Immunomodulatory Effects of Glutathione, Garlic Derivatives, and Hydrogen Sulfide



Abstract
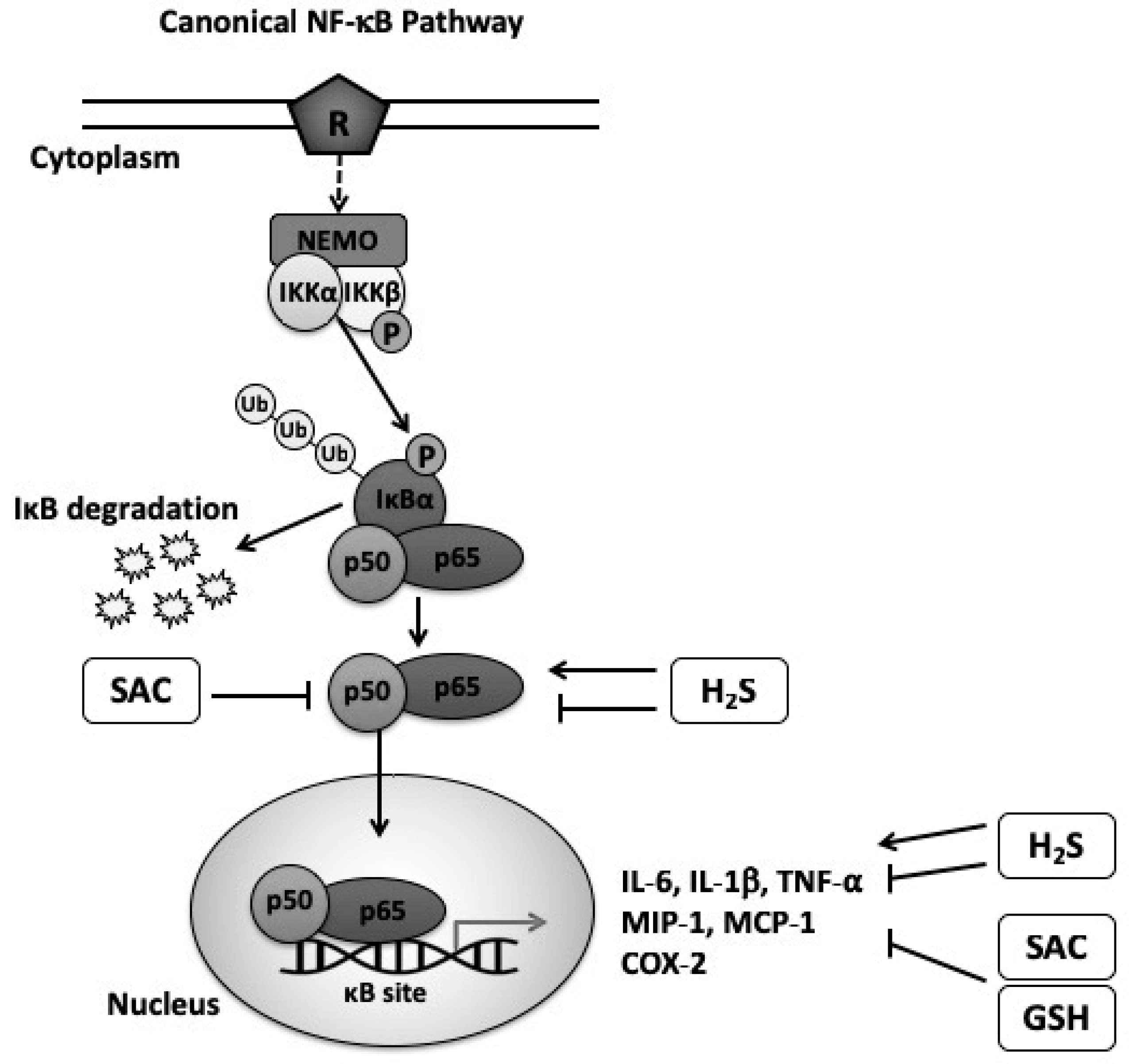
:1. Introduction
2. Glutathione
3. Garlic (Allium sativum L.)
Aged Garlic Extract (AGE)
4. Hydrogen Sulfide
5. Current Status of Knowledge
5.1. Glutathione and Immunity
5.2. Fresh Garlic Derivatives and Immunity
5.3. AGE and Immunity
5.4. Hydrogen Sulfide and Inflammation
6. Relationship between GSH, Garlic Derivatives and H2S
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Droge, W.; Breitkreutz, R. Glutathione and immune function. Proc. Nutr. Soc. 2000, 59, 595–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, T.; Meydani, S.N.; Blumberg, J.B. Reversal of age-associated decline in immune responsiveness by dietary glutathione supplementation in mice. Mech. Ageing Dev. 1987, 38, 107–117. [Google Scholar] [CrossRef]
- Geng, Z.; Rong, Y.; Lau, B.H. S-allyl cysteine inhibits activation of nuclear factor kappa B in human T cells. Free Radic. Biol. Med. 1997, 23, 345–350. [Google Scholar] [CrossRef]
- Ahmadi, N.; Nebavi, V.; Hajsadeghi, F.; Zeb, I.; Flores, F.; Ebrahimi, R.; Budoff, M. Aged garlic extract with supplement is associated with increase in brown adipose, decrease in white adipose tissue and predict lack of progression in coronary atherosclerosis. Int. J. Cardiol. 2013, 168, 2310–2314. [Google Scholar] [CrossRef] [PubMed]
- Mong, M.C.; Yin, M.C. Nuclear factor kappaB-dependent anti-inflammatory effects of s-allyl cysteine and s-propyl cysteine in kidney of diabetic mice. J. Agric. Food Chem. 2012, 60, 3158–3165. [Google Scholar] [CrossRef] [PubMed]
- Baluchnejadmojarad, T.; Kiasalari, Z.; Afshin-Majd, S.; Ghasemi, Z.; Roghani, M. S-allyl cysteine ameliorates cognitive deficits in streptozotocin-diabetic rats via suppression of oxidative stress, inflammation, and acetylcholinesterase. Eur. J. Pharmacol. 2017, 794, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Warenycia, M.W.; Goodwin, L.R.; Benishin, C.G.; Reiffenstein, R.J.; Francom, D.M.; Taylor, J.D.; Dieken, F.P. Acute hydrogen sulfide poisoning. Demonstration of selective uptake of sulfide by the brainstem by measurement of brain sulfide levels. Biochem. Pharmacol. 1989, 38, 973–981. [Google Scholar] [CrossRef]
- Stipanuk, M.H.; Dominy, J.E., Jr.; Lee, J.I.; Coloso, R.M. Mammalian cysteine metabolism: New insights into regulation of cysteine metabolism. J. Nutr. 2006, 136, 1652s–1659s. [Google Scholar] [CrossRef]
- Yang, G.; Wu, L.; Jiang, B.; Yang, W.; Qi, J.; Cao, K.; Meng, Q.; Mustafa, A.K.; Mu, W.; Zhang, S.; et al. H2S as a physiologic vasorelaxant: Hypertension in mice with deletion of cystathionine gamma-lyase. Science 2008, 322, 587–590. [Google Scholar] [CrossRef]
- Whiteman, M.; Gooding, K.M.; Whatmore, J.L.; Ball, C.I.; Mawson, D.; Skinner, K.; Tooke, J.E.; Shore, A.C. Adiposity is a major determinant of plasma levels of the novel vasodilator hydrogen sulphide. Diabetologia 2010, 53, 1722–1726. [Google Scholar] [CrossRef]
- Kennedy, M.A. A brief review of the basics of immunology: The innate and adaptive response. Vet. Clin. N. Am. Small Anim. Pract. 2010, 40, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Candela, J.; Velmurugan, G.V.; White, C. Hydrogen sulfide depletion contributes to microvascular remodeling in obesity. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H1071–H1080. [Google Scholar] [CrossRef] [PubMed]
- Zhi, L.; Ang, A.D.; Zhang, H.; Moore, P.K.; Bhatia, M. Hydrogen sulfide induces the synthesis of proinflammatory cytokines in human monocyte cell line U937 via the ERK-NF-kappaB pathway. J. Leukoc. Biol. 2007, 81, 1322–1332. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.W.; Feng, W.; Peh, M.T.; Peh, K.; Dymock, B.W.; Moore, P.K. A novel slow-releasing hydrogen sulfide donor, FW1256, exerts anti-inflammatory effects in mouse macrophages and in vivo. Pharmacol. Res. 2016, 113, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Meister, A. Glutathione, ascorbate, and cellular protection. Cancer Res. 1994, 54, 1969s–1975s. [Google Scholar] [PubMed]
- Jones, D.P.; Coates, R.J.; Flagg, E.W.; Eley, J.W.; Block, G.; Greenberg, R.S.; Gunter, E.W.; Jackson, B. Glutathione in foods listed in the National Cancer Institute’s Health Habits and History Food Frequency Questionnaire. Nutr. Cancer 1992, 17, 57–75. [Google Scholar] [CrossRef]
- He, M.; Openo, K.; Cullough, M.M.; Jones, D.P. Total equivalent of reactive chemicals in 142 human food items is highly variable within and between major food groups. J. Nutr. 2004, 134, 1114–1119. [Google Scholar] [CrossRef]
- Witschi, A.; Reddy, S.; Stofer, B.; Lauterburg, B.H. The systemic availability of oral glutathione. Eur. J. Clin. Pharmacol. 1992, 43, 667–669. [Google Scholar] [CrossRef]
- Meister, A.; Anderson, M.E. Glutathione. Annu. Rev. Biochem. 1983, 52, 711–760. [Google Scholar] [CrossRef]
- Zhao, W.; Mosley, B.S.; Cleves, M.A.; Melnyk, S.; James, S.J.; Hobbs, C.A. Neural tube defects and maternal biomarkers of folate, homocysteine, and glutathione metabolism. Birth Defects Res. A Clin. Mol. Teratol. 2006, 76, 230–236. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.C. Glutathione synthesis. Biochim. Biophys. Acta 2013, 1830, 3143–3153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.L.; Hammarqvist, F.; Andersson, K.; Wernerman, J. Surgical trauma decreases glutathione synthetic capacity in human skeletal muscle tissue. Am. J. Physiol. 1998, 275, E359–E365. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.A.; Yang, H.; Chen, C.; Zeng, Z.; Lu, S.C. Inducers of gamma-glutamylcysteine synthetase and their effects on glutathione synthetase expression. Biochim. Biophys. Acta 2000, 1493, 48–55. [Google Scholar] [CrossRef]
- Yang, H.; Zeng, Y.; Lee, T.D.; Yang, Y.; Ou, X.; Chen, L.; Haque, M.; Rippe, R.; Lu, S.C. Role of AP-1 in the coordinate induction of rat glutamate-cysteine ligase and glutathione synthetase by tert-butylhydroquinone. J. Biol. Chem. 2002, 277, 35232–35239. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, D.M. Structural, functional, and clinical aspects of gamma-glutamyltransferase. CRC Crit. Rev. Clin. Lab. Sci. 1980, 12, 1–58. [Google Scholar] [CrossRef] [PubMed]
- Orlowski, M.; Meister, A. The gamma-glutamyl cycle: A possible transport system for amino acids. Proc. Natl. Acad. Sci. USA 1970, 67, 1248–1255. [Google Scholar] [CrossRef] [PubMed]
- Aoyama, K.; Nakaki, T. Inhibition of GTRAP3-18 may increase neuroprotective glutathione (GSH) synthesis. Int. J. Mol. Sci. 2012, 13, 12017–12035. [Google Scholar] [CrossRef]
- Griffith, O.W. Biologic and pharmacologic regulation of mammalian glutathione synthesis. Free Radic. Biol. Med. 1999, 27, 922–935. [Google Scholar] [CrossRef]
- Lu, S.C. Regulation of glutathione synthesis. Curr. Top. Cell. Regul. 2000, 36, 95–116. [Google Scholar]
- Martensson, J. The effect of fasting on leukocyte and plasma glutathione and sulfur amino acid concentrations. Metabolism 1986, 35, 118–121. [Google Scholar] [CrossRef]
- Calvert, J.W.; Jha, S.; Gundewar, S.; Elrod, J.W.; Ramachandran, A.; Pattillo, C.B.; Kevil, C.G.; Lefer, D.J. Hydrogen sulfide mediates cardioprotection through Nrf2 signaling. Circ. Res. 2009, 105, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Goto, Y.; Kimura, H. Hydrogen sulfide increases glutathione production and suppresses oxidative stress in mitochondria. Antioxid. Redox Signal. 2010, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Whiteman, M.; Cheung, N.S.; Zhu, Y.Z.; Chu, S.H.; Siau, J.L.; Wong, B.S.; Armstrong, J.S.; Moore, P.K. Hydrogen sulphide: A novel inhibitor of hypochlorous acid-mediated oxidative damage in the brain? Biochem. Biophys. Res. Commun. 2005, 326, 794–798. [Google Scholar] [CrossRef] [PubMed]
- Blanco, R.A.; Ziegler, T.R.; Carlson, B.A.; Cheng, P.Y.; Park, Y.; Cotsonis, G.A.; Accardi, C.J.; Jones, D.P. Diurnal variation in glutathione and cysteine redox states in human plasma. Am. J. Clin. Nutr. 2007, 86, 1016–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nimni, M.E.; Han, B.; Cordoba, F. Are we getting enough sulfur in our diet? Nutr. Metab. 2007, 4, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.F.; Chen, L.T.; Boyce, H.W., Jr. Cruciferous vegetables and glutathione: Their effects on colon mucosal glutathione level and colon tumor development in rats induced by DMH. Nutr. Cancer 1995, 23, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Tarozzi, A.; Morroni, F.; Merlicco, A.; Hrelia, S.; Angeloni, C.; Cantelli-Forti, G.; Hrelia, P. Sulforaphane as an inducer of glutathione prevents oxidative stress-induced cell death in a dopaminergic-like neuroblastoma cell line. J. Neurochem. 2009, 111, 1161–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, F.; Morisco, F.; Verde, V.; Ritieni, A.; Alezio, A.; Caporaso, N.; Fogliano, V. Moderate coffee consumption increases plasma glutathione but not homocysteine in healthy subjects. Aliment Pharmacol. Ther. 2003, 17, 595–601. [Google Scholar] [CrossRef] [Green Version]
- Metro, D.; Cernaro, V.; Santoro, D.; Papa, M.; Buemi, M.; Benvenga, S.; Manasseri, L. Beneficial effects of oral pure caffeine on oxidative stress. J. Clin. Transl. Endocrinol. 2017, 10, 22–27. [Google Scholar] [CrossRef]
- Butt, M.S.; Sultan, M.T.; Iqbal, J. Garlic: Nature’s protection against physiological threats. Crit. Rev. Food Sci. Nutr. 2009, 49, 538–551. [Google Scholar] [CrossRef]
- Lawson, L.D.; Wood, S.G.; Hughes, B.G. HPLC analysis of allicin and other thiosulfinates in garlic clove homogenates. Planta Med. 1991, 57, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Amagase, H. Clarifying the real bioactive constituents of garlic. J. Nutr. 2006, 136, 716s–725s. [Google Scholar] [CrossRef] [PubMed]
- LaChance, P.A. (Ed.) Phytochemistry of garlic horticultural and processing procedures. In Neutraceuticals: Designer Foods III. Garlic, Soy and Licorice; Food and Nutrition Press: Trumbull, CT, USA, 1997. [Google Scholar]
- Chung, L.Y. The antioxidant properties of garlic compounds: Allyl cysteine, alliin, allicin, and allyl disulfide. J. Med. Food 2006, 9, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Manoj Kumar, V.; Henley, A.K.; Nelson, C.J.; Indumati, O.; Prabhakara Rao, Y.; Rajanna, S.; Rajanna, B. Protective effect of Allium sativum (garlic) aqueous extract against lead-induced oxidative stress in the rat brain, liver, and kidney. Environ. Sci. Pollut. Res. Int. 2017, 24, 1544–1552. [Google Scholar] [CrossRef] [PubMed]
- Panyod, S.; Wu, S.; Ho, C.T.; Lu, K.H.; Liu, C.T.; Chu, Y.L.; Lai, Y.S.; Chen, W.C.; Lin, Y.E.; Lin, S.H.; et al. Diet Supplementation with Allicin Protects against Alcoholic Fatty Liver Disease in Mice by Improving Anti-inflammation and Antioxidative Functions. J. Agric. Food Chem. 2016, 64, 7104–7113. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.S.; Kim, J.Y.; Paek, J.E.; Lee, Y.J.; Kim, H.R.; Park, D.S.; Kwon, O. Garlic powder intake and cardiovascular risk factors: A meta-analysis of randomized controlled clinical trials. Nutr. Res. Pract. 2014, 8, 644–654. [Google Scholar] [CrossRef] [PubMed]
- Moutia, M.; Seghrouchni, F.; Abouelazz, O.; Elouaddari, A.; Al Jahid, A.; Elhou, A.; Nadifi, S.; Jamal Eddine, J.; Habti, N.; Badou, A. Allium sativum L. regulates in vitro IL-17 gene expression in human peripheral blood mononuclear cells. BMC Complement. Altern. Med. 2016, 16, 377. [Google Scholar] [CrossRef] [PubMed]
- Fratianni, F.; Riccardi, R.; Spigno, P.; Ombra, M.N.; Cozzolino, A.; Tremonte, P.; Coppola, R.; Nazzaro, F. Biochemical Characterization and Antimicrobial and Antifungal Activity of Two Endemic Varieties of Garlic (Allium sativum L.) of the Campania Region, Southern Italy. J. Med. Food 2016, 19, 686–691. [Google Scholar] [CrossRef] [PubMed]
- Amagase, H.; Petesch, B.L.; Matsuura, H.; Kasuga, S.; Itakura, Y. Intake of garlic and its bioactive components. J. Nutr. 2001, 131, 955s–962s. [Google Scholar] [CrossRef]
- Nagae, S.; Ushijima, M.; Hatono, S.; Imai, J.; Kasuga, S.; Matsuura, H.; Itakura, Y.; Higashi, Y. Pharmacokinetics of the garlic compound S-allylcysteine. Planta Med. 1994, 60, 214–217. [Google Scholar] [CrossRef]
- Kodera, Y.; Suzuki, A.; Imada, O.; Kasuga, S.; Sumioka, I.; Kanezawa, A.; Taru, N.; Fujikawa, M.; Nagae, S.; Masamoto, K.; et al. Physical, chemical, and biological properties of s-allylcysteine, an amino acid derived from garlic. J. Agric. Food Chem. 2002, 50, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Elosta, A.; Slevin, M.; Rahman, K.; Ahmed, N. Aged garlic has more potent antiglycation and antioxidant properties compared to fresh garlic extract in vitro. Sci. Rep. 2017, 7, 39613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, J.; Ide, N.; Nagae, S.; Moriguchi, T.; Matsuura, H.; Itakura, Y. Antioxidant and radical scavenging effects of aged garlic extract and its constituents. Planta Med. 1994, 60, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Morihara, N.; Hayama, M.; Fujii, H. Aged garlic extract scavenges superoxide radicals. Plant Foods Hum. Nutr. 2011, 66, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Kohda, K.; Goda, H.; Itoh, K.; Samejima, K.; Fukuuchi, T. Aged Garlic Extract Reduces ROS Production and Cell Death Induced by 6-Hydroxydopamine through Activation of the Nrf2-ARE Pathway in SH-SY5Y Cells. Pharmacol. Pharm. 2013, 4, 31–40. [Google Scholar] [CrossRef]
- Borek, C. Antioxidant health effects of aged garlic extract. J. Nutr. 2001, 131, 1010s–1015s. [Google Scholar] [CrossRef]
- Perez-Torres, I.; Torres-Narváez, J.C.; Pedraza-Chaverri, J.; Rubio-Ruiz, M.; Díaz-Díaz, E.; Del Valle-Mondragón, L.; Martínez-Memije, R.; Varela López, E.; Guarner-Lans, V. Effect of the Aged Garlic Extract on Cardiovascular Function in Metabolic Syndrome Rats. Molecules 2016, 21, 1425. [Google Scholar] [CrossRef]
- Hiramatsu, K.; Tsuneyoshi, T.; Ogawa, T.; Morihara, N. Aged garlic extract enhances heme oxygenase-1 and glutamate-cysteine ligase modifier subunit expression via the nuclear factor erythroid 2-related factor 2-antioxidant response element signaling pathway in human endothelial cells. Nutr. Res. 2016, 36, 143–149. [Google Scholar] [CrossRef]
- Colín-González, A.L. The Antioxidant Mechanisms Underlying the Aged Garlic Extract- and S-Allylcysteine-Induced Protection. Oxid. Med. Cell. Longev. 2012, 2012, 907162. [Google Scholar] [CrossRef]
- Ried, K.; Frank, O.R.; Stocks, N.P. Aged garlic extract reduces blood pressure in hypertensives: A dose-response trial. Eur. J. Clin. Nutr. 2013, 67, 64–70. [Google Scholar] [CrossRef]
- Ebrahimi, T.; Behdad, B.; Abbasi, M.A.; Rabati, R.G.; Fayyaz, A.F.; Behnod, V.; Asgari, A. High doses of garlic extract significantly attenuated the ratio of serum LDL to HDL level in rat-fed with hypercholesterolemia diet. Diagn. Pathol. 2015, 10, 74. [Google Scholar] [CrossRef] [PubMed]
- Thomson, M.; Al-Katar, K.K.; Divya, J.S.; Ali, M. Anti-diabetic and anti-oxidant potential of aged garlic extract (AGE) in streptozotocin-induced diabetic rats. BMC Complement. Altern. Med. 2015, 16, 17. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Saeki, T.; Otani, T.; Suzuki, T.; Shimozuma, K.; Nishino, N.; Fukuda, S.; Morimoto, K. Aged garlic extract prevents a decline of NK cell number and activity in patients with advanced cancer. J. Nutr. 2006, 136, 816s–820s. [Google Scholar] [CrossRef] [PubMed]
- Orozco-Ibarra, M.; Muñoz-Sánchez, J.; Zavala-Medina, M.E.; Pineda, B.; Magaña-Maldonado, R.; Vázquez-Contreras, E.; Maldonado, P.D.; Pedraza-Chaverri, J.; Chánez-Cárdenas, M.E. Aged garlic extract and S-allylcysteine prevent apoptotic cell death in a chemical hypoxia model. Biol. Res. 2016, 49, 7. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, N.B.; Sandoval, J. Amelioration of early cognitive deficits by aged garlic extract in Alzheimer’s transgenic mice. Phytother. Res. 2007, 21, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Olson, K.R.; Straub, K.D. The Role of Hydrogen Sulfide in Evolution and the Evolution of Hydrogen Sulfide in Metabolism and Signaling. Physiology 2016, 31, 60–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tvedt, B.; Skyberg, K.; Aaserud, O.; Hobbesland, A.; Mathiesen, T. Brain damage caused by hydrogen sulfide: A follow-up study of six patients. Am. J. Ind. Med. 1991, 20, 91–101. [Google Scholar] [CrossRef]
- Wang, R. Hydrogen sulfide: The third gasotransmitter in biology and medicine. Antioxid. Redox Signal. 2010, 12, 1061–1064. [Google Scholar] [CrossRef]
- Stein, A.; Bailey, S.M. Redox Biology of Hydrogen Sulfide: Implications for Physiology, Pathophysiology, and Pharmacology. Redox Biol. 2013, 1, 32–39. [Google Scholar] [CrossRef]
- Kim, D.; Anantharam, P.V.; Whitley, E.M.; Gilbreath, E.; Mahama, B.Y.; Kanthasamy, A.; Rumbeiha, W.K. Acute Exposure to High Level of Hydrogen Sulfide Induces Neurodegeneration via Activation of Inflammasomes in Brain. FASEB J. 2016, 30, 1195.1. [Google Scholar]
- Lu, S.C.; Mato, J.M. S-adenosylmethionine in liver health, injury, and cancer. Physiol. Rev. 2012, 92, 1515–1542. [Google Scholar] [CrossRef] [PubMed]
- Stipanuk, M.H. Sulfur amino acid metabolism: Pathways for production and removal of homocysteine and cysteine. Annu. Rev. Nutr. 2004, 24, 539–577. [Google Scholar] [CrossRef] [PubMed]
- DeRatt, B.; Ralat, M.; Gregory, J. Characterization of Cystathionine Beta-Synthase and Cystathionine Gamma-Lyase in the Production of Hydrogen Sulfide Biomarkers, Lanthionine and Homolanthionine, in a HepG2 Cell Culture Model. FASEB J. 2016, 30, 1171. [Google Scholar]
- Shibuya, N.; Tanaka, M.; Yoshida, M.; Ogasawara, Y.; Togawa, T.; Ishii, K.; Kimura, H. 3-Mercaptopyruvate sulfurtransferase produces hydrogen sulfide and bound sulfane sulfur in the brain. Antioxid. Redox Signal. 2009, 11, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Kabil, O.; Banerjee, R. Redox biochemistry of hydrogen sulfide. J. Biol. Chem. 2010, 285, 21903–21907. [Google Scholar] [CrossRef] [PubMed]
- Wang, R. Two’s company, three’s a crowd: Can H2S be the third endogenous gaseous transmitter? FASEB J. 2002, 16, 1792–1798. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Gehlot, P. Inflammation and cancer: How friendly is the relationship for cancer patients? Curr. Opin. Pharmacol. 2009, 9, 351–369. [Google Scholar] [CrossRef]
- Predmore, B.L.; Lefer, D.J.; Gojon, G. Hydrogen sulfide in biochemistry and medicine. Antioxid. Redox Signal. 2012, 17, 119–140. [Google Scholar] [CrossRef]
- Tang, G.; Wu, L.; Wang, R. Interaction of hydrogen sulfide with ion channels. Clin. Exp. Pharmacol. Physiol. 2010, 37, 753–763. [Google Scholar] [CrossRef]
- Saha, S.; Chakraborty, P.K.; Xiong, X.; Dwivedi, S.K.; Mustafi, S.B.; Leigh, N.R.; Ramchandran, R.; Mukherjee, P.; Bhattacharya, R. Cystathionine beta-synthase regulates endothelial function via protein S-sulfhydration. FASEB J. 2016, 30, 441–456. [Google Scholar] [CrossRef]
- Zhang, Z.; Huang, H.; Liu, P.; Tang, C.; Wang, J. Hydrogen sulfide contributes to cardioprotection during ischemia-reperfusion injury by opening K ATP channels. Can. J. Physiol. Pharmacol. 2007, 85, 1248–1253. [Google Scholar] [CrossRef] [PubMed]
- Hourihan, J.M.; Kenna, J.G.; Hayes, J.D. The gasotransmitter hydrogen sulfide induces nrf2-target genes by inactivating the keap1 ubiquitin ligase substrate adaptor through formation of a disulfide bond between cys-226 and cys-613. Antioxid. Redox Signal. 2013, 19, 465–481. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Zhao, K.; Ju, Y.; Mani, S.; Cao, Q.; Puukila, S.; Khaper, N.; Wu, L.; Wang, R. Hydrogen sulfide protects against cellular senescence via S-sulfhydration of Keap1 and activation of Nrf2. Antioxid. Redox Signal. 2013, 18, 1906–1919. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Gu, Y.; Wen, M.; Zhao, S.; Wang, W.; Ma, Y.; Meng, G.; Han, Y.; Wang, Y.; Liu, G.; et al. Hydrogen Sulfide Induces Keap1 S-sulfhydration and Suppresses Diabetes-Accelerated Atherosclerosis via Nrf2 Activation. Diabetes 2016, 65, 3171–3184. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, A.K.; Gadalla, M.M.; Sen, N.; Kim, S.; Mu, W.; Gazi, S.K.; Barrow, R.K.; Yang, G.; Wang, R.; Snyder, S.H. H2S signals through protein S-sulfhydration. Sci. Signal. 2009, 2, ra72. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Wang, R. H(2)S-induced vasorelaxation and underlying cellular and molecular mechanisms. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H474–H480. [Google Scholar] [CrossRef]
- Fiorucci, S.; Antonelli, E.; Distrutti, E.; Rizzo, G.; Mencarelli, A.; Orlandi, S.; Zanardo, R.; Renga, B.; Di Sante, M.; Morelli, A.; et al. Inhibition of hydrogen sulfide generation contributes to gastric injury caused by anti-inflammatory nonsteroidal drugs. Gastroenterology 2005, 129, 1210–1224. [Google Scholar] [CrossRef]
- Wallace, J.L.; Dicay, M.; McKnight, W.; Martin, G.R. Hydrogen sulfide enhances ulcer healing in rats. FASEB J. 2007, 21, 4070–4076. [Google Scholar] [CrossRef]
- Benavides, G.A.; Squadrito, G.L.; Mills, R.W.; Patel, H.D.; Isbell, T.S.; Patel, R.P.; Darley-Usmar, V.M.; Doeller, J.E.; Kraus, D.W. Hydrogen sulfide mediates the vasoactivity of garlic. Proc. Natl. Acad. Sci. USA 2007, 104, 17977–17982. [Google Scholar] [CrossRef] [Green Version]
- Predmore, B.L.; Kondo, K.; Bhushan, S.; Zlatopolsky, M.A.; King, A.L.; Aragon, J.P.; Grinsfelder, D.B.; Condit, M.E.; Lefer, D.J. The polysulfide diallyl trisulfide protects the ischemic myocardium by preservation of endogenous hydrogen sulfide and increasing nitric oxide bioavailability. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H2410–H2418. [Google Scholar] [CrossRef] [Green Version]
- Hine, C.; Mitchell, J.R. Calorie restriction and methionine restriction in control of endogenous hydrogen sulfide production by the transsulfuration pathway. Exp. Gerontol. 2015, 68, 26–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.J.; Cai, G.Y.; Ning, Y.C.; Cui, J.; Hong, Q.; Bai, X.Y.; Xu, X.M.; Bu, R.; Sun, X.F.; Chen, X.M. Hydrogen sulfide mediates the protection of dietary restriction against renal senescence in aged F344 rats. Sci. Rep. 2016, 6, 30292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iantomasi, T.; Favilli, F.; Marraccini, P.; Magaldi, T.; Bruni, P.; Vincenzini, M.T. Glutathione transport system in human small intestine epithelial cells. Biochim. Biophys. Acta 1997, 1330, 274–283. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Fang, Y.Z.; Yang, S.; Lupton, J.R.; Turner, N.D. Glutathione metabolism and its implications for health. J. Nutr. 2004, 134, 489–492. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Garg, S.K.; Banerjee, R. Regulatory T cells interfere with glutathione metabolism in dendritic cells and T cells. J. Biol. Chem. 2010, 285, 41525–41532. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.M.; Albertini, D.F.; Berlin, R.D. Effects of glutathione-oxidizing agents on microtubule assembly and microtubule-dependent surface properties of human neutrophils. J. Cell Biol. 1976, 71, 921–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadzic, T.; Li, L.; Cheng, N.; Walsh, S.A.; Spitz, D.R.; Knudson, C.M. The role of low molecular weight thiols in T lymphocyte proliferation and IL-2 secretion. J. Immunol. 2005, 175, 7965–7972. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, X.; Fang, X.; Niimi, M.; Huang, Y.; Piao, H.; Gao, S.; Fan, J.; Yao, J. Glutathione inhibits antibody and complement-mediated immunologic cell injury via multiple mechanisms. Redox Biol. 2017, 12, 571–581. [Google Scholar] [CrossRef]
- De Rosa, S.C.; Zaretsky, M.D.; Dubs, J.G.; Roederer, M.; Anderson, M.; Green, A.; Miltra, D.; Watanabe, N.; Nakamura, H.; Tijoe, I.; et al. N-acetylcysteine replenishes glutathione in HIV infection. Eur. J. Clin. Invest. 2000, 30, 915–929. [Google Scholar] [CrossRef]
- Valdivia, A.; Ly, J.; Gonzalez, L.; Hussain, P.; Saing, T.; Islamoglu, H.; Pearce, D.; Ochoa, C.; Venketaraman, V. Restoring Cytokine Balance in HIV-Positive Individuals with Low CD4 T Cell Counts. AIDS Res. Hum. Retroviruses 2017, 33, 905–918. [Google Scholar] [CrossRef]
- Pena, L.R.; Hill, D.B.; McClain, C.J. Treatment with glutathione precursor decreases cytokine activity. JPEN J. Parenter. Enteral. Nutr. 1999, 23, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lou, H.; Kaplowitz, N. Glutathione depletion down-regulates tumor necrosis factor alpha-induced NF-kappaB activity via IkappaB kinase-dependent and -independent mechanisms. J. Biol. Chem. 2007, 282, 29470–29481. [Google Scholar] [CrossRef] [PubMed]
- Steele, M.L.; Fuller, S.; Patel, M.; Kersaitis, C.; Ooi, L.; Münch, G. Effect of Nrf2 activators on release of glutathione, cysteinylglycine and homocysteine by human U373 astroglial cells. Redox Biol. 2013, 1, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Jing, X.; Wei, X.; Perez, R.G.; Ren, M.; Zhang, X.; Lou, H. S-allyl cysteine activates the Nrf2-dependent antioxidant response and protects neurons against ischemic injury in vitro and in vivo. J. Neurochem. 2015, 133, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Harvey, C.J.; Thimmulappa, R.K.; Singh, A.; Blake, D.J.; Ling, G.; Wakabayashi, N.; Fujii, J.; Myers, A.; Biswal, S. Nrf2-regulated glutathione recycling independent of biosynthesis is critical for cell survival during oxidative stress. Free Radic. Biol. Med. 2009, 46, 443–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samiec, P.S.; Drews-Botsch, C.; Flagg, E.W.; Kurtz, J.C.; Sternberg, P., Jr.; Reed, R.L.; Jones, D.P. Glutathione in human plasma: Decline in association with aging, age-related macular degeneration, and diabetes. Free Radic. Biol. Med. 1998, 24, 699–704. [Google Scholar] [CrossRef]
- Moriarty, S.E.; Shah, J.H.; Lynn, M.; Jiang, S.; Openo, K.; Jones, D.P.; Sternberg, P. Oxidation of glutathione and cysteine in human plasma associated with smoking. Free Radic. Biol. Med. 2003, 35, 1582–1588. [Google Scholar] [CrossRef] [PubMed]
- Ashfaq, S.; Abramson, J.L.; Jones, D.P.; Rhodes, S.D.; Weintraub, W.S.; Hooper, W.C.; Vaccarino, V.; Harrison, D.G.; Quyyumi, A.A. The relationship between plasma levels of oxidized and reduced thiols and early atherosclerosis in healthy adults. J. Am. Coll. Cardiol. 2006, 47, 1005–1011. [Google Scholar] [CrossRef]
- Yeh, M.Y.; Burnham, E.L.; Moss, M.; Brown, L.A. Chronic alcoholism alters systemic and pulmonary glutathione redox status. Am. J. Respir. Crit. Care Med. 2007, 176, 270–276. [Google Scholar] [CrossRef]
- Vincent, H.K.; Innes, K.E.; Vincent, K.R. Oxidative stress and potential interventions to reduce oxidative stress in overweight and obesity. Diabetes Obes. Metab. 2007, 9, 813–839. [Google Scholar] [CrossRef]
- Lang, C.A.; Mills, B.J.; Mastropaolo, W.; Liu, M.C. Blood glutathione decreases in chronic diseases. J. Lab. Clin. Med. 2000, 135, 402–405. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Meydani, S.N.; Sastre, J.; Hayek, M.; Meydani, M. In vitro glutathione supplementation enhances interleukin-2 production and mitogenic response of peripheral blood mononuclear cells from young and old subjects. J. Nutr. 1994, 124, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Arranz, L.; Fernández, C.; Rodríguez, A.; Ribera, J.M.; De la Fuente, M. The glutathione precursor N-acetylcysteine improves immune function in postmenopausal women. Free Radic. Biol. Med. 2008, 45, 1252–1262. [Google Scholar] [CrossRef] [PubMed]
- Millman, A.C.; Salman, M.; Dayaram, Y.K.; Connell, N.D.; Venketaraman, V. Natural killer cells, glutathione, cytokines, and innate immunity against Mycobacterium tuberculosis. J. Interferon Cytokine Res. 2008, 28, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Venketaraman, V.; Millman, A.; Salman, M.; Swaminathan, S.; Goetz, M.; Lardizabal, A.; David, H.; Connell, N.D. Glutathione levels and immune responses in tuberculosis patients. Microb. Pathog. 2008, 44, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, A.; Tsuyuki, S.; Inamoto, T.; Yamaoka, Y. Liver immunity and glutathione. Antioxid. Redox Signal. 1999, 1, 245–253. [Google Scholar] [CrossRef]
- Guerra, C.; Morris, D.; Sipin, A.; Kung, S.; Franklin, M.; Gray, D.; Tanzil, M.; Guilford, F.; Khasawneh, F.T.; Venketaraman, V. Glutathione and adaptive immune responses against Mycobacterium tuberculosis infection in healthy and HIV infected individuals. PLoS ONE 2011, 6, e28378. [Google Scholar] [CrossRef]
- Villa, P.; Saccani, A.; Sica, A.; Ghezzi, P. Glutathione protects mice from lethal sepsis by limiting inflammation and potentiating host defense. J. Infect. Dis. 2002, 185, 1115–1120. [Google Scholar] [CrossRef]
- Cai, J.; Chen, Y.; Seth, S.; Furukawa, S.; Compans, R.W.; Jones, D.P. Inhibition of influenza infection by glutathione. Free Radic. Biol. Med. 2003, 34, 928–936. [Google Scholar]
- Morris, D.; Khurasany, M.; Nguyen, T.; Kim, J.; Guilford, F.; Mehta, R.; Gray, D.; Saviola, B.; Venketaraman, V. Glutathione and infection. Biochim. Biophys. Acta 2013, 1830, 3329–3349. [Google Scholar] [CrossRef]
- Levring, T.B.; Kongsbak, M.; Rode, A.K.; Woetmann, A.; Ødum, N.; Bonefeld, C.M.; Geisler, C. Human CD4+ T cells require exogenous cystine for glutathione and DNA synthesis. Oncotarget 2015, 6, 21853–21864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, F.M.; Shang, H.S.; Tang, N.Y.; Lin, J.J.; Lu, K.W.; Lin, J.P.; Ko, Y.C.; Yu, C.C.; Wang, H.L.; Liao, J.C.; et al. Effects of diallyl trisulfide on induction of apoptotic death in murine leukemia WEHI-3 cells in vitro and alterations of the immune responses in normal and leukemic mice in vivo. Environ. Toxicol. 2015, 30, 1343–1353. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, M.; Mohammad Hassan, Z.; Mostafaie, A.; Zare Mehrjardi, N.; Ghazanfari, T. Purif ied Protein Fraction of Garlic Extract Modulates Cellular Immune Response against Breast Transplanted Tumors in BALB/c Mice Model. Cell J. 2013, 15, 65–75. [Google Scholar] [PubMed]
- Hassan, Z.M.; Yaraee, R.; Zare, N.; Ghazanfari, T.; Sarraf Nejad, A.H.; Nazori, B. Immunomodulatory affect of R10 fraction of garlic extract on natural killer activity. Int. Immunopharmacol. 2003, 3, 1483–1489. [Google Scholar] [CrossRef]
- Charron, C.S.; Dawson, H.D.; Albaugh, G.P.; Solverson, P.M.; Vinyard, B.T.; Solano-Aguilar, G.I.; Molokin, A.; Novotny, J.A. A Single Meal Containing Raw, Crushed Garlic Influences Expression of Immunity- and Cancer-Related Genes in Whole Blood of Humans. J. Nutr. 2015, 145, 2448–2455. [Google Scholar] [CrossRef] [Green Version]
- Quintero-Fabian, S.; Ortuño-Sahagún, D.; Vázquez-Carrera, M.; López-Roa, R.I. Alliin, a garlic (Allium sativum) compound, prevents LPS-induced inflammation in 3T3-L1 adipocytes. Mediat. Inflamm. 2013, 2013, 381815. [Google Scholar] [CrossRef] [PubMed]
- Saud, S.M.; Li, W.; Gray, Z.; Matter, M.S.; Colburn, N.H.; Young, M.R.; Kim, Y.S. Diallyl Disulfide (DADS), a Constituent of Garlic, Inactivates NF-kappaB and Prevents Colitis-Induced Colorectal Cancer by Inhibiting GSK-3beta. Cancer Prev. Res. 2016, 9, 607–615. [Google Scholar] [CrossRef]
- Shi, L.; Lin, Q.; Li, X.; Nie, Y.; Sun, S.; Deng, X.; Wang, L.; Lu, J.; Tang, Y.; Luo, F. Alliin, a garlic organosulfur compound, ameliorates gut inflammation through MAPK-NF-kappaB/AP-1/STAT-1 inactivation and PPAR-gamma activation. Mol. Nutr. Food Res. 2017, 61, 1601013. [Google Scholar] [CrossRef]
- Kyo, E.; Uda, N.; Suzuki, A.; Kakimoto, M.; Ushijima, M.; Kasuga, S.; Itakura, Y. Immunomodulation and antitumor activities of Aged Garlic Extract. Phytomedicine 1998, 5, 259–267. [Google Scholar] [CrossRef]
- Larypoor, M.; Bayat, M.; Zuhair, M.H.; Akhavan Sepahy, A.; Amanlou, M. Evaluation of The Number of CD4(+) CD25(+) FoxP3(+) Treg Cells in Normal Mice Exposed to AFB1 and Treated with Aged Garlic Extract. Cell J. 2013, 15, 37–44. [Google Scholar]
- Fallah-Rostami, F.; Tabari, M.A.; Esfandiari, B.; Aghajanzadeh, H.; Behzadi, M.Y. Immunomodulatory activity of aged garlic extract against implanted fibrosarcoma tumor in mice. N. Am. J. Med. Sci. 2013, 5, 207–212. [Google Scholar] [PubMed]
- Nillert, N.; Pannangrong, W.; Welbat, J.U.; Chaijaroonkhanarak, W.; Sripanidkulchai, K.; Sripanidkulchai, B. Neuroprotective Effects of Aged Garlic Extract on Cognitive Dysfunction and Neuroinflammation Induced by β-Amyloid in Rats. Nutrients 2017, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Nantz, M.P.; Rowe, C.A.; Muller, C.E.; Creasy, R.A.; Stanilka, J.M.; Percival, S.S. Supplementation with aged garlic extract improves both NK and gammadelta-T cell function and reduces the severity of cold and flu symptoms: A randomized, double-blind, placebo-controlled nutrition intervention. Clin. Nutr. 2012, 31, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Mathews, A.E.; Rodrigues, C.; Eudy, B.J.; Rowe, C.A.; O’Donoughue, A.; Percival, S.S. Aged garlic extract supplementation modifies inflammation and immunity of adults with obesity: A randomized, double-blind, placebo-controlled clinical trial. Clin. Nutr. ESPEN 2018, 24, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Anandasadagopan, S.K.; Sundaramoorthy, C.; Pandurangan, A.K.; Nagarajan, V.; Srinivasan, K.; Ganapasam, S. S-Allyl cysteine alleviates inflammation by modulating the expression of NF-B during chromium (VI)-induced hepatotoxicity in rats. Hum. Exp. Toxicol. 2017, 36, 1186–1200. [Google Scholar] [CrossRef] [PubMed]
- Zanardo, R.C.; Brancaleone, V.; Distrutti, E.; Fiorucci, S.; Cirino, G.; Wallace, J.L. Hydrogen sulfide is an endogenous modulator of leukocyte-mediated inflammation. FASEB J. 2006, 20, 2118–2120. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Guo, C.; Wu, D.; Zhang, A.; Gu, T.; Wang, L.; Wang, C. Hydrogen sulfide inhibits the development of atherosclerosis with suppressing CX3CR1 and CX3CL1 expression. PLoS ONE 2012, 7, e41147. [Google Scholar] [CrossRef]
- Wallace, J.L.; Wang, R. Hydrogen sulfide-based therapeutics: Exploiting a unique but ubiquitous gasotransmitter. Nat. Rev. Drug Discov. 2015, 14, 329–345. [Google Scholar] [CrossRef]
- Candela, J.J.; Macrophages, W.C. Drive Microvascular Dysfunction in Obesity by Depleting Vessel Hydrogen Sulfide. FASEB J. 2017, 37, 889–899. [Google Scholar]
- Rios, E.C.; Szczesny, B.; Soriano, F.G.; Olah, G.; Szabo, C. Hydrogen sulfide attenuates cytokine production through the modulation of chromatin remodeling. Int. J. Mol. Med. 2015, 35, 1741–1746. [Google Scholar] [CrossRef] [Green Version]
- Fiorucci, S.; Orlandi, S.; Mencarelli, A.; Caliendo, G.; Santagada, V.; Distrutti, E.; Santucci, L.; Cirino, G.; Wallace, J.L. Enhanced activity of a hydrogen sulphide-releasing derivative of mesalamine (ATB-429) in a mouse model of colitis. Br. J. Pharmacol. 2007, 150, 996–1002. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.L.; Vong, L.; McKnight, W.; Dicay, M.; Martin, G.R. Endogenous and exogenous hydrogen sulfide promotes resolution of colitis in rats. Gastroenterology 2009, 137, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Kaya-Yasar, Y.; Karaman, Y.; Bozjurt, T.E.; Onder, S.C.; Sahin-Erdemli, I. Effects of intranasal treatment with slow (GYY4137) and rapid (NaHS) donors of hydrogen sulfide in lipopolysaccharide-induced airway inflammation in mice. Pulm. Pharmacol. Ther. 2017, 45, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Luo, N.; Wang, L.; Zhao, Z.; Bu, H.; Xu, G.; Yan, Y.; Che, X.; Jiao, Z.; Zhao, T.; et al. Hydrogen sulfide ameliorates chronic renal failure in rats by inhibiting apoptosis and inflammation through ROS/MAPK and NF-kappaB signaling pathways. Sci. Rep. 2017, 7, 455. [Google Scholar] [CrossRef] [PubMed]
- Sen, N.; Paul, B.D.; Gadalla, M.M.; Mustafa, A.K.; Sen, T.; Xu, R.; Kim, S.; Snyder, S.H. Hydrogen sulfide-linked sulfhydration of NF-kappaB mediates its antiapoptotic actions. Mol. Cell 2012, 45, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Badiei, A.; Rivers-Auty, J.; Ang, A.D.; Bhatia, M. Inhibition of hydrogen sulfide production by gene silencing attenuates inflammatory activity by downregulation of NF-kappaB and MAP kinase activity in LPS-activated RAW 264.7 cells. Biomed. Res. Int. 2014, 2014, 848570. [Google Scholar] [CrossRef] [PubMed]
- Stuhlmeier, K.M.; Broll, J.; Iliev, B. NF-kappaB independent activation of a series of proinflammatory genes by hydrogen sulfide. Exp. Biol. Med. 2009, 234, 1327–1338. [Google Scholar] [CrossRef]
- Niu, W.N.; Yadav, P.K.; Adamec, J.; Benerjee, R. S-glutathionylation enhances human cystathionine beta-synthase activity under oxidative stress conditions. Antioxid. Redox Signal. 2015, 22, 350–361. [Google Scholar] [CrossRef]
- Liang, D.; Wu, H.; Wong, M.W.; Huang, D. Diallyl Trisulfide Is a Fast H2S Donor, but Diallyl Disulfide Is a Slow One: The Reaction Pathways and Intermediates of Glutathione with Polysulfides. Org. Lett. 2015, 17, 4196–4199. [Google Scholar] [CrossRef]
- DeLeon, E.R.; Gao, Y.; Huang, E.; Olson, K.R. Garlic oil polysulfides: H2S- and O2-independent prooxidants in buffer and antioxidants in cells. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R1212–R1225. [Google Scholar] [CrossRef]
- Chuah, S.C.; Moore, P.K.; Zhu, Y.Z. S-allylcysteine mediates cardioprotection in an acute myocardial infarction rat model via a hydrogen sulfide-mediated pathway. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H2693–H2701. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Model | Treatment | Dose | Immune Response | Reference |
|---|---|---|---|---|
| Cell culture of isolated DC and T cells from study mice | Cysteine | T Cell Proliferation Assay: 50 μM cysteine Metabolic Labeling: 1 μCi/mL [35S] cysteine per 2 × 106 cells | -Redox modeling of Tregs through reduction of GSH synthesis in DC. -Blocking of GSH redistribution from the nucleus to the cytoplasm in Tregs in response to T cell activation. | [96] |
| Cell culture of isolated leukocytes from healthy human blood | GSH-oxidizing agents (diamide and BHP) | 50 nmoL per 106 cells | -Regulation of phagocytosis in PMN through the inhibition of the assembly of microtubules and consequently reduction of H2O2 production in GSH homeostasis. | [97] |
| Cell culture of isolated splenic naive T cells from mice | NAC in GSH depleted cells | 20 mM NAC per 106 cells in the presence of 1 mM BSO | -Inhibition of T cell proliferation and reduction IL-2 production in GSH depleted cells. -Stimulation of T cell proliferation and increase IL-2 production in restored GSH cells. | [98] |
| Cell culture of isolated glomerular mesangial cell from rats | GSH | 3–5 mM GSH per 0.5–1 × 106 cells | -Inhibition of the complement activation cascade and the binding of antibodies to antigens. | [99] |
| Human randomized controlled clinical trial in HIV-infected patients | NAC | ~7 g/day, oral | -Increase in GSH levels in the blood and in CD4 T cells. | [100] |
| Human randomized controlled clinical trial in HIV-infected patients | Liposomal-GSH | 1.26 g/day, oral | -Increase in the levels of total GSH in CD4 T cells, IL-6, IL-12, IL-2, IFN-γ and reduction in the levels of IL-10. | [101] |
| Patients with cirrhosis | GSH precursor (oxathizolidine-4-carboxylic acid) | 210 mg/kg/day, intravenous | -Reduction in the levels of IL-8 and IL-6 from isolated monocytes. | [102] |
| Model | Treatment | Dose | Immune response | Reference |
|---|---|---|---|---|
| Cell culture of isolated PBMC from healthy individuals | Fresh garlic extract | 0.5–4 μg/mL per 106 cells | -Decrease of IL-17 expression when stimulated with PHA. | [48] |
| Cell culture of 3T3-L1 adipocytes | Alliin | 100 μmol/L (number of cells were not specified) | -Down-regulation of IL-6, MCP-1 genes expression and decrease of IL-6, MCP-1 protein levels. -Up-regulation of genes involved with immune response. | [127] |
| Cell culture of human colorectal cancer SW480 cells | DADS | 2.5–40 μmol/L (number of cells were not specified) | -Reduction of phosphorylation and nuclear translocation of NF-κB/p65 protein in a dose dependent way. | [128] |
| Cell culture of isolated splenocytes from mice | AGE | 0.5-1 v/v% per 0.2–1 × 107 cells | -Increase in NK cell activation and cytotoxic T cells with suppression of tumor cell growth. -Increase of IL-2, TNF-α and IFN-γ levels in spleen cells. | [130] |
| Cell culture of Jurkat T cells | SAC | 0, 0.5, 1, 1.5, or 2 mg/mL of SAC in 106 cells | -Inhibition of NF-κB p50/65 heterodimer activation. | [3] |
| Mice with leukemia | DATS | 1 and 10 mg/kg, oral | -Increase of phagocytosis activity on PBMC. | [123] |
| Mice with implanted breast tumors | Purified protein fraction from garlic extract | 0.01–0.04 mg/mL, inoculation into the lesion | -Increase of T-cell activation and increase of intra-tumor infiltration of lymphocytes in all doses | [124] |
| Mice with alcoholic fatty liver disease | Allicin | 5 mg/kg/day and 20 mg/kg/day, oral | -Reduction of TNF-α, IL-1β and IL-6 levels with 20 mg dose. | [46] |
| Mice | Purified fraction of garlic extract | 10–40 mg/mL, oral | -Increase of NK activity with the 20 mg/mL dose. | [125] |
| Mice with colitis | Alliin | 500 mg/kg, oral | -Suppression of LPS-induced AP-1/NF-κB/STAT-1 activation. | [129] |
| Mice exposed to Aflatoxin B1 | AGE | 20 mg/kg/day, intraperitoneal | -Decrease in IFN-γ and IL-4 levels in isolated splenocytes. -Decrease in the number of Treg in the spleen. | [131] |
| Mice with implanted fibrosarcoma tumor | AGE | 100 mg/kg (containing 0.4 g of garlic/mL), intraperitoneal | -Increase in spleen lymphocyte subpopulation ratio and IFN-γ levels. -Increase in the amount of CD4+/CD8+ ratio. | [132] |
| Rats with induced neuroinflammation | AGE | 125, 250, 500 mg/kg, oral | -Decrease in the density of CD11b-positive microglia immunoreactivity and decrease in up-regulation of IL-1β in hippocampal region at all doses. | [133] |
| Mice with diabetes | SAC | 0.5 g/L, oral by drinking water | -Reduction in renal levels of IL-6 and TNF-α. -Suppression of renal NF-κB activity, mRNA expression and protein production. | [5] |
| Rats with diabetes | SAC | 150 mg/kg/day, oral | -Reduction in hippocampal NF-κB, TLR4 and TNF-α. | [6] |
| Rats with chromium (VI)-induced hepatotoxicity | SAC | 100 mg/kg, oral | -Decrease in the expression of NF-κB, TNF-α and iNOS in the liver. | [136] |
| Human randomized crossover study in healthy individuals | Raw crushed garlic | 5 g, oral | -Increase of NFAM1 gene expression. | [126] |
| Human randomized controlled clinical trial in patients with advanced colon, liver and pancreatic cancer | AGE | 500 mg/day, oral | -Increase in NK cell count and activity. | [64] |
| Human randomized controlled clinical trial in healthy individuals | AGE | 2.56 g/day, oral | -Increase in γδ-T cells and NK cells. -Attenuation of the severity of reported of cold and flu-like symptoms. | [134] |
| Human randomized controlled clinical trial in obese individuals | AGE | 3.6 g/day, oral | -Increase in γδ-T cell population and decrease in NKT cell population, IL-6 and TNF-α levels in the serum. | [135] |
| Model | Treatment | Dose | Immune response | Reference |
|---|---|---|---|---|
| Cell culture of RAW264.7 cells | FW1256 (H2S donor) | 200 μM per 2 × 105 or 106 cells | -Decrease in TNF-α, IL-6, PGE2, NO levels and expression of IL-1β, COX2, and iNOS. -Inhibition of NF-κB activation. | [14] |
| Cell culture of THP-1 cells | NaHS | 0.01, 0.1, 0.5 or 1 mM per 2 × 106 cells | -Decrease in IL-6 and TNF-α levels in a dose dependent manner. | [141] |
| Cell culture of U937 cells | NaHS | 0.01, 0.1, or 1 mM (number of cells were not specified) | -Increase in mRNA expression and protein levels of TNF-α, IL-1β and IL-6 with 0.1 and 1mM doses. -Up-regulation of CD11b with 0.1 mM dose. -Increase in NF-κB p65 activity with 0.1 and 1mM doses. | [13] |
| Cell culture of human fibroblast-like synoviocytes | NaHS | 0.05–5 mM (number of cells were not specified) | -Increase in mRNA expression of TNF-α, IL-8, IL-1α, IL-1β and COX-2 at 2 mM dose. -Increase in mRNA expression of IL-8 and COX-2 at doses below 0.25 mM. -NaHS did not affect NF-κB activation at none of the doses. | [148] |
| Mice with colitis | ATB-429 (H2S donor) | 33–130 mg/kg, oral | -Reduction of mRNA expression of TNF-α, IFN-γ, IL-1, IL-2, IL-12 p40 and RANTES. | [142] |
| Rats with colitis | NaHS and Lawesson’s reagent (H2S donors) | 30 μmol/kg, intracolonically | -Reduction of mRNA expression of TNF-α with both donors. -Reduction in TNF-α protein levels with NaHS. | [143] |
| Mice with LPS-induced airway inflammation | GYY4137 (H2S donor) and NaHS | GYY4137:0.3 or 1 mg/kg NaHS: 1 or 3 mg/kg, intranasal | -Prevention of neutrophil increase in bronchoalveolar lavage fluid with both donors. -Prevention of IL-1β levels increase with both donors. | [144] |
| Rats with chronic renal failure | NaHS | 100 μmol/kg, intraperitoneal | -Decrease in TNF-α, IL-6, IL-10, and MCP-1 renal levels with both donors. -Decrease in NF-κB-p50, p65, and p-p65 levels with both donors. | [145] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, C.; Percival, S.S. Immunomodulatory Effects of Glutathione, Garlic Derivatives, and Hydrogen Sulfide. Nutrients 2019, 11, 295. https://doi.org/10.3390/nu11020295
Rodrigues C, Percival SS. Immunomodulatory Effects of Glutathione, Garlic Derivatives, and Hydrogen Sulfide. Nutrients. 2019; 11(2):295. https://doi.org/10.3390/nu11020295
Chicago/Turabian StyleRodrigues, Camila, and Susan S. Percival. 2019. "Immunomodulatory Effects of Glutathione, Garlic Derivatives, and Hydrogen Sulfide" Nutrients 11, no. 2: 295. https://doi.org/10.3390/nu11020295
APA StyleRodrigues, C., & Percival, S. S. (2019). Immunomodulatory Effects of Glutathione, Garlic Derivatives, and Hydrogen Sulfide. Nutrients, 11(2), 295. https://doi.org/10.3390/nu11020295