Quantification of Human Milk Phospholipids: the Effect of Gestational and Lactational Age on Phospholipid Composition

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Chemicals and Reagents
2.3. Preparation of Standards
2.4. Milk Sample Collection
2.5. Liquid-Liquid Extraction of Phospholipids
2.6. Analysis of Phospholipid Species
2.7. Preprocessing of Data and Statistical Analysis
3. Results
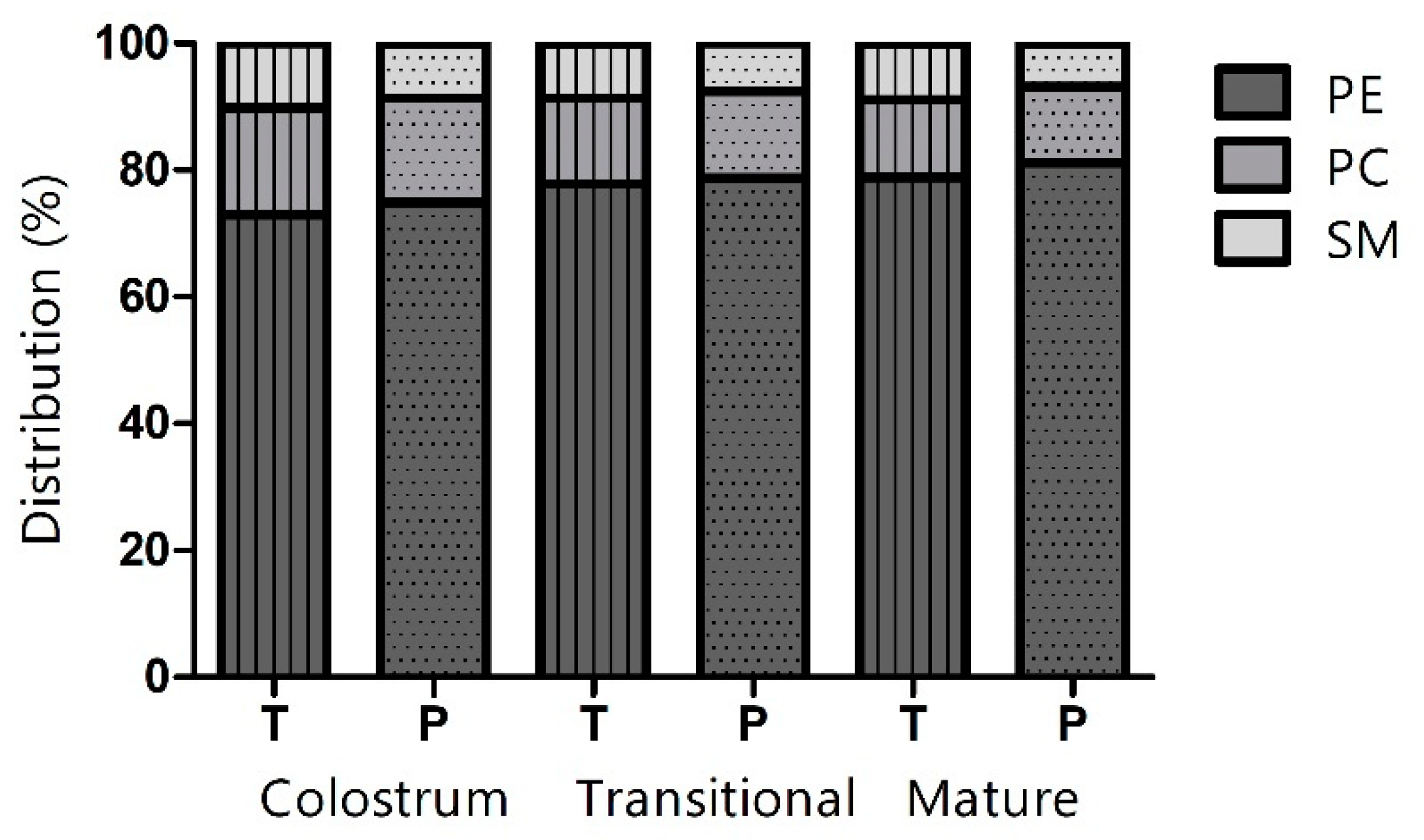
3.1. Percentage Distribution of Phospholipids
3.2. Total Concentration of Phospholipids
3.3. Concentrations of Individual Phospholipid Species
3.4. Exploratory Analysis of Preterm and Term Milk
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cai, X.; Wardlaw, T.; Brown, D.W. Global Trends in Exclusive Breastfeeding. Int. Breastfeed. J. 2012, 7, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Bridgman, S.L.; Konya, T.; Azad, M.B.; Sears, M.R.; Becker, A.B.; Turvey, S.E.; Mandhane, P.J.; Subbarao, P.; CHILD Study Investigators; Scott, J.A.; et al. Infant Gut Immunity: A Preliminary Study of IgA Associations with Breastfeeding. J. Dev. Orig. Health Dis. 2016, 7, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Delplanque, B.; Gibson, R.; Koletzko, B.; Lapillonne, A.; Strandvik, B. Lipid Quality in Infant Nutrition: Current Knowledge and Future Opportunities. J. Pediatr. Gastroenterol. Nutr. 2015, 61, 8–17. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the Essential Composition of Infant and Follow-on Formulae. EFSA J. 2014, 12, 3760. [Google Scholar] [CrossRef]
- Shoji, H.; Shimizu, T.; Kaneko, N.; Shinohara, K.; Shiga, S.; Saito, M.; Oshida, K.; Shimizu, T.; Takase, M.; Yamashiro, Y. Comparison of the Phospholipid Classes in Human Milk in Japanese Mothers of Term and Preterm Infants. Acta Paediatr. 2006, 95, 996–1000. [Google Scholar] [CrossRef] [PubMed]
- Gallier, S.; Gragson, D.; Cabral, C.; Jiménez-Flores, R.; Everett, D.W. Composition and Fatty Acid Distribution of Bovine Milk Phospholipids From Processed Milk Products. J. Agric. Food Chem. 2010, 58, 10503–10511. [Google Scholar] [CrossRef] [PubMed]
- Blans, K.; Hansen, M.S.; Sørensen, L.V.; Hvam, M.L.; Howard, K.A.; Möller, A.; Wiking, L.; Larsen, L.B.; Rasmussen, J.T. Pellet-free Isolation of Human and Bovine Milk Extracellular Vesicles by Size-Exclusion Chromatography. J. Extracell. Vesicles 2017, 6, 1–16. [Google Scholar] [CrossRef]
- Sæle, Ø.; Rød, K.E.L.; Quinlivan, V.H.; Li, S.; Farber, S.A. A Novel System to Quantify Intestinal Lipid Digestion and Transport. BBA Mol. Cell Biol. Lipids 2018, 1863, 948–957. [Google Scholar] [CrossRef]
- Wang, L.; Shimizu, Y.; Kaneko, S.; Hanaka, S.; Abe, T.; Shimasaki, H.; Hisaki, H.; Nakajima, H. Comparison of the Fatty Acid Composition of Total Lipids and Phospholipids in Breast Milk from Japanese Women. Pediatr. Int. 2000, 42, 14–20. [Google Scholar] [CrossRef]
- Moukarzel, S.; Dyer, R.A.; Keller, B.O.; Elango, R.; Innis, S.M. Human Milk Plasmalogens Are Highly Enriched in Long-Chain PUFAs. J. Nutr. 2016, 146, 2412–2417. [Google Scholar] [CrossRef]
- Maas, C.; Franz, A.R.; Shunova, A.; Mathes, M.; Bleeker, C.; Poets, C.F.; Schleicher, E.; Bernhard, W. Choline and Polyunsaturated Fatty Acids in Preterm Infants’ Maternal Milk. Eur. J. Nutr. 2017, 56, 1733–1742. [Google Scholar] [CrossRef] [PubMed]
- Cilla, A.; Quintaes, K.D.; Barberá, R.; Alegría, A. Phospholipids in Human Milk and Infant Formulas: Benefits and Needs for Correct Infant Nutrition. Crit. Rev. Food Sci. Nutr. 2016, 56, 1880–1892. [Google Scholar] [CrossRef] [PubMed]
- Gidrewicz, D.A.; Fenton, T.R. A Systematic Review and Meta-Analysis of the Nutrient Content of Preterm and Term Breast Milk. BMC Pediatr. 2014, 14, 216. [Google Scholar] [CrossRef] [PubMed]
- Lemons, J.A.; Moye, L.; Hall, D.; Simmons, M. Differences in the Composition of Preterm and Term Human Milk During Early Lactation. Pediatr. Res. 1982, 16, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Sundekilde, U.K.; Downey, E.; O’Mahony, J.; O’Shea, C.; Ryan, C.A.; Kelly, A.; Bertram, H.C. The Effect of Gestational and Lactational Age on the Human Milk Metabolome. Nutrients 2016, 8, 304. [Google Scholar] [CrossRef] [PubMed]
- Guerra, E.; Downey, E.; O’Mahony, J.A.; Caboni, M.F.; O’Shea, C.; Ryan, A.C.; Kelly, A.L. Influence of Duration of Gestation on Fatty Acid Profiles of Human Milk. Eur. J. Lipid Sci. Technol. 2016, 118, 1775–1787. [Google Scholar] [CrossRef]
- Spevacek, A.R.; Smilowitz, J.T.; Chin, E.L.; Underwood, M.A.; German, J.B.; Slupsky, C.M. Infant Maturity at Birth Reveals Minor Differences in the Maternal Milk Metabolome in the First Month of Lactation. J. Nutr. 2015, 145, 1698–1708. [Google Scholar] [CrossRef] [Green Version]
- Bligh, E.G.; Dyer, W.J. A Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Zhao, Y.Y.; Xiong, Y.; Curtis, J.M. Measurement of Phospholipids by Hydrophilic Interaction Liquid Chromatography Coupled to Tandem Mass Spectrometry: The Determination of Choline Containing Compounds in Foods. J. Chromatogr. A 2011, 1218, 5470–5479. [Google Scholar] [CrossRef]
- Sala-Vila, A.; Castellote, A.I.; Rodriguez-Palmero, M.; Campoy, C.; López-Sabater, M.C. Lipid Composition in Human Breast Milk from Granada (Spain): Changes During Lactation. Nutrition 2005, 21, 467–473. [Google Scholar] [CrossRef]
- Bitman, J.; Wood, L.; Hamosh, M.; Mehta, N.R. Comparison of the Phospholipid Composition of Breast Milk from Mothers of Term and Preterm Infants During Lactation. Am. J. Clin. Nutr. 1984, 40, 1103–1119. [Google Scholar] [CrossRef] [PubMed]
- Benoit, B.; Fauquant, C.; Daira, P.; Peretti, N.; Guichardant, M.; Michalski, M.C. Phospholipid Species and Minor Sterols in French Human Milks. Food Chem. 2010, 120, 684–691. [Google Scholar] [CrossRef]
- Claumarchirant, L.; Cilla, A.; Matencio, E.; Sanchez-Siles, L.M.; Castro-Gomez, P.; Fontecha, J.; Alegría, A.; Lagarda, M.J. Addition of Milk Fat Globule Membrane as an Ingredient of Infant Formulas for Resembling the Polar Lipids of Human Milk. Int. Dairy J. 2016, 61, 228–238. [Google Scholar] [CrossRef]
- Ma, L.; MacGibbon, A.K.H.; Jan Mohamed, H.J.B.; Loy, S.L.; Rowan, A.; McJarrow, P.; Fong, B.Y. Determination of phospholipid concentrations in breast milk and serum using a high performance liquid chromatography–mass spectrometry–multiple reaction monitoring method. Int. Dairy J. 2017, 71, 50–59. [Google Scholar] [CrossRef]
- Smoczynski, M. Role of Phospholipid Flux during Milk Secretion in the Mammary Gland. J. Mammary Gland Biol. Neoplasia 2017, 22, 117–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Wang, C.; Han, X. Selection of Internal Standards for Accurate Quantification of Complex Lipid Species in Biological Extracts by Electrospray Ionization Mass Spectrometry—What, How and Why? Mass Spectrom. Rev. 2017, 36, 693–714. [Google Scholar] [CrossRef] [PubMed]
- Koletzko, B.; Rodriguez-palmero, M. Polyunsaturated Fatty Acids in Human Milk and Their Role in Early Infant Development. J. Mammary Gland Boil. Neoplasia 1999, 4, 269–284. [Google Scholar] [CrossRef]
- Simonin, C.; Rüegg, M.; Sidiropoulos, D. Comparison of the fat content and fat globule size distribution of breast milk from mothers delivering term and preterm. Am. J. Clin. Nutr. 1984, 40, 820–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, H.; Gallier, S. Nature’s Complex Emulsion: The Fat Globules of Milk. Food Hydrocoll. 2017, 68, 81–89. [Google Scholar] [CrossRef]
- Lopez, C.; Briard-Bion, V.; Ménard, O.; Beaucher, E.; Rousseau, F.; Fauquant, J.; Leconte, N.; Robert, B. Fat Globules Selected from Whole Milk According to Their Size: Different Compositions and Structure of the Biomembrane, Revealing Sphingomyelin-rich Domains. Food Chem. 2011, 125, 355–368. [Google Scholar] [CrossRef]
- González, H.F.; Visentin, S. Nutrients and Neurodevelopment: Lipids. Update. Arch. Argent. Pediatr. 2016, 114, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Campoy, C.; Escolano-Margarit, V.; Anjos, T.; Szajewska, H.; Uauy, R. Omega 3 Fatty Acids on Child Growth, Visual Acuity and Neurodevelopment. Br. J. Nutr. 2012, 107, S85–S106. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.C.; Zeisel, S.H.; Mar, M.; Sadler, T.W. Inhibitors of Choline Uptake and Metabolism Cause Developmental Abnormalities in Neurulating Mouse Embryos. Teratology 2001, 64, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Bernhard, W.; Full, A.; Arand, J.; Maas, C.; Poets, C.F.; Franz, A.R. Choline Supply of Preterm Infants: Assessment of Dietary Intake and Pathophysiological Considerations. Eur. J. Nutr. 2013, 52, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Bernhard, W.; Raith, M.; Kunze, R.; Koch, V.; Heni, M.; Maas, C.; Abele, H.; Poets, C.F.; Franz, A.R. Choline Concentrations are Lower in Postnatal Plasma of Preterm Infants Than in Cord Plasma. Eur. J. Nutr. 2015, 54, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Bauer, J.; Gerss, J. Longitudinal Analysis of Macronutrients and Minerals in Human Milk Produced by Mothers of Preterm Infants. Clin. Nutr. 2011, 30, 215–220. [Google Scholar] [CrossRef]
- Tudehope, D.I. Human milk and the nutritional needs of preterm infants. J. Pediatr. 2013, 162, S17–S25. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.T.; Martin, C.R. Fatty acid requirements for the preterm infant. Semin. Fetal Neonatal Med. 2017, 22, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Caudill, M.A. Pre- and Postnatal Health: Evidence of Increased Choline Needs. J. Am. Diet. Assoc. 2010, 110, 1198–1206. [Google Scholar] [CrossRef]
- Sabel, K.G.; Lundqvist-Persson, C.; Bona, E.; Petzold, M.; Strandvik, B. Fatty acid patterns early after premature birth, simultaneously analysed in mothers’ food, breast milk and serum phospholipids of mothers and infants. Lipids Health Dis. 2009, 8, 1–15. [Google Scholar] [CrossRef]
- Ballard, O.; Morrow, A.L. Human Milk Composition: Nutrients and Bioactive Factors. Pediatr. Clin. N. Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, A.K.; Löfqvist, C.; Najm, S.; Hellgren, G.; Sävman, K.; Andersson, M.X.; Smith, L.E.H.; Hellström, A. Long-chain polyunsaturated fatty acids decline rapidly in milk from mothers delivering extremely preterm indicating the need for supplementation. Acta Paediatr. Int. J. Paediatr. 2018, 1–8. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Phospholipid Class | Preterm | Term | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Colostrum | Transitional | Mature | Colostrum | Transitional | Mature | |||||||
| PE | 87.85 | (27.30) A,a | 75.25 | (24.85) A,a | 46.64 | (19.81) B,a | 49.40 | (13.68) A,a | 37.86 | (14.00) A,a | 29.15 | (13.04) B,a |
| PC | 19.31 | (11.46) A,a | 13.09 | (3.94) B,a | 6.92 | (3.23) C,a | 11.44 | (2.64) A,b | 6.56 | (3.26) B,b | 4.50 | (1.97) C,b |
| SM | 10.13 | (6.24) A,a | 7.21 | (1.95) B,a | 3.93 | (1.63) C,a | 6.90 | (1.26) A,b | 4.23 | (1.88) B,b | 3.29 | (1.73) C,b |
| Total PL | 117.30 | (42.08) A,a | 95.49 | (29.43) B,a | 57.49 | (23.77) C,a | 67.74 | (14.47) A,b | 48.65 | (18.11) B,b | 36.94 | (16.41) C,b |
| # | Phospholipid Species | Preterm | Term | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Colostrum | Transitional | Mature | Colostrum | Transitional | Mature | ||||||||
| 1 | PE (18:1/18:1) | 0.78 | (0.47) a | 1.09 | (0.63) a | 0.92 | (0.54) a | 0.43 | (0.26) b | 0.38 | (0.15) b | 0.52 | (0.39) b |
| 2 | PE (18:1/18:0) | >12.00 | (5.80) A,a | >12.00 | (5.53) B,a | 6.66 | (3.28) C,a | 12.00 | (4.67) A,b | 6.58 | (3.02) B,b | 3.52 | (2.10) C,b |
| 3 | PE (18:1/16:0) | 7.55 | (2.88) A,a | 4.63 | (2.03) B,a | 2.79 | (1.58) C,a | 4.46 | (2.36) A,b | 2.76 | (0.58) B,b | 1.56 | (0.85) C,b |
| 4 | PE (18:0/22:6) | 7.45 | (3.64) A,a | 7.94 | (3.55) A,a | 3.57 | (2.44) B,a | 3.60 | (1.21) A,b | 3.05 | (1.64) A,b | 1.81 | (1.34) B,b |
| 5 | PE (18:0/20:4) | 6.43 | (1.97) i | 4.09 | (1.19) ii | 2.79 | (1.13) iii | 3.05 | (1.04) ii,iii | 2.58 | (1.46) ii,iii | 2.13 | (0.96) iii |
| 6 | PE (18:0/18:2) | 6.88 | (3.13) a | 7.20 | (2.54) a | 5.80 | (2.56) a | 3.86 | (1.42) b | 4.24 | (2.50) b | 4.18 | (1.82) b |
| 7 | PE (18:0/18:1) | 2.51 | (1.11) A,a | 2.49 | (0.85) AB,a | 1.50 | (0.66) B,a | 1.46 | (0.36) A,b | 1.18 | (0.58) AB,b | 1.19 | (0.64) B,b |
| 8 | PE (18:0/16:0) | 10.16 | (3.50) A,a | 7.69 | (3.57) A,a | 4.40 | (2.07) B,a | 5.43 | (2.25) A,b | 4.71 | (1.17) A,b | 2.86 | (1.26) B,b |
| 9 | PE (16:0/22:6) | 4.99 | (2.15) A,a | 4.34 | (1.87) AB,a | 2.42 | (1.64) B,a | 2.24 | (0.63) A,b | 1.57 | (0.77) AB,b | 1.02 | (0.63) B,b |
| 10 | PE (16:0/20:4) | >12.00 | (6.97) A,a | 11.78 | (4.07) AB,a | 8.43 | (3.97) B,a | 6.57 | (2.37) A,b | 5.31 | (2.46) AB,b | 4.94 | (1.83) B,b |
| 11 | PE (16:0/18:2) | 9.00 | (3.58) a | 9.11 | (3.32) a | 6.12 | (3.14) a | 5.07 | (1.31) b | 4.50 | (2.24) b | 4.52 | (2.21) b |
| 12 | PE (16:0/18:1) | 1.99 | (1.13) a | 2.30 | (1.14) a | 1.25 | (0.91) a | 1.23 | (0.31) b | 1.01 | (0.55) b | 0.90 | (0.57) b |
| 13 | PC (18:1/20:4) | 0.41 | (0.36) i | 0.22 | (0.06) ii | 0.12 | (0.06) iii | 0.15 | (0.02) ii,iii | 0.11 | (0.06) ii,iii | 0.07 | (0.04) iii |
| 14 | PC (18:0/18:2) | 1.25 | (0.68) A,a | 1.01 | (0.27) AB,a | 0.66 | (0.25) B,a | 0.67 | (0.13) A,b | 0.63 | (0.41) AB,b | 0.57 | (0.29) B,b |
| 15 | PC (18:0/18:1) | 0.60 | (0.38) i | 0.40 | (0.11) ii | 0.21 | (0.08) iii | 0.33 | (0.05) ii | 0.20 | (0.12) ii,iii | 0.19 | (0.10) iii |
| 16 | PC (16:0/20:5) | 1.97 | (1.46) A,a | 1.42 | (0.47) AB,a | 0.80 | (0.49) B,a | 0.76 | (0.13) A,b | 0.64 | (0.33) AB,b | 0.46 | (0.26) B,b |
| 17 | PC (16:0/20:4) | 0.54 | (0.37) i | 0.35 | (0.09) ii | 0.20 | (0.11) iii | 0.20 | (0.02) ii,iii | 0.17 | (0.10) ii,iii | 0.11 | (0.05) iii |
| 18 | PC (16:0/20:3) | 1.61 | (1.05) A,a | 1.30 | (0.38) AB,a | 0.79 | (0.40) B,a | 0.77 | (0.15) A,b | 0.61 | (0.35) AB,b | 0.50 | (0.20) B,b |
| 19 | PC (16:0/18:2) | 0.61 | (0.34) A,a | 0.42 | (0.12) B,a | 0.25 | (0.10) B,a | 0.33 | (0.05) A,b | 0.20 | (0.11) B,b | 0.18 | (0.06) B,b |
| 20 | PC (16:0/18:1) | 0.55 | (0.37) A,a | 0.34 | (0.10) B,a | 0.18 | (0.08) B,a | 0.34 | (0.10) A,b | 0.17 | (0.08) B,b | 0.13 | (0.05) B,b |
| 21 | PC (16:0/16:1) | 1.09 | (0.59) A,a | 0.79 | (0.30) B,a | 0.37 | (0.18) C,a | 0.81 | (0.25) A,b | 0.38 | (0.24) B,b | 0.23 | (0.11) C,b |
| 22 | PC (16:0/16:0) | 10.68 | (6.06) A,a | 6.78 | (2.38) B,a | 3.36 | (1.66) C,a | 7.06 | (2.29) A,b | 3.44 | (1.72) B,b | 2.06 | (0.94) C,b |
| 23 | SM (18:1/23:0) | 0.22 | (0.16) | 0.19 | (0.13) | 0.15 | (0.14) | 0.13 | (0.09) | 0.12 | (0.07) | 0.09 | (0.06) |
| 24 | SM (18:1/20:0) | 1.33 | (0.98) A,a | 0.85 | (0.25) B,a | 0.48 | (0.22) B,a | 0.96 | (0.26) A,b | 0.47 | (0.21) B,b | 0.40 | (0.17) B,b |
| 25 | SM (18:1/14:0) | 0.52 | (0.28) a | 0.56 | (0.27) a | 0.31 | (0.16) a | 0.36 | (0.09) b | 0.21 | (0.02) b | 0.23 | (0.14) b |
| 26 | SM (18:0/24:0) | 0.08 | (0.08) a | 0.04 | (0.04) a | 0.03 | (0.03) a | 0.02 | (0.01) b | 0.01 | (0.01) b | 0.02 | (0.02) b |
| 27 | SM (18:0/23:0) | 0.02 | (0.05) | 0.01 | (0.01) | 0.01 | (0.01) | 0.02 | (0.01) | 0.01 | (0.004) | 0.01 | (0.005) |
| 28 | SM (18:0/22:1) | 1.35 | (0.96) A,a | 0.90 | (0.24) B,a | 0.52 | (0.19) B,a | 0.85 | (0.11) A,b | 0.52 | (0.28) B,b | 0.51 | (0.28) B,b |
| 29 | SM (18:0/22:0) | 1.29 | (0.88) A,a | 0.89 | (0.28) B,a | 0.45 | (0.22) B,a | 0.81 | (0.14) A,b | 0.47 | (0.30) B,b | 0.40 | (0.23) B,b |
| 30 | SM (18:0/20:0) | 5.02 | (2.99) A,a | 3.39 | (0.93) B,a | 1.78 | (0.76) C,a | 3.56 | (0.71) A,b | 2.27 | (1.10) B,b | 1.47 | (0.83) C,b |
| 31 | SM (18:0/14:0) | 0.29 | (0.17) a | 0.38 | (0.20) a | 0.22 | (0.11) a | 0.19 | (0.05) b | 0.15 | (0.02) b | 0.17 | (0.09) b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ingvordsen Lindahl, I.E.; Artegoitia, V.M.; Downey, E.; O’Mahony, J.A.; O’Shea, C.-A.; Ryan, C.A.; Kelly, A.L.; Bertram, H.C.; Sundekilde, U.K. Quantification of Human Milk Phospholipids: the Effect of Gestational and Lactational Age on Phospholipid Composition. Nutrients 2019, 11, 222. https://doi.org/10.3390/nu11020222
Ingvordsen Lindahl IE, Artegoitia VM, Downey E, O’Mahony JA, O’Shea C-A, Ryan CA, Kelly AL, Bertram HC, Sundekilde UK. Quantification of Human Milk Phospholipids: the Effect of Gestational and Lactational Age on Phospholipid Composition. Nutrients. 2019; 11(2):222. https://doi.org/10.3390/nu11020222
Chicago/Turabian StyleIngvordsen Lindahl, Ida Emilie, Virginia M. Artegoitia, Eimear Downey, James A. O’Mahony, Carol-Anne O’Shea, C. Anthony Ryan, Alan L. Kelly, Hanne C. Bertram, and Ulrik K. Sundekilde. 2019. "Quantification of Human Milk Phospholipids: the Effect of Gestational and Lactational Age on Phospholipid Composition" Nutrients 11, no. 2: 222. https://doi.org/10.3390/nu11020222
APA StyleIngvordsen Lindahl, I. E., Artegoitia, V. M., Downey, E., O’Mahony, J. A., O’Shea, C. -A., Ryan, C. A., Kelly, A. L., Bertram, H. C., & Sundekilde, U. K. (2019). Quantification of Human Milk Phospholipids: the Effect of Gestational and Lactational Age on Phospholipid Composition. Nutrients, 11(2), 222. https://doi.org/10.3390/nu11020222