irfan, A.; Alrumman, S.A.; Hesham, A.E. Antibacterial activities, DFT and QSAR studies of quinazolinone compounds. Bull. Chem. Soc. Ethiop. 2016, 30, 307–316. [Google Scholar] [CrossRef]
- Kuruvilla, T.K.; Muthu, S.; Prasana, J.C.; George, J.; SaraSaji, R.; Geoffrey, B.; David, R.H.A. Molecular docking, spectroscopic studies on 4-[2-(Dipropylamino) ethyl]-1, 3-dihydro-2H-indol-2-one and QSAR study of a group of dopamine agonists by density functional method. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2019, 222, 117185. [Google Scholar] [CrossRef] [PubMed]
- Swiss-Prot. Available online: http://www.uniprot.org (accessed on 10 October 2018).
- BIOPEP. Available online: http://www.uwm.edu.pl/biochemia/index.php/pl/biopep (accessed on 10 October 2018).
- Dziuba, J.; Iwaniak, A.; Minkiewicz, P. Computer-aided characteristics of proteins as potential precursors of bioactive peptides. Polimery 2003, 48, 50–53. [Google Scholar] [CrossRef] [Green Version]
- Minkiewicz, P.; Dziuba, J.; Iwaniak, A.; Dziuba, M.; Darewicz, M. BIOPEP database and other programs for processing bioactive peptide sequences. J. AOAC Int. 2008, 91, 965–980. [Google Scholar]
- PepStat Tools. Available online: http://www.innovagen.com/proteomics-tools (accessed on 10 October 2018).
- PeptideRanker. Available online: http://bioware.ucd.ie/~compass/biowareweb (accessed on 10 October 2018).
- Becke, A.D. Becke’s three parameter hybrid method using the LYP correlation functional. J. Chem. Phys. 1993, 98, 5648–5652. [Google Scholar] [CrossRef]
- Krishnan, R.; Binkley, J.S.; Seeger, R.; Pople, J.A. Self-consistent molecular orbital methods. XX. A basis set for correlated wave functions. J. Chem. Phys. 1980, 72, 650–654. [Google Scholar] [CrossRef]
- Frisch, M.J.; Pople, J.A.; Binkley, J.S. Self-consistent molecular orbital methods 25. Supplementary functions for Gaussian basis sets. J. Chem. Phys. 1984, 80, 3265–3269. [Google Scholar] [CrossRef]
- PQS Version 4.0, Parallel Quantum Solutions; Green Acres Road: Fayetteville, AR, USA, 2013.
- Baker, J.; Wolinski, K.; Malagoli, M.; Kinghorn, D.; Wolinski, P.; Magyarfalvi, G.; Saebo, S.; Janowski, T.; Pulay, P. Quantum chemistry in parallel with PQS. J. Comput. Chem. 2009, 30, 317–335. [Google Scholar] [CrossRef]
- Keska, P.; Stadnik, J. Angiotensin I-converting enzyme inhibitory peptides generated from in vitro gastrointestinal digestion of pork proteins—In silico study. In Badania i Rozwój Młodych Naukowców w Polsce-Nauki Przyrodnicze Część I. Monografie; Panfil, M., Ed.; Młodzi Naukowcy: Poznań, Poland, 2016; pp. 58–64. ISBN 978-83-942083-6-3. [Google Scholar]
- Lan, V.T.T.; Ito, K.; Ito, S.; Kawarasaki, Y. Trp-Arg-Xaa tripeptides act as uncompetitive-type inhibitors of human dipeptidyl peptidase IV. Peptides 2014, 54, 166–170. [Google Scholar] [CrossRef]
- Lan, V.T.T.; Ito, K.; Ohno, M.; Motoyama, T.; Ito, S.; Kawarasaki, Y. Analyzing a dipeptide library to identify human dipeptidyl peptidase IV inhibitor. Food Chem. 2015, 175, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Iwai, K.; Hasegawa, T.; Taguchi, Y.; Morimatsu, F.; Sato, K.; Nakamura, Y.; Higashi, A.; Kido, Y.; Nakobo, Y.; Ohtsuki, K. Identification of food-derived collagen peptides in human blood after oral ingestion of gelatin hydrolysates. J. Agric. Food Chem. 2005, 53, 6531–6536. [Google Scholar] [CrossRef] [PubMed]
- Rahfeld, J.; Schierborn, M.; Hartrodt, B.; Neubert, K.; Heins, J. Are diprotin A (Ile-Pro-Ile) and diprotin B (Val-Pro-Leu) inhibitors or substrates of dipeptidyl peptidase IV? Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1991, 1076, 314–316. [Google Scholar] [CrossRef]
- Lacroix, I.M.; Li-Chan, E.C. Food-derived dipeptidyl-peptidase IV inhibitors as a potential approach for glycemic regulation–Current knowledge and future research considerations. Trends Food Sci. Technol. 2016, 54, 1–16. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; Mooney, C.; Shields, D.C.; FitzGerald, R.J. In silico approaches to predict the potential of milk protein-derived peptides as dipeptidyl peptidase IV (DPP-IV) inhibitors. Peptides 2014, 57, 43–51. [Google Scholar] [CrossRef]
- Tulipano, G.; Faggi, L.; Nardone, A.; Cocchi, D.; Caroli, A.M. Characterisation of the potential of β-lactoglobulin and α-lactalbumin as sources of bioactive peptides affecting incretin function: In silico and in vitro comparative studies. Int. Dairy J. 2015, 48, 66–72. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; Mooney, C.; Shields, D.C.; FitzGerald, R.J. Inhibition of dipeptidyl peptidase IV and xanthine oxidase by amino acids and dipeptides. Food Chem. 2013, 141, 644–653. [Google Scholar] [CrossRef] [Green Version]
- Tulipano, G.; Sibilia, V.; Caroli, A.M.; Cocchi, D. Whey proteins as source of dipeptidyl dipeptidase IV (dipeptidyl peptidase-4) inhibitors. Peptides 2011, 32, 835–838. [Google Scholar] [CrossRef]
- Schwartz, G.J. Central leucine sensing in the control of energy homeostasis. Endocrinol. Metab. Clin. N. Am. 2013, 42, 81–87. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, R.; Chen, X.; Zeng, Z.; Ma, H.; Chen, S. Dipeptidyl peptidase IV-inhibitory peptides derived from silver carp (Hypophthalmichthys molitrix Val.) proteins. J. Agric. Food Chem. 2016, 64, 831–839. [Google Scholar] [CrossRef]
- Morifuji, M.; Koga, J.; Kawanaka, K.; Higuchi, M. Branched-chain amino acid-containing dipeptides, identified from whey protein hydrolysates, stimulate glucose uptake rate in L6 myotubes and isolated skeletal muscles. J. Nutr. Sci. Vitaminol. 2009, 55, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Sakkiah, S.; Meganathan, C.; Sohn, Y.S.; Namadevan, S.; Lee, K.W. Identification of important chemical features of 11β-hydroxysteroid dehydrogenase type 1 inhibitors: Application of ligand based virtual screening and density functional theory. Int. J. Mol. Sci. 2012, 13, 5138–5162. [Google Scholar] [CrossRef] [PubMed]
- Al-Sehemi, A.G.; Irfan, A.; Aljubiri, S.M.; Shaker, K.H. Density functional theory investigations of radical scavenging activity of 3’-Methyl-quercetin. J. Saudi Chem. Soc. 2016, 20, S21–S28. [Google Scholar] [CrossRef]
- Auriekson, N.Q.; Bruno, A.Q.G.; Waldir, M.M.; Rosivaldo, S.B. A theoretical antioxidant pharmacophore for resveratrol. Eur. J. Med. Chem. 2009, 44, 1644–1649. [Google Scholar] [CrossRef]
- Montoro-Garcia, S.; Zafrilla-Rentero, M.P.; Celdran-de Haro, F.M.; Pinero-de Armas, J.J.; Toldra, F.; Tejada-Portero, L.; Abellan-Aleman, J. Effects of dry-cured ham rich in bioactive peptides on cardiovascular health: A randomized controlled trial. J. Funct. Foods 2017, 38, 160–167. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Protein Name | Abbreviation | Entry Name (ID) | Protein Name | Abbreviation | Entry Name (ID) |
|---|---|---|---|---|---|
| Myofibrillar Proteins | Sarcoplasmic Proteins | ||||
| Actin, alpha skeletal muscle | ACTS | P68137 | Myoglobin | MB | P02189 |
| Myosin-2 | MYH2 | Q9TV63 | Creatine kinase U-type, | CKMT1 | Q29577 |
| Tropomyosin alpha-3 chain | TPM3 | A1XQV4 | Creatine kinase M-type | CKM | Q5XLD3 |
| Troponin C, skeletal muscle | TNNC2 | P02587 | Glyceraldehyde-3-phosphat dehydrogenase | GAPDH | P00355 |
| Troponin T, fast skeletal muscle | TNNT3 | Q75NG9 | L-lactate dehydrogenase A-chain | LDHA | P00339 |
| Troponin T, slow skeletal muscle | TNNT1 | Q75ZZ6 | Phosphoglycerate kinase 1 | PGK1 | Q7SIB7 |
| Titin (fragment) | TTN | Q29117 | Alpha-1,4 glucan phosphorylase | PYGM | F1RQQ8 |
| Nebulin | NEB | Q3Y5G4 | Fructose-bisphosphate aldolase | ALDOA | Q6UV40 |
| Protein | DPP-IV Inhibitory | Stimulating a | ||
|---|---|---|---|---|
| Parameter A | Parameter B | Parameter A | Parameter B | |
| ACTS | 0.6499 | 0.000258 | 0.0451 | - |
| MYH2 | 0.5910 | 0.000255 | 0.0552 | - |
| TPM3 | 0.5035 | 0.000141 | 0.0810 | - |
| TNNC2 | 0.5031 | 0.000247 | 0.0181 | - |
| TNNT3 | 0.5941 | 0.000237 | 0.1218 | - |
| TNNT1 | 0.5196 | 0.000323 | 0.1489 | - |
| TTN | 0.6713 | 0.000307 | 0.0385 | - |
| NEB | 0.6475 | 0.000361 | 0.0296 | - |
| MB | 0.6494 | 0.000534 | 0.0714 | - |
| CKMT1 | 0.6394 | 0.000200 | 0.0457 | - |
| CKM | 0.6352 | 0.000434 | 0.0472 | - |
| GAPDH | 0.6697 | 0.000401 | 0.0300 | - |
| LDHA | 0.6175 | 0.000341 | 0.0542 | - |
| PGK1 | 0.6451 | 0.000395 | 0.0432 | - |
| PYGM | 0.6449 | 0.000413 | 0.0356 | - |
| ALDOA | 0.6147 | 0.000659 | 0.0642 | - |
| Protein | DPP-IV Inhibitory | Stimulating a | ||
|---|---|---|---|---|
| Parameter AE | Parameter W | Parameter AE | Parameter W | |
| ACTS | 0.0307 | 0.0496 | 0.0026 | 0.0598 |
| MYH2 | 0.0414 | 0.0737 | 0.0030 | 0.0567 |
| TPM3 | 0.0272 | 0.5670 | 0.0034 | 0.0455 |
| TNNC2 | 0.0182 | 0.0380 | - | - |
| TNNT3 | 0.0249 | 0.0440 | 0.0036 | 0.0307 |
| TNNT1 | 0.0257 | 0.0492 | 0.0074 | 0.0516 |
| TTN | 0.0270 | 0.0423 | 0.0017 | 0.0457 |
| NEB | 0.0371 | 0.0604 | 0.0017 | 0.0607 |
| MB | 0.0437 | 0.0706 | 0.0063 | 0.0916 |
| CKMT1 | 0.0465 | 0.0766 | 0.0070 | 0.1584 |
| CKM | 0.0456 | 0.0760 | 0.0051 | 0.1118 |
| GAPDH | 0.0232 | 0.0584 | - | - |
| LDHA | 0.0320 | 0.0548 | 0.0029 | 0.0587 |
| PGK1 | 0.0348 | 0.0568 | 0.0046 | 0.1100 |
| PYGM | 0.0505 | 0.0828 | 0.0023 | 0.0669 |
| ALDOA | 0.0177 | 0.0299 | 0.0088 | 0.0237 |
| Protein | Activity of Peptides | |
|---|---|---|
| DPP-IV Inhibitory | Stimulating c | |
| ACTS | EK(3) b [121-122][222-223][245-246]; AL [176-177]; SL [147-148]; GY [205-206]; IL [200-201]; IW [89-90]; MK [198-199]; SF [207-208]; TL [70-71]; VK [19-20] | IL [200-201] |
| MYH2 | EK(10) [280-281][450-451][915-916][1003-1004][1031-1032] [1262-1263][1264-1265][1290-1291][1465-1466][1581-1582]; AL (4) [413-414][1034-1035][1408-1409][1868-1869]; SL (2)[599-600][1528-1529]; GL (2) [799-800][1903-1904]; VR (2) [696-697] [1883-1884]; PL [870-871]; AF (2) [641-642][850-851]; GF (3) [353-354][734-735][824-825]; HW [856-857]; IL (3) [739-740][1504-1505][1614-1615]; IR [259-260]; MF [452-453]; MK (3) [860-861][1536-1537][1844-1845]; ML (2) [170-171][1479-1480]; MR [1216-1217]; NF [1500-1501]; NL (3) [107-108][1013-1014][1558-1559]; NY [1700-1701]; PF (2) [31-32][324-325]; PK (2) [562-563][587-588]; PW [858-859]; QF [168-169]; QL (4) [287-288][920-921][1336-1337][1366-1367]; QY [471-472]; SK (3) [1307-1308][1410-1411][1960-1961]; SY (2) [293-294][1943-1944]; TF [528-529]; TK (7) [891-892][945-946][1029-1030][1059-1060][1063-1064][1421-1422][1467-1468]; TL (3) [639-640] [996-997] [1295-1296]; TY (2) [119-120][1915-1916]; VF [791-792]; VK (7) [48-49][419-420][1011-1012][1257-1258][1911-1912][1941-1942][1990-1991]; VL (3) [514-515][715-716][752-753] | IL(4) [739-740] [1504-1505] [1614-1615] [1927-1928]; VL (3) [514-515] [715-716][752-753] |
| TPM3 | EK [258-259]; AL [138-139]; DR (2) [20-21] [102-103]; IL [233-234]; NR [91-92]; QL [38-39]; SK [36-37] | IL [233-234] |
| TNNC2 | DR [105-106]; ML [45-46]; SY [9-10] | - |
| TNNT3 | AL [163-164]; PL [204-205]; EY [235-236]; IR [126-127]; PK [54-55]; QL [228-229]; VL [197-198] | VL [197-198] |
| TNNT1 | EK [211-212]; PL [198-199]; IL [191-192]; MR [137-138]; PK [40-41]; VK [170-171]; VL [157-158] | VL [157-158]; IL [191-192] |
| TTN | SL(2) [412-413][445-446]; PL [500-501]; NF [129-130]; NY [290-291]; PK [486-487]; SW (3) [74-75][377-378][576-577]; SY [302-303]; TK (2) [379-380][493-494]; TL (2) [233-234][235-236]; VK [410-411]; VL [220-221] | VL [220-221] |
| NEB | VPL [485-487]; HL (3) [610-611][1112-1113][1520-1521]; EK (4) [532-533] [569-570][603-604][1067-1068]; AL (2) [1435-1436][1744-1745]; SL (2) [155-156][1078-1079]; GL (4)[659-660][910-911][1654-1655][1706-1707]; VR [78-79]; PL (2) [1140-1141][1746-1747]; AW [671-672]; AF [793-794]; AY (2) [657-658][908-909]; DR [1571-1572]; EY [848-849]; GF (3) [601-602] [784-785][852-853]; HF [1589-1590]; HR [1500-1501]; HW [102-103]; IR [1261-1262]; MK (6) [294-295][350-351][673-674][924-925][1069-1070][1587-1588]; MR [1175-1176]; NL (4) [75-76][687-688][1281-1282] [1677-1678]; NY (3) [469-470][537-538][702-703]; PF [1295-1296]; PY [52-53]; QY (2) [665-666][1350-1351]; SK (3) [107-108][605-606][1029-1030]; TF [1173-1174]; TL (4) [442-443][539-540][651-652][1503-1504]; TR (2) [1390-1391][1679-1680]; VK (2) [548-549][1318-1319]; VL (2) [366-367][575-576]; VY [1525-1526] | VL(2) [366-367] [575-576] |
| MB | HL [49-50]; EK [42-43]; AL [139-140]; GF [155-156]; IR [31-32]; QL [9-10]; VL [11-12] | VL [11-12] |
| CKMT1 | AL [19-20]; SL [178-179]; GL (2) [176-177][297-298]; VR [168-169]; PL [343-344]; MW[315-316]; DR [385-386]; GY [321-322]; IL (2) [323-324][354-355]; IR [153-154]; IW [268-269]; PK [352-353]; QY [46-47]; SF [265-266]; SK [346-347]; TL [87-88]; VF [290-291]; VL [163-164] | VL [163-164]; IL (2)[323-324] [354-355] |
| CKM | AL [33-34]; EK [380-381]; VR [135-136]; PL [179-180]; MW [280-281]; GY (2) [143-144][286-287]; NF [13-14]; SF [230-231]; SK (2) [24-25][313-314]; TL (3) [35-36][52-53][145-146]; TR [323-324]; VL (2) [130-131][288-289], VW [233-234] | VL(2) [130-131] [288-289] |
| GAPDH | EK [256-257]; GY (2) [281-282][327-328]; HY [39-40]; MF [44-45]; QY [46-47]; VK [316-317] | - |
| LDHA | GL [287-288]; GY [254-255]; IL [195-196]; MK [41-42]; NL (3) [110-111] [219-220][274-275]; NR [160-161]; PK [158-159]; QF [341-342]; TY [148-149] | IL [195-196] |
| PGK1 | AL(2) [206-207][364-365]; SL (2) [79-80][89-90]; EW [354-355]; MK [196-197]; NY [201-202]; PF [213-214]; PK [188-189]; SK [159-160]; TF [251-252]; TL [8-9]; VK [144-145]; VL (2) [255-256][419-420] | VL(2) [255-256] [419-420] |
| PYGM | HL(2) [35-36][412-413]; EK (3) [79-80][197-198][375-376]; AL (2) [55-56][106-107]; SL [89-90]; GL (3) [18-19][119-120] [156-157]; PL [4-5]; AW [377-378]; AY [57-58]; DG (3) [43-44][277-278][758-759]; EW [823-824]; EY [571-572]; GY (2) [161-162][750-751]; HF [37-38]; IL [363-364]; IR (2) [71-72] [548-549]; ML [705-706]; MR [361-362]; NF [31-32]; NL (2) [262-263][279-280]; NR (3) [33-34][425-426][607-608]; QL (2) [117-118][595-596]; SY [540-541]; TL (3) [39-40][97-98][204-205]; TR [825-826]; VF [798-799]; VK (2) [41-42][554-555]; VL [287-288] | VL [287-288]; IL [363-364] |
| ALDOA | AL [100-101]; VL [93-94] | VL [93-94] |
| Peptide | Molecular Weight (gmol−1) | Isoelectric Point | Net charge | Solubility a | Bioactivity Score b | Peptide | Molecular Weight (gmol−1) | Isoelectric Point | Net Charge | Solubility a | Bioactivity Score b |
|---|---|---|---|---|---|---|---|---|---|---|---|
| AF | 236.27 | 3.77 | 0 | - | 0.97 | NR | 288.30 | 10.42 | 1 | + | 0.26 |
| AL | 202.25 | 3.70 | 0 | - | 0.44 | NY | 295.29 | 3.24 | 0 | - | 0.22 |
| AW | 275.30 | 3.66 | 0 | - | 0.97 | PL | 228.29 | 4.08 | 0 | - | 0.81 |
| AY | 252.27 | 3.66 | 0 | - | 0.35 | PF | 262.30 | 4.15 | 0 | - | 0.99 |
| DG | 190.15 | 0.68 | −1 | + | 0.39 | PK | 243.30 | 10.57 | 1 | + | 0.33 |
| DR | 289.29 | 6.68 | 0 | + | 0.29 | PW | 301.34 | 4.04 | 0 | - | 0.99 |
| EK | 275.30 | 6.85 | 0 | + | 0.02 | PY | 278.30 | 3.85 | 0 | - | 0.74 |
| EW | 333.34 | 0.88 | −1 | + | 0.59 | SF | 252.27 | 3.43 | 0 | - | 0.95 |
| EY | 310.30 | 0.95 | −1 | + | 0.07 | SK | 233.27 | 9.86 | 1 | + | 0.07 |
| GF | 222.24 | 3.70 | 0 | - | 0.99 | SL | 218.25 | 3.37 | 0 | - | 0.33 |
| GL | 188.22 | 3.63 | 0 | - | 0.81 | SY | 268.27 | 3.39 | 0 | - | 0.26 |
| GY | 238.24 | 3.61 | 0 | - | 0.74 | SW | 219.30 | 3.34 | 0 | - | 0.93 |
| HF | 302.33 | 7.56 | 0 | - | 0.95 | TF | 266.29 | 3.36 | 0 | - | 0.83 |
| HL | 268.31 | 7.56 | 0.1 | - | 0.37 | TK | 247.29 | 9.28 | 1 | + | 0.03 |
| HR | 311.34 | 10.59 | 1.1 | + | 0.33 | TL | 232.28 | 3.32 | 0 | - | 0.14 |
| HW | 341.36 | 7.56 | 0.1 | - | 0.95 | TR | 275.31 | 10.53 | 1 | + | 0.13 |
| HY | 318.33 | 7.54 | 0.1 | - | 0.30 | TY | 282.29 | 3.35 | 0 | - | 0.11 |
| IL | 244.33 | 3.64 | 0 | - | 0.39 | QF | 293.32 | 3.41 | 0 | - | 0.95 |
| IR | 287.36 | 10.85 | 1 | + | 0.33 | QL | 259.30 | 3.34 | 0 | - | 0.29 |
| IW | 317.38 | 3.61 | 0 | - | 0.94 | QY | 309.32 | 3.36 | 0 | - | 0.23 |
| MF | 296.39 | 3.45 | 0 | - | 1.00 | VF | 264.32 | 3.67 | 0 | - | 0.82 |
| MK | 277.39 | 9.88 | 1 | + | 0.45 | VK | 245.32 | 10.10 | 1 | + | 0.03 |
| ML | 262.37 | 3.38 | 0 | - | 0.89 | VL | 230.30 | 3.60 | 0 | - | 0.13 |
| MR | 305.40 | 10.59 | 1 | + | 0.85 | VR | 273.33 | 10.10 | 1 | + | 0.11 |
| MW | 335.42 | 3.35 | 0 | - | 1.00 | VW | 303.36 | 3.57 | 0 | - | 0.80 |
| NF | 279.29 | 3.28 | 0 | - | 0.94 | VY | 280.32 | 3.59 | 0 | - | 0.10 |
| NL | 245.28 | 3.21 | 0 | - | 0.29 | VPL | 327.42 | 3.60 | 0 | - | 0.37 |
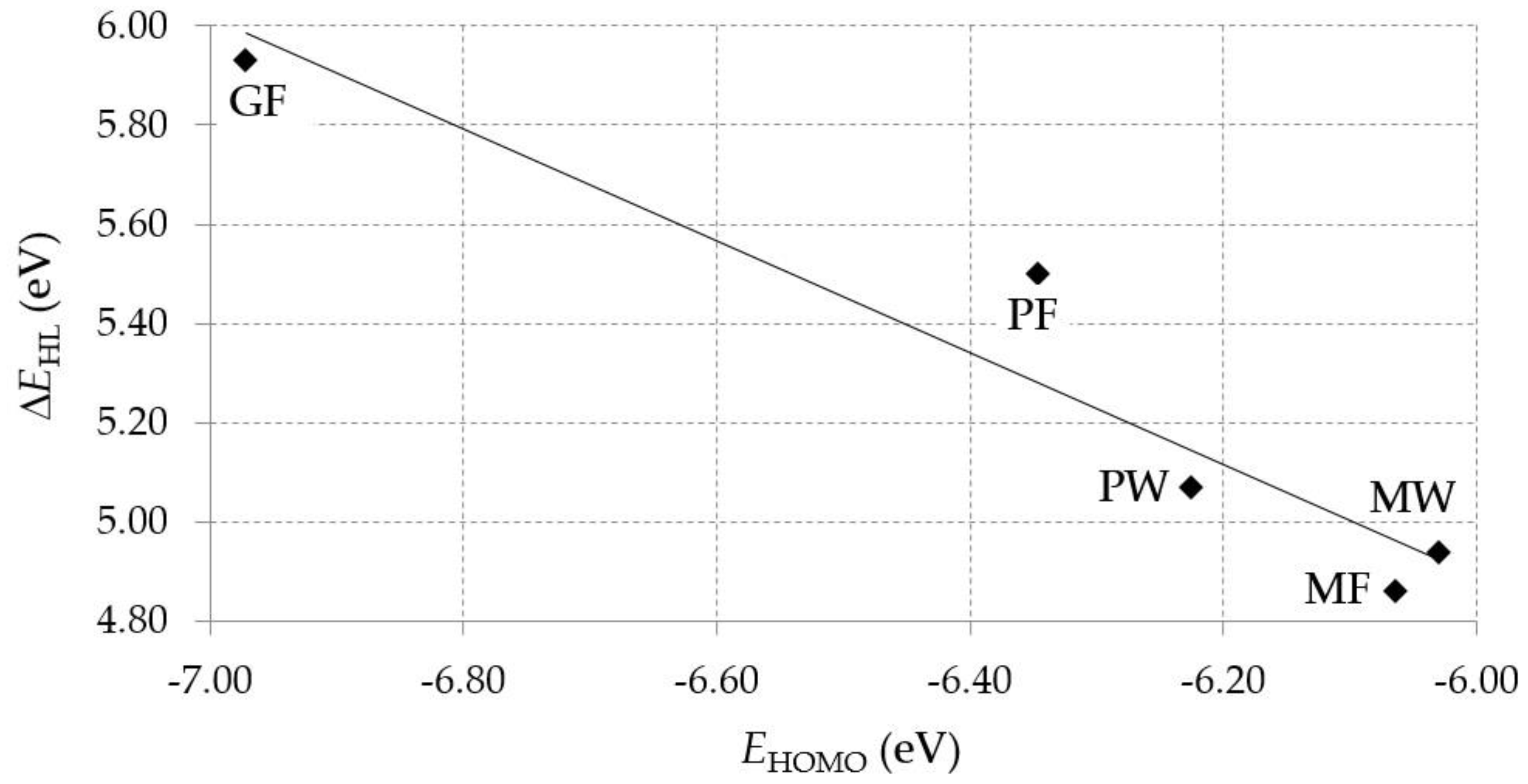
| Dipeptide | HOMO, E (eV) | LUMO, E (eV) | ∆EHL (eV) |
|---|---|---|---|
GF | −6.97 | −1.04 | 5.93 |
MW | −6.03 | −1.09 | 4.94 |
MF | −6.06 | −1.20 | 4.86 |
PP | −6.35 | −0.85 | 5.50 |
PW | −6.23 | −1.16 | 5.07 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kęska, P.; Stadnik, J.; Bąk, O.; Borowski, P. Meat Proteins as Dipeptidyl Peptidase IV Inhibitors and Glucose Uptake Stimulating Peptides for the Management of a Type 2 Diabetes Mellitus In Silico Study. Nutrients 2019, 11, 2537. https://doi.org/10.3390/nu11102537
Kęska P, Stadnik J, Bąk O, Borowski P. Meat Proteins as Dipeptidyl Peptidase IV Inhibitors and Glucose Uptake Stimulating Peptides for the Management of a Type 2 Diabetes Mellitus In Silico Study. Nutrients. 2019; 11(10):2537. https://doi.org/10.3390/nu11102537
Chicago/Turabian StyleKęska, Paulina, Joanna Stadnik, Olga Bąk, and Piotr Borowski. 2019. "Meat Proteins as Dipeptidyl Peptidase IV Inhibitors and Glucose Uptake Stimulating Peptides for the Management of a Type 2 Diabetes Mellitus In Silico Study" Nutrients 11, no. 10: 2537. https://doi.org/10.3390/nu11102537
APA StyleKęska, P., Stadnik, J., Bąk, O., & Borowski, P. (2019). Meat Proteins as Dipeptidyl Peptidase IV Inhibitors and Glucose Uptake Stimulating Peptides for the Management of a Type 2 Diabetes Mellitus In Silico Study. Nutrients, 11(10), 2537. https://doi.org/10.3390/nu11102537