Hypoglycemic and Hypolipidemic Effects of Phellinus Linteus Mycelial Extract from Solid-State Culture in A Rat Model of Type 2 Diabetes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Preparation of P. linteus Extract (PLE)
2.3. Component Analysis of PLE
2.4. Animals
2.5. Induction of Type 2 Diabetes and Experimental Design
2.6. Collection and Preservation of Experimental Samples
2.7. Serum Biochemical Indexes
2.8. Hepatic TG and T-CHO
2.9. RNA Extraction from Liver, and Quantitative Real-Time Polymerase Chain Reaction (Qrt-PCR)
2.10. Statistical Analysis
3. Results
3.1. Main Components of PLE
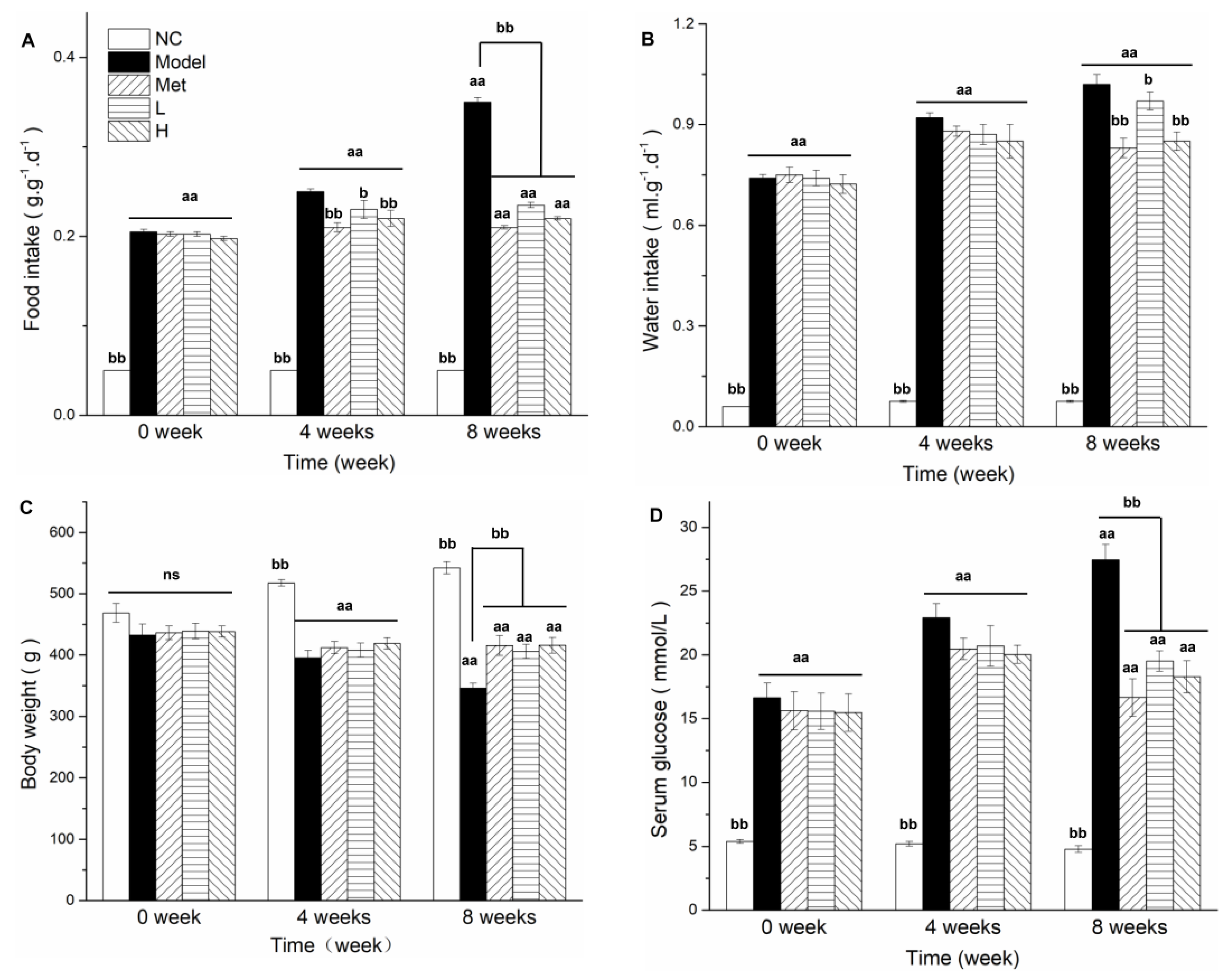
3.2. PLE and Metformin Treatments Inhibited Overeating, Body Weight Loss, and FBG Level in Diabetic Rat Model
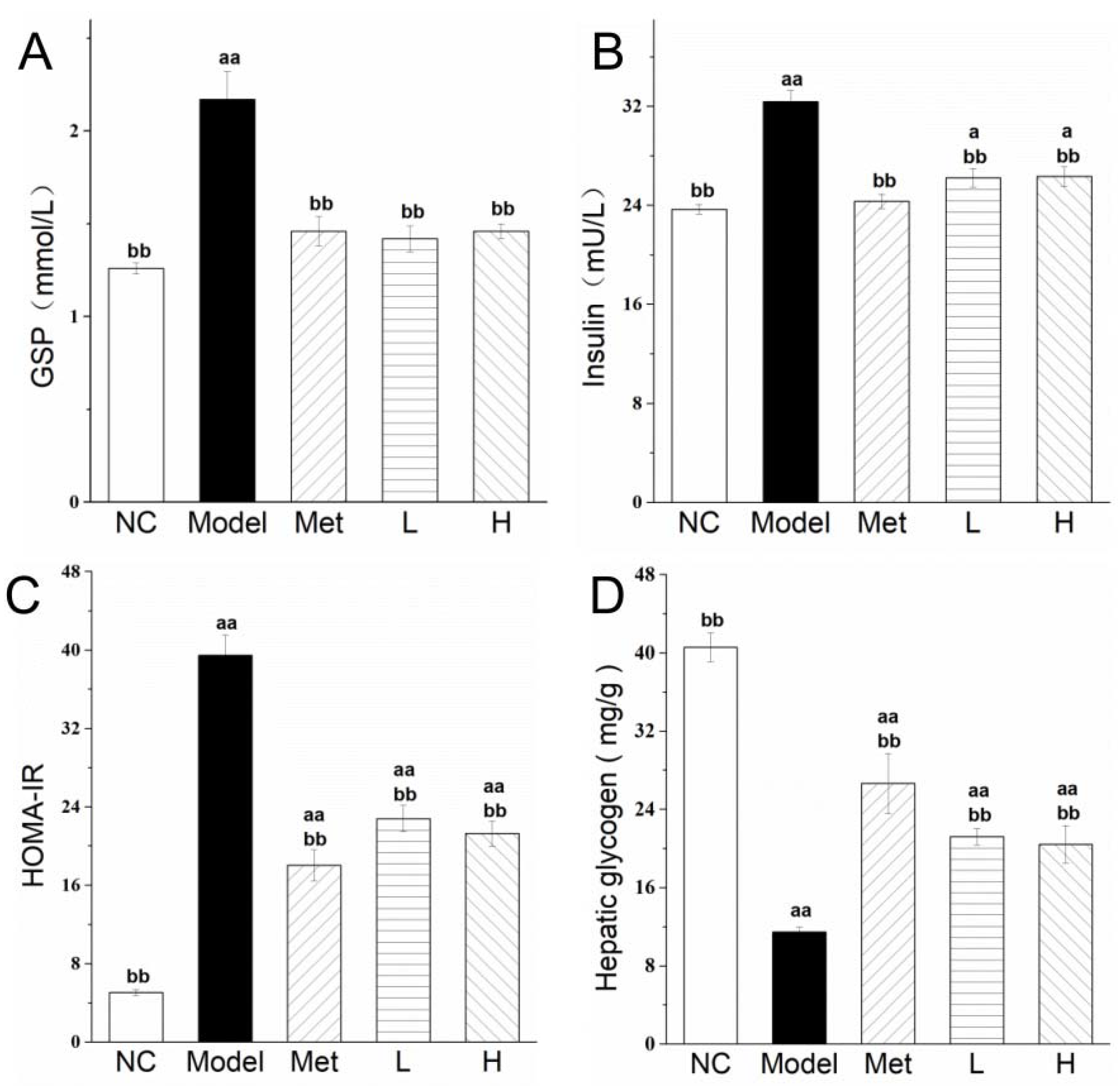
3.3. PLE and Metformin Treatments Reduced Glycosylated Serum Protein (GSP) Level, and Improved Insulin Resistance and Promoted Hepatic Glycogen Storage
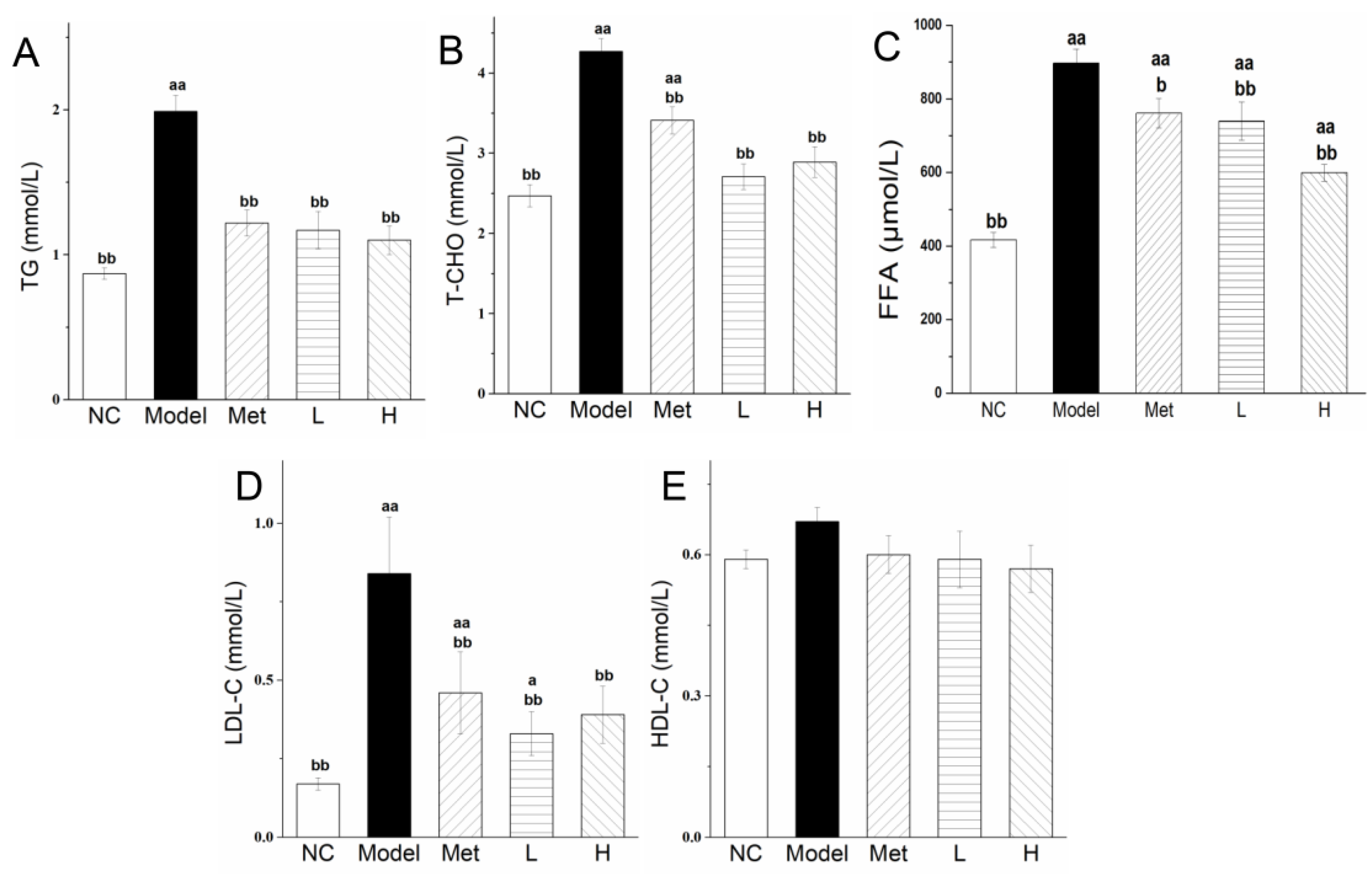
3.4. PLE and Metformin Treatments Improved Serum Lipid and Lipoprotein Profiles
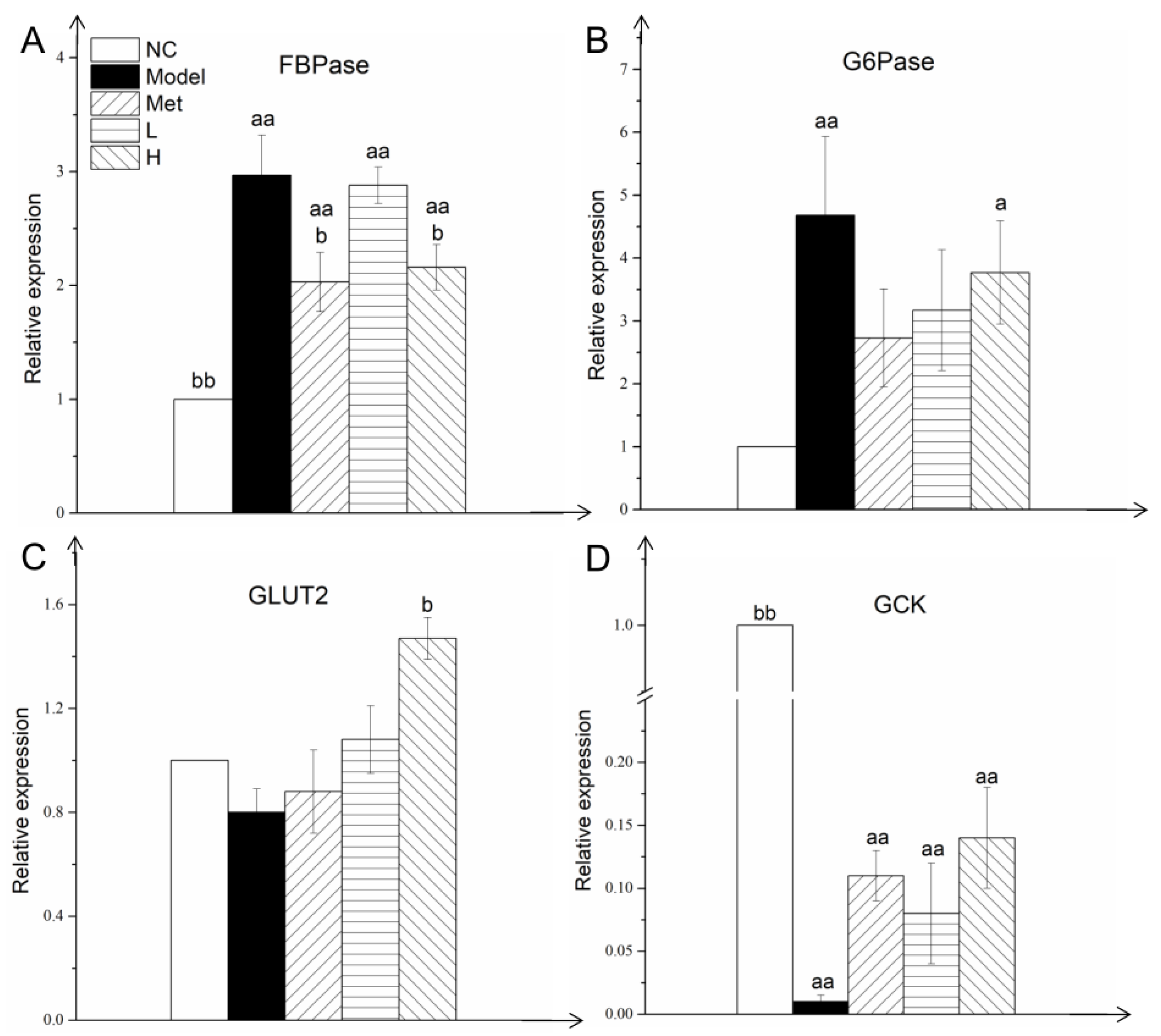
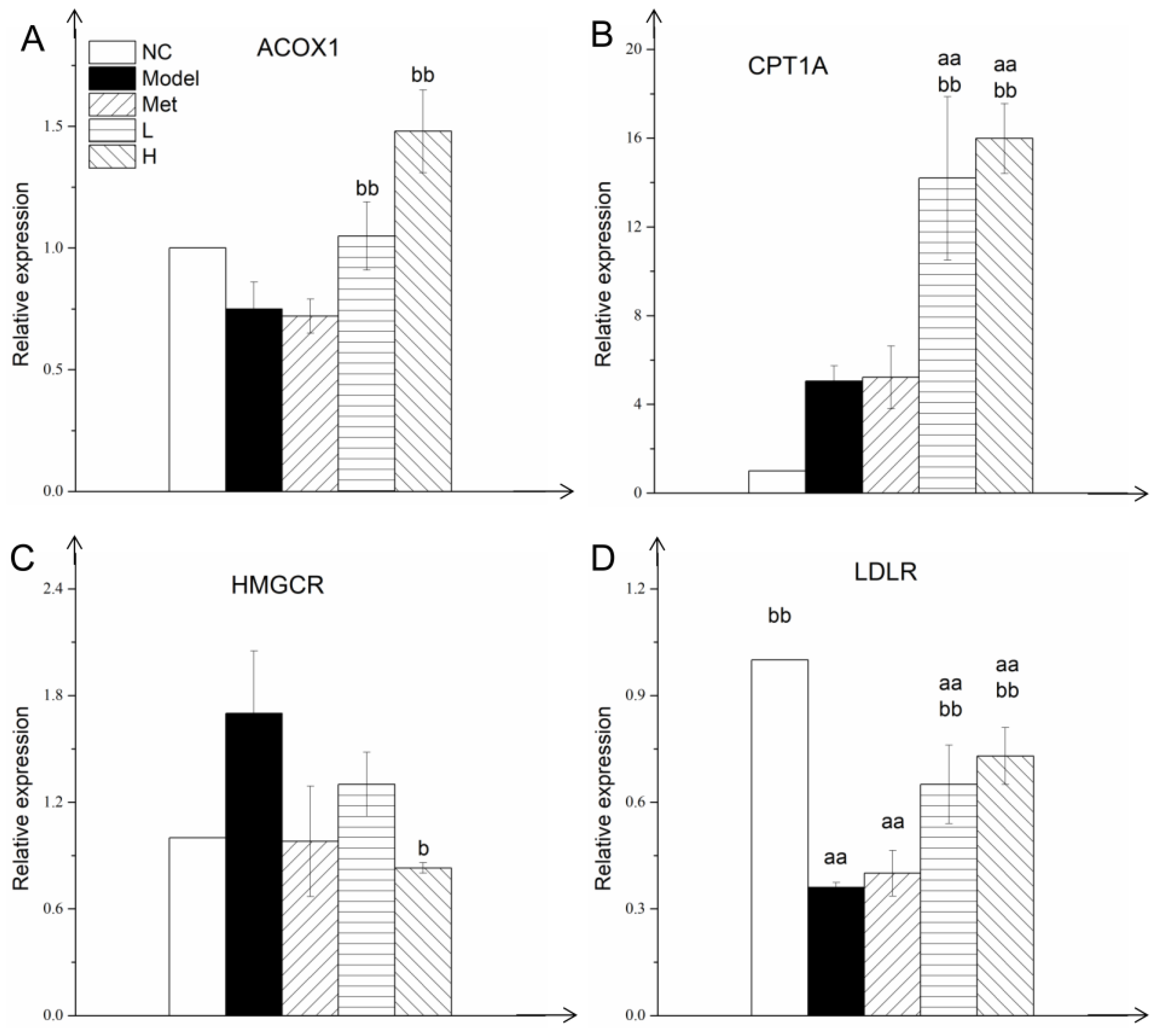
3.5. PLE and Metformin Regulated Expression of Key Gluconeogenesis and Glycolysis Enzymes in Liver
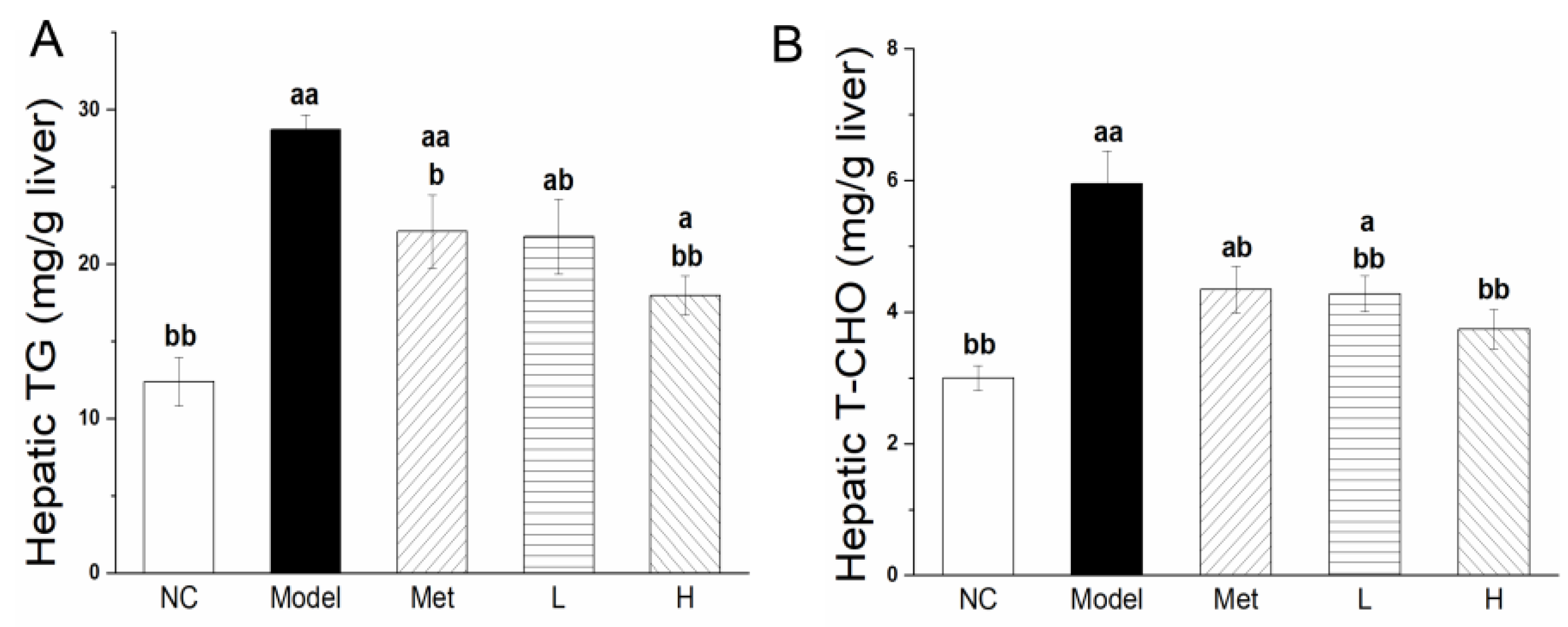
3.6. PLE and Metformin Decreased Hepatic TG and T-CHO Contents and Regulated Expression of Key Enzymes of Lipid Metabolism in Liver
3.7. PLE and Metformin Ameliorate Liver Injury and Kidney Injury
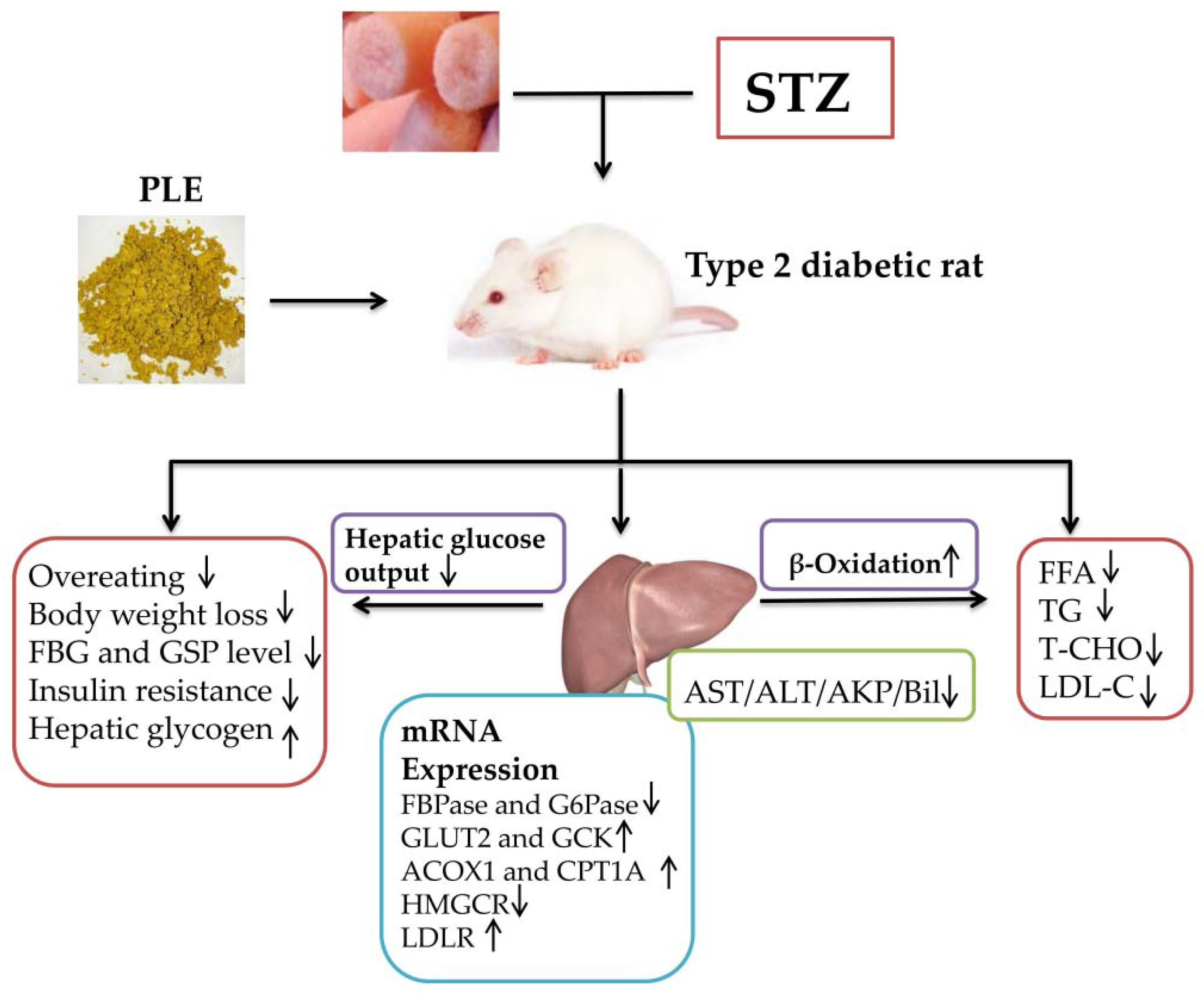
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Amer Diabet, A. Diagnosis and classification of diabetes mellitus. Diabetes Care 2009, 32, S62–S67. [Google Scholar]
- Dey, A.; Swaminathan, K. Hyperglycemia-induced mitochondrial alterations in liver. Life Sci. 2010, 87, 197–214. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Valades, A.G.; Mendez-Lucas, A.; Vidal-Alabro, A.; Blasco, F.X.; Chillon, M.; Bartrons, R.; Bermudez, J.; Perales, J.C. Pck1 gene silencing in the liver improves glycemia control, insulin sensitivity, and dyslipidemia in db/db mice. Diabetes 2008, 57, 2199–2210. [Google Scholar] [CrossRef] [PubMed]
- Rines, A.K.; Sharabi, K.; Tavares, C.D.J.; Puigserver, P. Targeting hepatic glucose metabolism in the treatment of type 2 diabetes. Nat. Rev. Drug Discov. 2016, 15, 786–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.H.; Chien, Y.W.; Chang, M.L.; Hou, C.C.; Chan, C.H.; Tang, H.W.; Huang, H.Y. Taiwanese green propolis ethanol extract delays the progression of type 2 diabetes mellitus in rats treated with streptozotocin/high-fat diet. Nutrients 2018, 10, 503. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, T.; Shimoda, M.; Sanada, J.; Fushimi, Y.; Hirata, Y.; Irie, S.; Obata, A.; Kimura, T.; Hirukawa, H.; Kohara, K.; et al. There is a close association between the recovery of liver injury and glycemic control after sglt2 inhibitor treatment in japanese subjects with type 2 diabetes: A retrospective clinical study. Diabetes Ther. 2018, 9, 1569–1580. [Google Scholar] [CrossRef]
- Xia, X.A.; Yan, J.H.; Shen, Y.F.; Tang, K.X.; Yin, J.; Zhang, Y.H.; Yang, D.J.; Liang, H.; Ye, J.P.; Weng, J.P. Berberine improves glucose metabolism in diabetic rats by inhibition of hepatic gluconeogenesis. PLoS ONE 2011, 6, e16556. [Google Scholar] [CrossRef]
- Morton, N.M.; Holmes, M.C.; Fievet, C.; Staels, B.; Tailleux, A.; Mullins, J.J.; Seckl, J.R. Improved lipid and lipoprotein profile, hepatic insulin sensitivity, and glucose tolerance in 11 beta-hydroxysteroid dehydrogenase type 1 null mice. J. Biol. Chem. 2001, 276, 41293–41300. [Google Scholar] [CrossRef]
- Totland, G.K.; Madsen, L.; Klementsen, B.; Vaagenes, H.; Kryvi, H.; Froyland, L.; Hexeberg, S.; Berge, R.K. Proliferation of mitochondria and gene expression of carnitine palmitoyltransferase and fatty acyl-coa oxidase in rat skeletal muscle, heart and liver by hypolipidemic fatty acids. Biol. Cell 2000, 92, 317–329. [Google Scholar] [CrossRef]
- Saydah, S.H.; Fradkin, J.; Cowie, C.C. Poor control of risk factors for vascular disease among adults with previously diagnosed diabetes. JAMA-J. Am. Med. Assoc. 2004, 291, 335–342. [Google Scholar] [CrossRef]
- You, Y.M.; Ren, T.; Zhang, S.Q.; Shirima, G.G.; Cheng, Y.J.; Liu, X. Hypoglycemic effects of Zanthoxylum alkylamides by enhancing glucose metabolism and ameliorating pancreatic dysfunction in streptozotocin-induced diabetic rats. Food Funct. 2015, 6, 3144–3154. [Google Scholar] [CrossRef]
- Lindequist, U.; Niedermeyer, T.H.J.; Julich, W.D. The pharmacological potential of mushrooms. Evid.-Based Complement Altern. Med. 2005, 2, 285–299. [Google Scholar] [CrossRef]
- Chang, C.J.; Lin, C.S.; Lu, C.C.; Martel, J.; Ko, Y.F.; Ojcius, D.M.; Tseng, S.F.; Wu, T.R.; Chen, Y.Y.M.; Young, J.D.; et al. Ganoderma lucidum reduces obesity in mice by modulating the composition of the gut microbiota. Nat. Commun. 2015, 6, 7489. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Bao, L.; Zhou, N.; Zhang, J.J.; Liao, M.F.; Zheng, Z.Y.; Wang, Y.J.; Liu, C.; Wang, J.; Wang, L.F.; et al. Structural modification of natural product ganomycin i leading to discovery of a a-glucosidase and HMG-CoA reductase dual inhibitor improving obesity and metabolic dysfunction in vivo. J. Med. Chem. 2018, 61, 3609–3625. [Google Scholar] [CrossRef]
- Wang, J.; Wang, C.; Li, S.Q.; Li, W.W.; Yuan, G.Q.; Pan, Y.X.; Chen, H.X. Anti-diabetic effects of Inonotus obliquus polysaccharides in streptozotocin-induced type 2 diabetic mice and potential mechanism via PI3K-Akt signal pathway. Biomed. Pharmacother. 2017, 95, 1669–1677. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Wu, Q.P.; Zhang, J.M.; Xie, Y.Z.; Cai, W.; Tan, J.B. Antidiabetic activity of Ganoderma lucidum polysaccharides F31 down-regulated hepatic glucose regulatory enzymes in diabetic mice. J. Ethnopharmacol. 2017, 196, 47–57. [Google Scholar] [CrossRef]
- Zhao, C.; Liao, Z.S.; Wu, X.Q.; Liu, Y.L.; Liu, X.Y.; Lin, Z.X.; Huang, Y.F.; Liu, B. Isolation, purification, and structural features of a polysaccharide from Phellinus linteus and its hypoglycemic effect in alloxan-induced diabetic mice. J. Food Sci. 2014, 79, H1002–H1010. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.B.; Kim, S.H.; Chen, C.Y. A medicinal mushroom: Phellinus linteus. Curr. Med. Chem. 2008, 15, 1330–1335. [Google Scholar] [CrossRef]
- Chen, H.; Tian, T.; Miao, H.; Zhao, Y.Y. Traditional uses, fermentation, phytochemistry and pharmacology of Phellinus linteus: A review. Fitoterapia 2016, 113, 6–26. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.X.; Zhu, H.; Hu, Q.M.; Liu, Y.Y.; Zhao, S.M.; Peng, N.; Liang, Y.X. A novel polysaccharide from mycelia of cultured Phellinus linteus displays antitumor activity through apoptosis. Carbohydr. Polym. 2015, 124, 90–97. [Google Scholar] [CrossRef]
- Lee, M.S.; Hwang, B.S.; Lee, I.K.; Seo, G.S.; Yun, B.S. Chemical constituents of the culture broth of Phellinus linteus and their antioxidant activity. Mycobiology 2015, 43, 43–48. [Google Scholar] [CrossRef]
- Wang, Z.B.; Pei, J.J.; Ma, H.L.; Cai, P.F.; Yan, J.K. Effect of extraction media on preliminary characterizations and antioxidant activities of Phellinus linteus polysaccharides. Carbohydr. Polym. 2014, 109, 49–55. [Google Scholar] [CrossRef]
- Lee, S.; Lee, D.; Jang, T.S.; Kang, K.S.; Nam, J.W.; Lee, H.J.; Kim, K.H. Anti-inflammatory phenolic metabolites from the edible fungus Phellinus baumii in LPS-stimulated RAW264.7 cells. Molecules 2017, 22, 1583. [Google Scholar] [CrossRef]
- Park, H.J. Phellinus linteus grown on germinated brown rice suppress metastasis and induce apoptosis of colon cancer cells by suppressing NF-kappa B and Wnt/beta-catenin signaling pathways. J. Funct. Food. 2015, 14, 289–298. [Google Scholar] [CrossRef]
- Hu, T.; Lin, Q.L.; Guo, T.; Yang, T.; Zhou, W.H.; Deng, X.F.; Yan, J.K.; Luo, Y.; Ju, M.M.; Luo, F.J. Polysaccharide isolated from Phellinus linteus mycelia exerts anti-inflammatory effects via MAPK and PPAR signaling pathways. Carbohydr. Polym. 2018, 200, 487–497. [Google Scholar] [CrossRef]
- Wang, H.L.; Wu, G.; Park, H.J.; Jiang, P.P.; Sit, W.H.; van Griensven, L.; Wan, J.M.F. Protective effect of Phellinus linteus polysaccharide extracts against thioacetamide-induced liver fibrosis in rats: A proteomics analysis. Chin. Med. 2012, 7, 23. [Google Scholar] [CrossRef]
- Siriwattanametanon, W.; Kanchanarach, W.; Thiwthong, R.; Dodgson, J.L.A. Culture filtrates from laboratory grown Phellinus mushrooms for use as antibacterial agents. Chiang Mai J. Sci. 2014, 41, 243–247. [Google Scholar]
- Kim, D.H.; Yang, B.K.; Jeong, S.C.; Park, J.B.; Cho, S.P.; Das, S.; Yun, J.W.; Song, C.H. Production of a hypoglycemic, extracellular polysaccharide from the submerged culture of the mushroom, Phellinus linteus. Biotechnol. Lett. 2001, 23, 513–517. [Google Scholar] [CrossRef]
- Kim, H.M.; Kang, J.S.; Kim, J.Y.; Park, S.K.; Kim, H.S.; Lee, Y.J.; Yun, J.; Hong, J.T.; Kim, Y.; Han, S.B. Evaluation of antidiabetic activity of polysaccharide isolated from Phellinus linteus in non-obese diabetic mouse. Int. Immunopharmacol. 2010, 10, 72–78. [Google Scholar] [CrossRef]
- Yamac, M.; Zeytinoglu, M.; Senturk, H.; Kartkaya, K.; Kanbak, G.; Bayramoglu, G.; Oglakci, A.; Van Griensven, L. Effects of black hoof medicinal mushroom, Phellinus linteus (agaricomycetes), polysaccharide extract in streptozotocin-induced diabetic rats. Int. J. Med. Mushrooms 2016, 18, 301–311. [Google Scholar] [CrossRef]
- Feng, H.; Zhang, S.J.; Wan, J.M.F.; Gui, L.F.; Ruan, M.C.; Li, N.; Zhang, H.Y.; Liu, Z.G.; Wang, H.L. Polysaccharides extracted from Phellinus linteus ameliorate high-fat high-fructose diet induced insulin resistance in mice. Carbohydr. Polym. 2018, 200, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.S.; Lee, J.S.; Lee, J.H.; Kwon, D.S.; Lee, K.E.; Lee, S.Y.; Hong, E.K. Hispidin produced from Phellinus linteus protects pancreatic beta-cells from damage by hydrogen peroxide. Arch. Pharm. Res. 2010, 33, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Kang, I.J.; Won, M.H.; Lee, J.Y.; Kim, J.K.; Lim, S.S. Inhibition of protein tyrosine phosphatase 1 beta by hispidin derivatives isolated from the fruiting body of Phellinus linteus. Nat. Prod. Commun. 2010, 5, 1927–1930. [Google Scholar] [PubMed]
- Lee, J.S.; Kim, Y.R.; Jung, W.C.; Lee, K.E.; Lee, S.Y.; Hong, E.K. Hispidin isolated from Phellinus linteus protects against hydrogen peroxide-induced oxidative stress in pancreatic min6n beta-cells. J. Med. Food 2011, 14, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Jia, Z.; Tang, M.C.; Wu, J.M. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar]
- Wang, F.; Zhou, Z.K.; Ren, X.C.; Wang, Y.Y.; Yang, R.; Luo, J.H.; Strappe, P. Effect of Ganoderma lucidum spores intervention on glucose and lipid metabolism gene expression profiles in type 2 diabetic rats. Lipids Health Dis. 2015, 14, 49. [Google Scholar] [CrossRef]
- Srinivasan, K.; Viswanad, B.; Asrat, L.; Kaul, C.L.; Ramarao, P. Combination of high-fat diet-fed and low-dose streptozotocin-treated rat: A model for type 2 diabetes and pharmacological screening. Pharmacol. Res. 2005, 52, 313–320. [Google Scholar] [CrossRef]
- Hanley, A.J.G.; Williams, K.; Stern, M.P.; Haffner, S.M. Homeostasis model assessment of insulin resistance in relation to the incidence of cardiovascular disease—The san antonio heart study. Diabetes Care 2002, 25, 1177–1184. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Martins, S.; Mussatto, S.I.; Martinez-Avila, G.; Montanez-Saenz, J.; Aguilar, C.N.; Teixeira, J.A. Bioactive phenolic compounds: Production and extraction by solid-state fermentation. A review. Biotechnol. Adv. 2011, 29, 365–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Indumathi, D.; Sujithra, K.; Srinivasan, S.; Vinothkumar, V. Ameliorating effect of betanin, a natural chromoalkaloid by modulating hepatic carbohydrate metabolic enzyme activities and glycogen content in streptozotocin—Nicotinamide induced experimental rats. Biomed. Pharmacother. 2017, 88, 1069–1079. [Google Scholar]
- Teng, B.S.; Wang, C.D.; Zhang, D.; Wu, J.S.; Pan, D.; Pan, L.F.; Yang, H.J.; Zhou, P. Hypoglycemic effect and mechanism of a proteoglycan from Ganoderma lucidum on streptozotocin-induced type 2 diabetic rats. Eur. Rev. Med. Pharmacol. Sci. 2012, 16, 166–175. [Google Scholar] [PubMed]
- Subash Babu, P.; Prabuseenivasan, S.; Ignacimuthu, S. Cinnamaldehyde—A potential antidiabetic agent. Phytomed. Int. J. Phytother. Phytophar. 2007, 14, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Sattar, E.; El-Maraghy, S.A.; El-Dine, R.S.; Rizk, S.M. Russelioside b, a pregnane glycoside ameliorates hyperglycemia in streptozotocin induced diabetic rats by regulating key enzymes of glucose metabolism. Chem.-Biol. Interact. 2016, 252, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Poodineh, J.; Nakhaee, A. Hypoglycemic and hypolipidemic effects of Caralluma tuberculata and its safety on liver and kidneys of diabetic rats. Turk. J. Biochem. 2016, 41, 136–143. [Google Scholar]
- Postic, C.; Dentin, R.; Girard, J. Role of the liver in the control of carbohydrate and lipid homeostasis. Diabetes Metab. 2004, 30, 398–408. [Google Scholar] [CrossRef]
- Agius, L. New hepatic targets for glycaemic control in diabetes. Best Pract. Res. Clin. Endoc. Metab. 2007, 21, 587–605. [Google Scholar] [CrossRef]
- Trinh, K.Y.; O’Doherty, R.M.; Anderson, P.; Lange, A.J.; Newgard, C.B. Perturbation of fuel homeostasis caused by overexpression of the glucose-6-phosphatase catalytic subunit in liver of normal rats. J. Biol. Chem. 1998, 273, 31615–31620. [Google Scholar] [CrossRef]
- Brown, M.S.; Goldstein, J.L. Selective versus total insulin resistance: A pathogenic paradox. Cell Metab. 2008, 7, 95–96. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Wu, Q.P.; Cai, W.; Tan, J.B.; Yang, X.B.; Zhang, J.M. Hypoglycemic effects of Ganoderma lucidum polysaccharides in type 2 diabetic mice. Arch. Pharm. Res. 2012, 35, 1793–1801. [Google Scholar] [CrossRef] [PubMed]
- Seto, S.W.; Lam, T.Y.; Tam, H.L.; Au, A.L.S.; Chan, S.W.; Wu, J.H.; Yu, P.H.F.; Leung, G.P.H.; Ngai, S.M.; Yeung, J.H.K.; et al. Novel hypoglycemic effects of Ganoderma lucidum water-extract in obese/diabetic (+db/+db) mice. Phytomedicine 2009, 16, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.Q.; Ma, X.H.; Zhang, L.; Sun, H.; Liu, X. Capsaicin reduces blood glucose by increasing insulin levels and glycogen content better than capsiate in streptozotocin-induced diabetic rats. J. Agric. Food Chem. 2017, 65, 2323–2330. [Google Scholar] [CrossRef] [PubMed]
- Katsanos, C.S. Lipid-induced insulin resistance in the liver—Role of exercise. Sports Med. 2004, 34, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Lowell, B.B.; Shulmanz, G.I. Mitochondrial dysfunction and type 2 diabetes. Science 2005, 307, 384–387. [Google Scholar] [CrossRef]
- Zhu, L.L.; Zhu, X.Y.; Sun, G.B.; Meng, X.B.; Wang, M.; Cui, H.M.; Wang, J.L.; Zhai, Y.D.; Yang, K.; Tang, Y.; et al. Dai-zong-fang, a traditional chinese herbal formula, ameliorates insulin resistance in db/db mice. Front. Physiol. 2018, 9, 224. [Google Scholar] [CrossRef]
- Savage, D.B.; Choi, C.S.; Samuel, V.T.; Liu, Z.X.; Zhang, D.Y.; Wang, A.; Zhang, X.M.; Cline, G.W.; Yu, X.X.; Geisler, J.G.; et al. Reversal of diet-induced hepatic steatosis and hepatic insulin resistance by antisense oligonucleotide inhibitors of acetyl-coa carboxylases 1 and 2. J. Clin. Investig. 2006, 116, 817–824. [Google Scholar] [CrossRef]
- Niesen, M.; Bedi, M.; Lopez, D. Diabetes alters LDL receptor and PCSK9 expression in rat liver. Arch. Biochem. Biophys. 2008, 470, 111–115. [Google Scholar] [CrossRef]
- Jeong, S.Y.; Kang, S.; Kim, D.S.; Park, S. Codonopsis lanceolata water extract increases hepatic insulin sensitivity in rats with experimentally-induced type 2 diabetes. Nutrients 2017, 9, 1200. [Google Scholar] [CrossRef]
- Mbikay, M.; Mayne, J.; Sirois, F.; Fedoryak, O.; Raymond, A.; Noad, J.; Chretien, M. Mice fed a high-cholesterol diet supplemented with quercetin-3-glucoside show attenuated hyperlipidemia and hyperinsulinemia associated with differential regulation of PCSK9 and LDLR in their liver and pancreas. Mol. Nutr. Food Res. 2018, 62, 1700729. [Google Scholar] [CrossRef]
- Dai, J.Y.; Liang, K.; Zhao, S.; Jia, W.T.; Liu, Y.; Wu, H.K.; Lv, J.; Cao, C.; Chen, T.; Zhuang, S.T.; et al. Chemoproteomics reveals baicalin activates hepatic CPT1 to ameliorate diet-induced obesity and hepatic steatosis. Proc. Natl. Acad. Sci. USA 2018, 115, E5896–E5905. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.M.; Ma, J.Q.; Sun, J.M.; Cheng, C.; Feng, Z.J.; Jiang, H.; Yang, W. Flavonoid-rich extract of Paulownia fortunei flowers attenuates diet-induced hyperlipidemia, hepatic steatosis and insulin resistance in obesity mice by ampk pathway. Nutrients 2017, 9, 959. [Google Scholar] [CrossRef]
- Frances, D.E.; Ronco, M.T.; Monti, J.A.; Ingaramo, P.I.; Pisani, G.B.; Parody, J.P.; Pellegrino, J.M.; Sanz, P.M.; Carrillo, M.C.; Carnovale, C.E. Hyperglycemia induces apoptosis in rat liver through the increase of hydroxyl radical: New insights into the insulin effect. J. Endocrinol. 2010, 205, 187–200. [Google Scholar] [CrossRef]
- Hwang, H.J.; Kim, S.W.; Lim, J.M.; Joo, J.H.; Kim, H.O.; Kim, H.M.; Yun, J.W. Hypoglycemic effect of crude exopolysaccharides produced by a medicinal mushroom Phellinus baumii in streptozotocin-induced diabetic rats. Life Sci. 2005, 76, 3069–3080. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Chen, X.X.; Tang, S.C.W.; Lao, L.X.; Sze, S.C.W.; Lee, K.F.; Zhang, K.Y.B. Edible plants from traditional chinese medicine is a promising alternative for the management of diabetic nephropathy. J. Funct. Food. 2015, 14, 12–22. [Google Scholar] [CrossRef]
- Gao, C.P.; Zhong, L.F.; Jiang, L.P.; Geng, C.Y.; Yao, X.F.; Cao, J. Phellinus linteus mushroom protects against tacrine-induced mitochondrial impairment and oxidative stress in HepG2 cells. Phytomedicine 2013, 20, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.J.; Huang, S.S.; Deng, J.S. Anti-inflammatory activities of inotilone from Phellinus linteus through the inhibition of MMP-9, NF-kappa B, and MAPK activation in vitro and in vivo. PLoS ONE 2012, 7, e35922. [Google Scholar]
- Lee, Y.S.; Kang, Y.H.; Jung, J.Y.; Lee, S.; Ohuchi, K.; Shin, K.H.; Kang, I.J.; Park, J.H.Y.; Shin, H.K.; Lim, S.S. Protein glycation inhibitors from the fruiting body of Phellinus linteus. Biol. Pharm. Bull. 2008, 31, 1968–1972. [Google Scholar] [CrossRef]
- Jayachandran, M.; Xiao, J.B.; Xu, B.J. A critical review on health promoting benefits of edible mushrooms through gut microbiota. Int. J. Mol. Sci. 2017, 18, 1934. [Google Scholar] [CrossRef]
- Sonnenburg, E.D.; Sonnenburg, J.L. Starving our microbial self: The deleterious consequences of a diet deficient in microbiota-accessible carbohydrates. Cell Metab. 2014, 20, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Neyrinck, A.M.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.-R.; Lin, C.-S.; Chang, C.-J.; Lin, T.-L.; Martel, J.; Ko, Y.-F.; Ojcius, D.M.; Lu, C.-C.; Young, J.D.; Lai, H.-C. Gut commensal Parabacteroides goldsteinii plays a predominant role in the anti-obesity effects of polysaccharides isolated from Hirsutella sinensis. Gut 2018, 68, 248–262. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Content (%) | Detection Method |
|---|---|---|
| Polysaccharides | 62.56% | Phenol-sulfuric acid method |
| Proteins | 28.34% | Lowry method |
| Flavonoids | 3.87% | Aluminum chloride spectrophotometric method |
| Parameter | Group | ||||
|---|---|---|---|---|---|
| NC | Model | Met | L | H | |
| AST (IU/L) | 14.99 ± 1.27 bb | 46.27 ± 6.39 aa | 23.55 ± 2.31 a,bb | 19.76 ± 1.68 bb | 17.13±1.87 bb |
| ALT (IU/L) | 27.51 ± 4.58 b | 123.17 ± 16.61 a | 43.86 ± 4.40 b | 43.32 ± 6.53 b | 36.55 ± 2.68 b |
| AKP (U/L) | 27.93 ± 1.99 bb | 102.12 ± 6.37 aa | 64.24 ± 6.76 aa,bb | 58.18 ± 6.10 a,bb | 62.34 ± 6.08 aa,bb |
| Bil (μmol/L) | 1.80 ± 0.19 b | 6.13 ± 0.89 a | 5.91 ± 1.06 | 5.41 ± 0.89 | 4.96 ± 0.93 |
| UA (mol/L) | 111.43 ± 1.52 b | 164.08 ± 8.97 a | 134.63 ± 10.67 | 116.60 ± 7.69 b | 110.66 ± 5.80 bb |
| Cre (μmol/L) | 45.79 ± 3.55 bb | 71.46 ± 2.86 aa | 60.06 ± 5.76 | 53.61 ± 3.18 b | 49.46 ± 2.59 bb |
| BUN (mmol/L) | 2.21 ± 0.15 bb | 7.15 ± 0.58 aa | 6.48 ± 0.46 aa | 6.08 ± 0.30 aa | 4.86 ± 0.12 aa |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Wang, C.; Li, J.; Mei, Y.; Liang, Y. Hypoglycemic and Hypolipidemic Effects of Phellinus Linteus Mycelial Extract from Solid-State Culture in A Rat Model of Type 2 Diabetes. Nutrients 2019, 11, 296. https://doi.org/10.3390/nu11020296
Liu Y, Wang C, Li J, Mei Y, Liang Y. Hypoglycemic and Hypolipidemic Effects of Phellinus Linteus Mycelial Extract from Solid-State Culture in A Rat Model of Type 2 Diabetes. Nutrients. 2019; 11(2):296. https://doi.org/10.3390/nu11020296
Chicago/Turabian StyleLiu, Yangyang, Chaorui Wang, Jinshan Li, Yuxia Mei, and Yunxiang Liang. 2019. "Hypoglycemic and Hypolipidemic Effects of Phellinus Linteus Mycelial Extract from Solid-State Culture in A Rat Model of Type 2 Diabetes" Nutrients 11, no. 2: 296. https://doi.org/10.3390/nu11020296
APA StyleLiu, Y., Wang, C., Li, J., Mei, Y., & Liang, Y. (2019). Hypoglycemic and Hypolipidemic Effects of Phellinus Linteus Mycelial Extract from Solid-State Culture in A Rat Model of Type 2 Diabetes. Nutrients, 11(2), 296. https://doi.org/10.3390/nu11020296