Why Is Very High Cholesterol Content Beneficial for the Eye Lens but Negative for Other Organs?


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
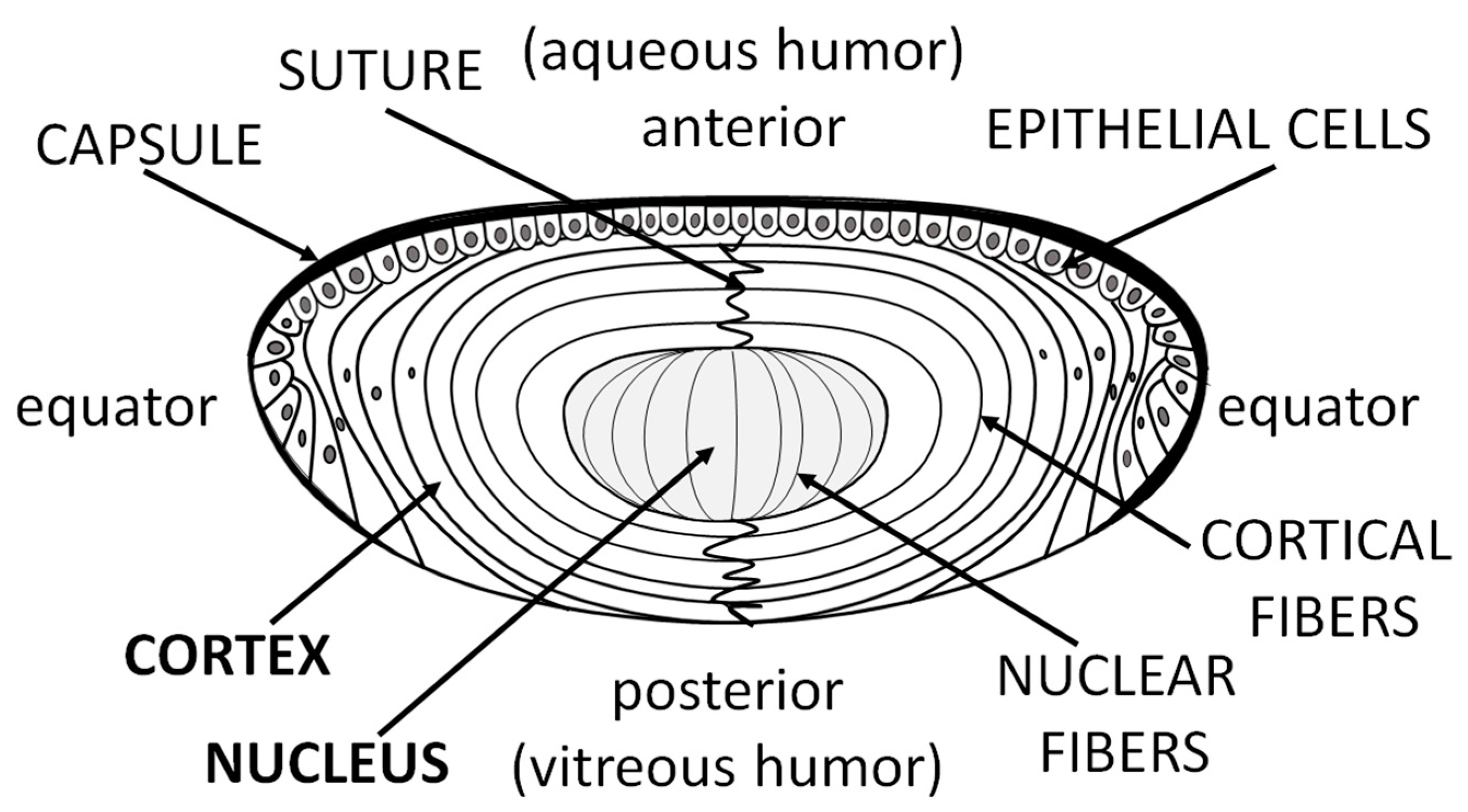
2. Human Lens Cholesterol
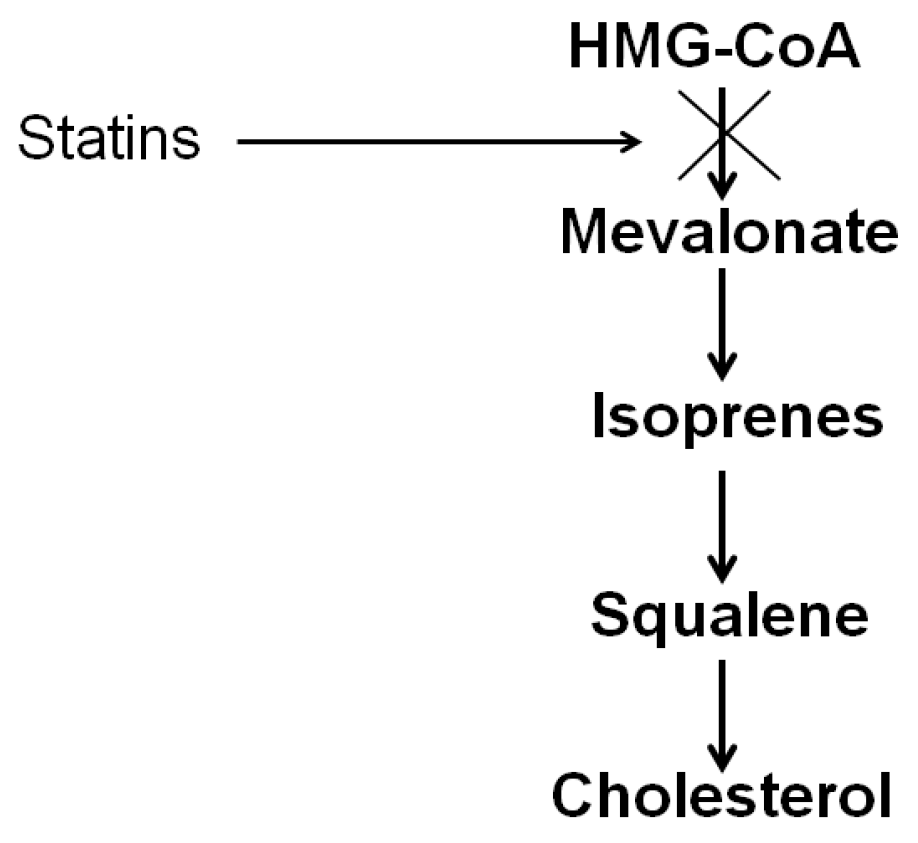
2.1. Cholesterol Synthesis
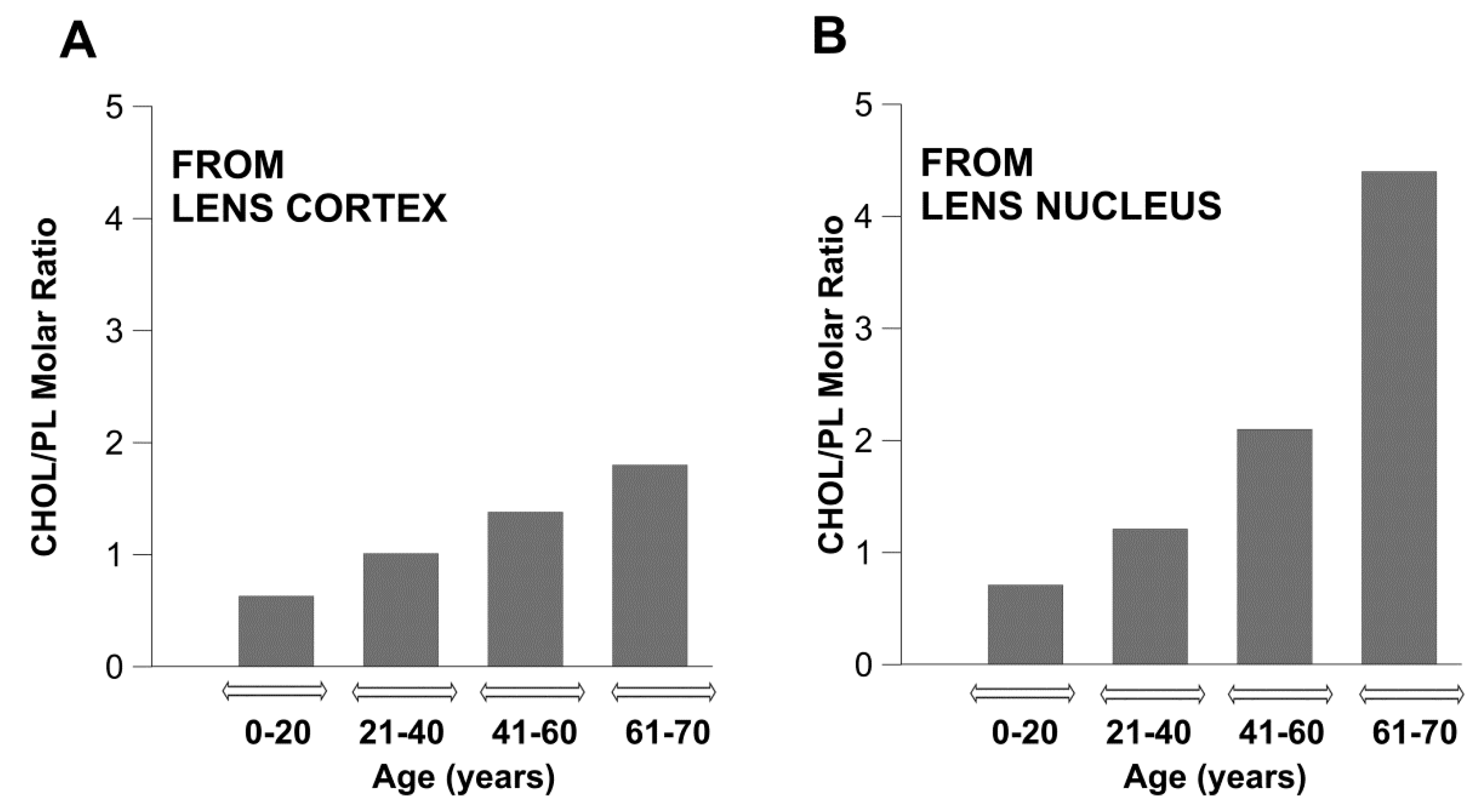
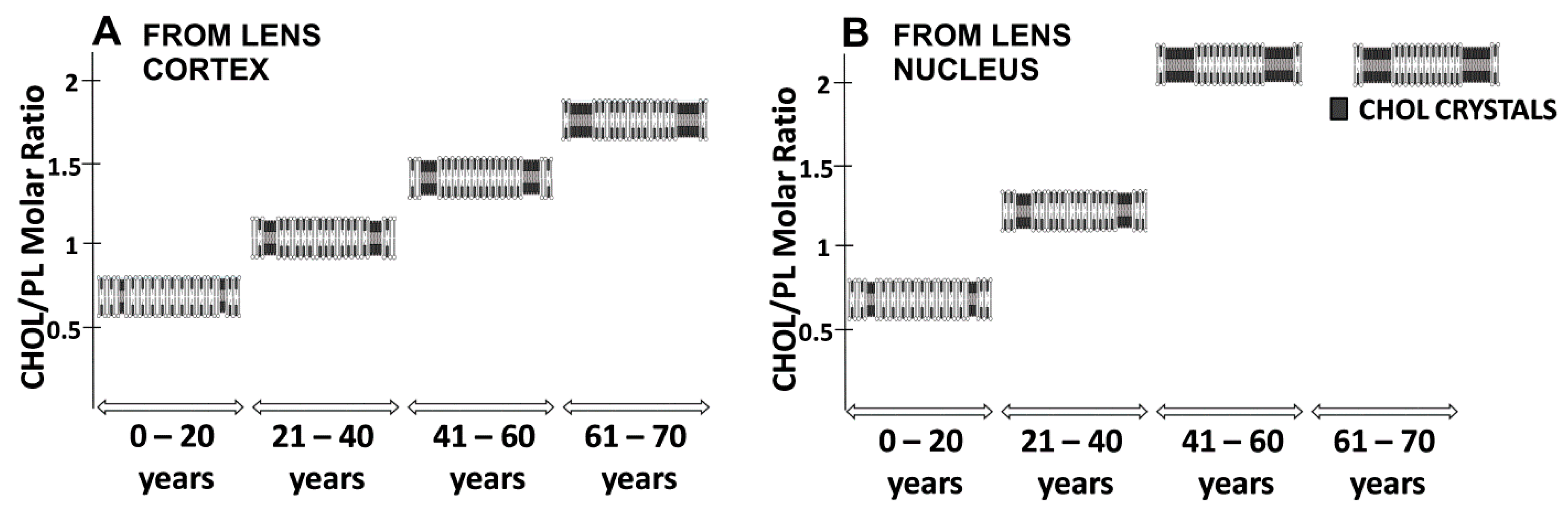
2.2. Changes with Age and Cataract
2.3. Cholesterol-Lowering Drugs and the Human Diet
3. Mechanisms Maintaining the Saturating Level of Cholesterol in Phospholipid Bilayers
3.1. Lipid Composition Changes
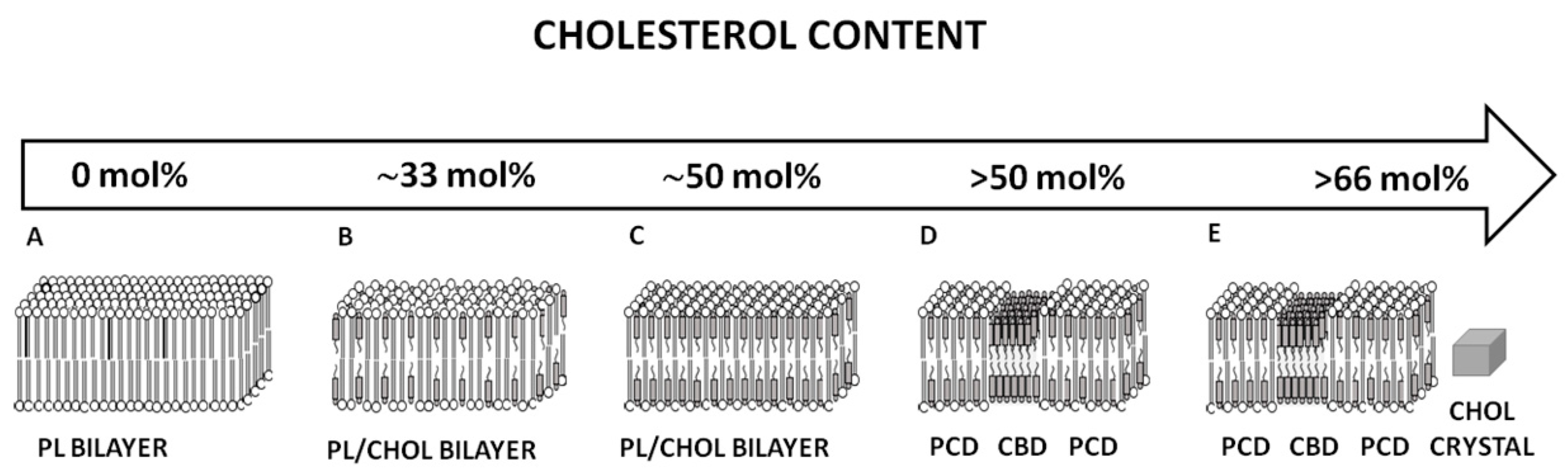
3.2. Cholesterol Bilayer Domain Formation
4. Function of Cholesterol in the Lens
4.1. Fiber Cell Membrane Homeostasis
4.2. Fiber Cell Homeostasis
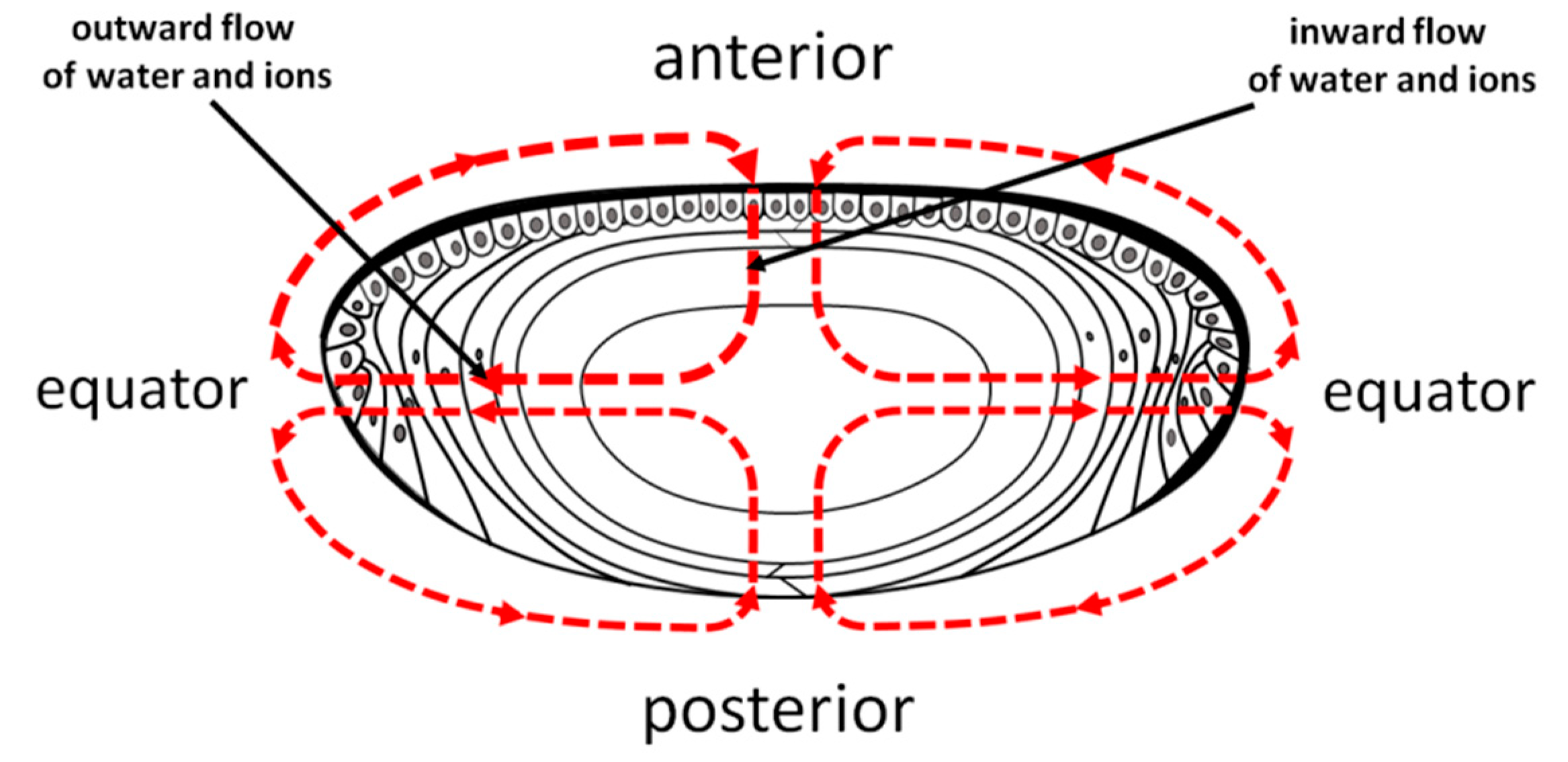
4.3. Lens Homeostasis and Maintaining Lens Transparency
5. Other Tissue/Organ Cells
5.1. High Cholesterol and Oxidative Stress
5.2. Cholesterol-Lowering Drugs and the Human Diet
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Li, L.K.; So, L.; Spector, A. Membrane cholesterol and phospholipid in consecutive concentric sections of human lenses. J. Lipid Res. 1985, 26, 600–609. [Google Scholar]
- Mainali, L.; Raguz, M.; O’Brien, W.J.; Subczynski, W.K. Properties of membranes derived from the total lipids extracted from clear and cataractous lenses of 61–70-year-old human donors. Eur. Biophys. J. 2015, 44, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Yappert, M.C.; Rujoi, M.; Borchman, D.; Vorobyov, I.; Estrada, R. Glycero-versus sphingo-phospholipids: Correlations with human and non-human mammalian lens growth. Exp. Eye Res. 2003, 76, 725–734. [Google Scholar] [CrossRef]
- Huang, L.; Grami, V.; Marrero, Y.; Tang, D.; Yappert, M.C.; Rasi, V.; Borchman, D. Human lens phospholipid changes with age and cataract. Investig. Ophthalmol. Vis. Sci. 2005, 46, 1682–1689. [Google Scholar] [CrossRef]
- Hughes, J.R.; Deeley, J.M.; Blanksby, S.J.; Leisch, F.; Ellis, S.R.; Truscott, R.J.W.; Mitchell, T.W. Instability of the cellular lipidome with age. Age 2012, 34, 935–947. [Google Scholar] [CrossRef] [PubMed]
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Preston Mason, R.; Tulenko, T.N.; Jacob, R.F. Direct evidence for cholesterol crystalline domains in biological membranes: Role in human pathobiology. Biochim. Biophys. Acta Biomembr. 2003, 1610, 198–207. [Google Scholar] [CrossRef]
- Grebe, A.; Latz, E. Cholesterol Crystals and Inflammation. Curr. Rheumatol. Rep. 2013, 15, 313. [Google Scholar] [CrossRef] [PubMed]
- Lim, R.S.; Suhalim, J.L.; Miyazaki-Anzai, S.; Miyazaki, M.; Levi, M.; Potma, E.O.; Tromberg, B.J. Identification of cholesterol crystals in plaques of atherosclerotic mice using hyperspectral CARS imaging. J. Lipid Res. 2011, 52, 2177–2186. [Google Scholar] [CrossRef] [Green Version]
- Janoudi, A.; Shamoun, F.E.; Kalavakunta, J.K.; Abela, G.S. Cholesterol crystal induced arterial inflammation and destabilization of atherosclerotic plaque. Eur. Heart J. 2016, 37, 1959–1967. [Google Scholar] [CrossRef]
- Cenedella, R.J. Sterol synthesis by the ocular lens of the rat during postnatal development. J. Lipid Res. 1982, 23, 619–626. [Google Scholar]
- Brown, M.S.; Goldstein, J.L. How LDL Receptors Influence Cholesterol and Atherosclerosis. Sci. Am. 1984, 251, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Kurze, V.; Huber, T.; Afdhal, N.H.; Beyer, K.; Hamilton, J.A. A solid-state NMR study of phospholipid-cholesterol interactions: Sphingomyelin-cholesterol binary systems. Biophys. J. 2002, 83, 1465–1478. [Google Scholar] [CrossRef]
- Bogren, H.; Larsson, K. An X-ray-diffraction study of crystalline cholesterol in some pathological deposits in man. Biochim. Biophys. Acta 1963, 75, 65–69. [Google Scholar] [CrossRef]
- Nealon, J.R.; Blanksby, S.J.; Abbott, S.K.; Hulbert, A.J.; Mitchell, T.W.; Truscott, R.J.W. Phospholipid composition of the rat lens is independent of diet. Exp. Eye Res. 2008, 87, 502–514. [Google Scholar] [CrossRef] [PubMed]
- Zelenka, P.S. Lens lipids. Curr. Eye Res. 1984, 3, 1337–1359. [Google Scholar] [CrossRef] [PubMed]
- Cenedella, R.J. Cholesterol and cataracts. Surv. Ophthalmol. 1996, 40, 320–337. [Google Scholar] [CrossRef]
- Berman, E.R. Biochemistry of the Eye; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013; ISBN 978-1-4757-9441-0. [Google Scholar]
- De Vries, A.C.; Vermeer, M.A.; Hendriks, A.L.; Bloemendal, H.; Cohen, L.H. Biosynthetic capacity of the human lens upon aging. Exp. Eye Res. 1991, 53, 519–524. [Google Scholar] [CrossRef]
- De Vries, A.C.; Vermeer, M.A.; Bredman, J.J.; Bär, P.R.; Cohen, L.H. Cholesterol content of the rat lens is lowered by administration of simvastatin, but not by pravastatin. Exp. Eye Res. 1993, 56, 393–399. [Google Scholar] [CrossRef]
- Cenedella, R.J. Lipoproteins and lipids in cow and human aqueous humor. Biochim. Biophys. Acta 1984, 793, 448–454. [Google Scholar] [CrossRef]
- Brewer, H.B. Clinical significance of plasma lipid levels. Am. J. Cardiol. 1989, 64, 3G–9G. [Google Scholar] [CrossRef]
- Mainali, L.; Raguz, M.; O’Brien, W.J.; Subczynski, W.K. Changes in the properties and organization of human lens lipid membranes occurring with age. Curr. Eye Res. 2017, 42, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Lynnerup, N.; Kjeldsen, H.; Heegaard, S.; Jacobsen, C.; Heinemeier, J. Radiocarbon dating of the human eye lens crystallines reveal proteins without carbon turnover throughout life. PLoS ONE 2008, 3, e1529. [Google Scholar] [CrossRef] [PubMed]
- Jacob, R.F.; Cenedella, R.J.; Mason, R.P. Evidence for Distinct Cholesterol Domains in Fiber Cell Membranes from Cataractous Human Lenses. J. Biol. Chem. 2001, 276, 13573–13578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapourchali, F.R.; Surendiran, G.; Goulet, A.; Moghadasian, M.H. The Role of Dietary Cholesterol in Lipoprotein Metabolism and Related Metabolic Abnormalities: A Mini-review. Crit. Rev. Food Sci. Nutr. 2016, 56, 2408–2415. [Google Scholar] [CrossRef] [PubMed]
- Porter, F.D.; Herman, G.E. Malformation syndromes caused by disorders of cholesterol synthesis. J. Lipid Res. 2011, 52, 6–34. [Google Scholar] [CrossRef] [PubMed]
- Kretzer, F.L.; Hittner, H.M.; Mehta, R.S. Ocular Manifestations of the Smith-Lemli-Opitz Syndrome. Arch. Ophthalmol. 1981, 99, 2000–2006. [Google Scholar] [CrossRef] [PubMed]
- Cotlier, E.; Rice, P. Cataracts in the Smith-Lemli-Opitz syndrome. Am. J. Ophthalmol. 1971, 72, 955–959. [Google Scholar] [CrossRef]
- Simon, A.; Kremer, H.P.H.; Wevers, R.A.; Scheffer, H.; De Jong, J.G.; Van Der Meer, J.W.M.; Drenth, J.P.H. Mevalonate kinase deficiency: Evidence for a phenotypic continuum. Neurology 2004, 62, 994–997. [Google Scholar] [CrossRef] [PubMed]
- Wilker, S.C.; Dagnelie, G.; Goldberg, M.F. Retinitis pigmentosa and punctate cataracts in mevalonic aciduria. Retin. Cases Brief. Rep. 2010, 4, 34–36. [Google Scholar] [CrossRef]
- Federico, A.; Dotti, M.T. Cerebrotendinous xanthomatosis: Clinical manifestations, diagnostic criteria, pathogenesis, and therapy. J. Child. Neurol. 2003, 18, 633–638. [Google Scholar] [CrossRef]
- Cruysberg, J.R.; Wevers, R.A.; Tolboom, J.J. Juvenile cataract associated with chronic diarrhea in pediatric cerebrotendinous xanthomatosis. Am. J. Ophthalmol. 1991, 112, 606–607. [Google Scholar] [CrossRef]
- Traupe, H.; Müller, D.; Atherton, D.; Kalter, D.C.; Cremers, F.P.M.; van Oost, B.A.; Ropers, H.-H. Exclusion mapping of the X-linked dominant chondrodysplasia punctata/ichthyosis/cataract/short stature (Happle) syndrome: Possible involvement of an unstable pre-mutation. Hum. Genet. 1992, 89, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Pyrah, I.T.; Kalinowski, A.; Jackson, D.; Davies, W.; Davis, S.; Aldridge, A.; Greaves, P. Toxicologic lesions associated with two related inhibitors of oxidosqualene cyclase in the dog and mouse. Toxicol. Pathol. 2001, 29, 174–179. [Google Scholar] [CrossRef]
- Cenedella, R.J.; Kuszak, J.R.; Al-Ghoul, K.J.; Qin, S.; Sexton, P.S. Discordant expression of the sterol pathway in lens underlies simvastatin-induced cataracts in Chbb: Thom rats. J. Lipid Res. 2003, 44, 198–211. [Google Scholar] [CrossRef] [PubMed]
- Gerson, R.J.; MacDonald, J.S.; Alberts, A.W.; Chen, J.; Yudkovitz, J.B.; Greenspan, M.D.; Rubin, L.F.; Bokelman, D.L. On the etiology of subcapsular lenticular opacities produced in dogs receiving HMG-CoA reductase inhibitors. Exp. Eye Res. 1990, 50, 65–78. [Google Scholar] [CrossRef]
- Lai, C.-L.; Shau, W.-Y.; Chang, C.-H.; Chen, M.-F.; Lai, M.-S. Statin use and cataract surgery: A nationwide retrospective cohort study in elderly ethnic Chinese patients. Drug Saf. 2013, 36, 1017–1024. [Google Scholar] [CrossRef] [PubMed]
- Leuschen, J.; Mortensen, E.M.; Frei, C.R.; Mansi, E.A.; Panday, V.; Mansi, I. Association of statin use with cataracts: A propensity score-matched analysis. JAMA Ophthalmol. 2013, 131, 1427–1434. [Google Scholar] [CrossRef] [PubMed]
- Klein, B.E.K.; Klein, R.; Lee, K.E.; Grady, L.M. Statin use and incident nuclear cataract. JAMA 2006, 295, 2752–2758. [Google Scholar] [CrossRef] [PubMed]
- Erie, J.C.; Pueringer, M.R.; Brue, S.M.; Chamberlain, A.M.; Hodge, D.O. Statin Use and Incident Cataract Surgery: A Case-Control Study. Ophthalmic Epidemiol. 2016, 23, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Duell, P.B.; Salen, G.; Eichler, F.S.; DeBarber, A.E.; Connor, S.L.; Casaday, L.; Jayadev, S.; Kisanuki, Y.; Lekprasert, P.; Malloy, M.J.; et al. Diagnosis, treatment, and clinical outcomes in 43 cases with cerebrotendinous xanthomatosis. J. Clin. Lipidol. 2018, 12, 1169–1178. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.; Cenedella, R.J. Human lens cholesterol concentrations in patients who used lovastatin or simvastatin. Arch. Ophthalmol. 1999, 117, 653–657. [Google Scholar] [CrossRef]
- Kostis, J.B.; Dobrzynski, J.M. Prevention of cataracts by statins: A meta-analysis. J. Cardiovasc. Pharmacol. Ther. 2014, 19, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Saint-Gerons, D.M.; de la Honrubia, C.F.; Montero, D.; González-Bermejo, D.; de Andrés-Trelles, F.; Catalá-López, F. Statins use and risk of cataracts: Firm conclusions are still far off. J. Cardiovasc. Pharmacol. Ther. 2015, 20, 344–345. [Google Scholar] [CrossRef]
- Hughes, J.R.; Levchenko, V.A.; Blanksby, S.J.; Mitchell, T.W.; Williams, A.; Truscott, R.J. No turnover in lens lipids for the entire human lifespan. eLife 2015, 4, e06003. [Google Scholar] [CrossRef]
- Borchman, D.; Yappert, M.C.; Afzal, M. Lens lipids and maximum lifespan. Exp. Eye Res. 2004, 79, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Borchman, D.; Byrdwell, W.C.; Yappert, M.C. Regional and age-dependent differences in the phospholipid composition of human lens membranes. Investig. Ophthalmol. Vis. Sci. 1994, 35, 3938–3942. [Google Scholar]
- Mainali, L.; Subczynski, W.K. Pure Cholesterol Bilayer Domains are Formed at Cholesterol Contents Significantly Lower than Cholesterol Solubility Thresholds in Phospholipid Membranes: EPR and DSC Studies. Biophys. J. 2018, 114, 450a. [Google Scholar] [CrossRef]
- Huang, J.; Buboltz, J.T.; Feigenson, G.W. Maximum solubility of cholesterol in phosphatidylcholine and phosphatidylethanolamine bilayers. Biochim. Biophys. Acta Biomembr. 1999, 1417, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Mainali, L.; Raguz, M.; Subczynski, W.K. Phases and domains in sphingomyelin-cholesterol membranes: Structure and properties using EPR spin-labeling methods. Eur. Biophys. J. 2012, 41, 147–159. [Google Scholar] [CrossRef]
- Mainali, L.; Raguz, M.; Subczynski, W.K. Formation of cholesterol bilayer domains precedes formation of cholesterol crystals in cholesterol/dimyristoylphosphatidylcholine membranes: EPR and DSC studies. J. Phys. Chem. B 2013, 117, 8994–9003. [Google Scholar] [CrossRef] [PubMed]
- Epand, R.M. Cholesterol in Bilayers of Sphingomyelin or Dihydrosphingomyelin at Concentrations Found in Ocular Lens Membranes. Biophys. J. 2003, 84, 3102–3110. [Google Scholar] [CrossRef] [Green Version]
- Yappert, M.C.; Borchman, D. Sphingolipids in human lens membranes: An update on their composition and possible biological implications. Chem. Phys. Lipids 2004, 129, 1–20. [Google Scholar] [CrossRef]
- Deeley, J.M.; Hankin, J.A.; Friedrich, M.G.; Murphy, R.C.; Truscott, R.J.W.; Mitchell, T.W.; Blanksby, S.J. Sphingolipid distribution changes with age in the human lens. J. Lipid Res. 2010, 51, 2753–2760. [Google Scholar] [CrossRef] [PubMed]
- Broekhuyse, R.M. Phospholipids in tissues of the eye. 3. Composition and metabolism of phospholipids in human lens in relation to age and cataract formation. Biochim. Biophys. Acta 1969, 187, 354–365. [Google Scholar] [CrossRef]
- Goodenough, D.A. The crystalline lens. A system networked by gap junctional intercellular communication. Semin. Cell Biol. 1992, 3, 49–58. [Google Scholar] [CrossRef]
- Kar, R.; Batra, N.; Riquelme, M.A.; Jiang, J.X. Biological Role of Connexin Intercellular Channels and Hemichannels. Arch. Biochem. Biophys. 2012, 524, 2–15. [Google Scholar] [CrossRef] [Green Version]
- Buzhynskyy, N.; Sens, P.; Behar-Cohen, F.; Scheuring, S. Eye lens membrane junctional microdomains: A comparison between healthy and pathological cases. New J. Phys. 2011, 13, 085016. [Google Scholar] [CrossRef]
- Donaldson, P.; Kistler, J.; Mathias, R.T. Molecular Solutions to Mammalian Lens Transparency. Physiology 2001, 16, 118–123. [Google Scholar] [CrossRef]
- Mathias, R.T.; Kistler, J.; Donaldson, P. The lens circulation. J. Membr. Biol. 2007, 216, 1–16. [Google Scholar] [CrossRef]
- Subczynski, W.K.; Widomska, J.; Feix, J.B. Physical properties of lipid bilayers from EPR spin labeling and their influence on chemical reactions in a membrane environment. Free Radic. Biol. Med. 2009, 46, 707–718. [Google Scholar] [CrossRef]
- Palmquist, B.M.; Philipson, B.; Barr, P.O. Nuclear cataract and myopia during hyperbaric oxygen therapy. Br. J. Ophthalmol. 1984, 68, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Gengel, K.C.; Cooper, J.S. Hyperbaric Related Myopia and Cataract Formation. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Holekamp, N.M.; Shui, Y.-B.; Beebe, D.C. Vitrectomy surgery increases oxygen exposure to the lens: A possible mechanism for nuclear cataract formation. Am. J. Ophthalmol. 2005, 139, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Truscott, R.J.W. Age-related nuclear cataract-oxidation is the key. Exp. Eye Res. 2005, 80, 709–725. [Google Scholar] [CrossRef] [PubMed]
- Siegfried, C.J.; Shui, Y.-B.; Holekamp, N.M.; Bai, F.; Beebe, D.C. Oxygen distribution in the human eye: Relevance to the etiology of open-angle glaucoma after vitrectomy. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5731–5738. [Google Scholar] [CrossRef] [PubMed]
- McNulty, R.; Wang, H.; Mathias, R.T.; Ortwerth, B.J.; Truscott, R.J.W.; Bassnett, S. Regulation of tissue oxygen levels in the mammalian lens. J. Physiol. 2004, 559, 883–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shui, Y.-B.; Holekamp, N.M.; Kramer, B.C.; Crowley, J.R.; Wilkins, M.A.; Chu, F.; Malone, P.E.; Mangers, S.J.; Hou, J.H.; Siegfried, C.J.; et al. The gel state of the vitreous and ascorbate-dependent oxygen consumption: Relationship to the etiology of nuclear cataracts. Arch. Ophthalmol. 2009, 127, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Beebe, D.C.; Holekamp, N.M.; Siegfried, C.; Shui, Y.-B. Vitreoretinal influences on lens function and cataract. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2011, 366, 1293–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raguz, M.; Mainali, L.; O’Brien, W.J.; Subczynski, W.K. Lipid Domains in Intact Fiber-Cell Plasma Membranes Isolated from Cortical and Nuclear Regions of Human Eye Lenses of Donors from Different Age Groups. Exp. Eye Res. 2015, 132, 78–90. [Google Scholar] [CrossRef] [Green Version]
- Plesnar, E.; Szczelina, R.; Subczynski, W.K.; Pasenkiewicz-Gierula, M. Is the cholesterol bilayer domain a barrier to oxygen transport into the eye lens? Biochim. Biophys. Acta Biomembr. 2018, 1860, 434–441. [Google Scholar] [CrossRef]
- Freigang, S.; Ampenberger, F.; Spohn, G.; Heer, S.; Shamshiev, A.T.; Kisielow, J.; Hersberger, M.; Yamamoto, M.; Bachmann, M.F.; Kopf, M. Nrf2 is essential for cholesterol crystal-induced inflammasome activation and exacerbation of atherosclerosis. Eur. J. Immunol. 2011, 41, 2040–2051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabas, I. Consequences of cellular cholesterol accumulation: Basic concepts and physiological implications. J. Clin. Investig. 2002, 110, 905–911. [Google Scholar] [CrossRef] [PubMed]
- Dostert, C.; Pétrilli, V.; Van Bruggen, R.; Steele, C.; Mossman, B.T.; Tschopp, J. Innate immune activation through Nalp3 inflammasome sensing of asbestos and silica. Science 2008, 320, 674–677. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.G.; Kuwabara, M.; Johnson, R.; Bove, M.; Fogacci, F.; Rosticci, M.; Giovannini, M.; D’Addato, S.; Borghi, C. Brisighella Heart Study group LDL-oxidation, serum uric acid, kidney function and pulse-wave velocity: Data from the Brisighella Heart Study cohort. Int. J. Cardiol. 2018, 261, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.G.; Fogacci, F.; Giovannini, M.; Grandi, E.; D’Addato, S.; Borghi, C. Brisighella Heart Study group Interaction between low-density lipoprotein-cholesterolaemia, serum uric level and incident hypertension: Data from the Brisighella Heart Study. J. Hypertens. 2019, 37, 728–731. [Google Scholar] [CrossRef] [PubMed]
- Kuwabara, M.; Kanbay, M.; Hisatome, I. Uric Acid and Hypertension Because of Arterial Stiffness. Hypertension 2018, 72, 582–584. [Google Scholar] [CrossRef] [PubMed]
- Duewell, P.; Kono, H.; Rayner, K.J.; Sirois, C.M.; Vladimer, G.; Bauernfeind, F.G.; Abela, G.S.; Franchi, L.; Nuñez, G.; Schnurr, M.; et al. NLRP3 inflammasomes are required for atherogenesis and activated by cholesterol crystals. Nature 2010, 464, 1357–1361. [Google Scholar] [CrossRef] [PubMed]
- Tulenko, T.N.; Chen, M.; Mason, P.E.; Mason, R.P. Physical effects of cholesterol on arterial smooth muscle membranes: Evidence of immiscible cholesterol domains and alterations in bilayer width during atherogenesis. J. Lipid Res. 1998, 39, 947–956. [Google Scholar] [PubMed]
- Weber, C.; Noels, H. Atherosclerosis: Current pathogenesis and therapeutic options. Nat. Med. 2011, 17, 1410–1422. [Google Scholar] [CrossRef]
- Tabas, I. Macrophage death and defective inflammation resolution in atherosclerosis. Nat. Rev. Immunol. 2010, 10, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Small, D.M. George Lyman Duff memorial lecture. Progression and regression of atherosclerotic lesions. Insights from lipid physical biochemistry. Arteriosclerosis 1988, 8, 103–129. [Google Scholar] [CrossRef] [PubMed]
- Stary, H.C.; Chandler, A.B.; Dinsmore, R.E.; Fuster, V.; Glagov, S.; Insull, W.; Rosenfeld, M.E.; Schwartz, C.J.; Wagner, W.D.; Wissler, R.W. A definition of advanced types of atherosclerotic lesions and a histological classification of atherosclerosis. A report from the Committee on Vascular Lesions of the Council on Arteriosclerosis, American Heart Association. Circulation 1995, 92, 1355–1374. [Google Scholar] [CrossRef]
- Abela, G.S. Cholesterol crystals piercing the arterial plaque and intima trigger local and systemic inflammation. J. Clin. Lipidol. 2010, 4, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Abela, G.S.; Aziz, K. Cholesterol crystals cause mechanical damage to biological membranes: A proposed mechanism of plaque rupture and erosion leading to arterial thrombosis. Clin. Cardiol. 2005, 28, 413–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abela, G.S.; Aziz, K. Cholesterol crystals rupture biological membranes and human plaques during acute cardiovascular events—A novel insight into plaque rupture by scanning electron microscopy. Scanning 2006, 28, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Vedre, A.; Pathak, D.R.; Crimp, M.; Lum, C.; Koochesfahani, M.; Abela, G.S. Physical factors that trigger cholesterol crystallization leading to plaque rupture. Atherosclerosis 2009, 203, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Jacob, R.F.; Mason, R.P. Lipid Peroxidation Induces Cholesterol Domain Formation in Model Membranes. J. Biol. Chem. 2005, 280, 39380–39387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eder, M.I.; Miquel, J.F.; Jongst, D.; Paumgartner, G.; von Ritter, C. Reactive oxygen metabolites promote cholesterol crystal formation in model bile: Role of lipid peroxidation. Free Radic. Biol. Med. 1996, 20, 743–749. [Google Scholar] [CrossRef]
- Mainali, L.; Zareba, M.; Subczynski, W.K. Oxidation of Polyunsaturated Phospholipid Decreases the Cholesterol Content at which Cholesterol Bilayer Domains Start to form in Phospholipid-Cholesterol Membranes. Biophys. J. 2017, 112, 375a. [Google Scholar] [CrossRef]
- Wassall, S.R.; Stillwell, W. Polyunsaturated fatty acid-cholesterol interactions: Domain formation in membranes. Biochim. Biophys. Acta 2009, 1788, 24–32. [Google Scholar] [CrossRef]
- Brzustowicz, M.R.; Cherezov, V.; Caffrey, M.; Stillwell, W.; Wassall, S.R. Molecular organization of cholesterol in polyunsaturated membranes: Microdomain formation. Biophys. J. 2002, 82, 285–298. [Google Scholar] [CrossRef]
- Dietschy, J.M.; Turley, S.D.; Spady, D.K. Role of liver in the maintenance of cholesterol and low density lipoprotein homeostasis in different animal species, including humans. J. Lipid Res. 1993, 34, 1637–1659. [Google Scholar] [PubMed]
- Dietschy, J.M.; Turley, S.D. Thematic review series: Brain Lipids. Cholesterol metabolism in the central nervous system during early development and in the mature animal. J. Lipid Res. 2004, 45, 1375–1397. [Google Scholar] [CrossRef] [PubMed]
- Muldoon, M.F.; Ryan, C.M.; Sereika, S.M.; Flory, J.D.; Manuck, S.B. Randomized trial of the effects of simvastatin on cognitive functioning in hypercholesterolemic adults. Am. J. Med. 2004, 117, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Posvar, E.L.; Radulovic, L.L.; Cilla, D.D.; Whitfield, L.R.; Sedman, A.J. Tolerance and pharmacokinetics of single-dose atorvastatin, a potent inhibitor of HMG-CoA reductase, in healthy subjects. J. Clin. Pharmacol. 1996, 36, 728–731. [Google Scholar] [CrossRef]
- Benito-León, J.; Louis, E.D.; Vega, S.; Bermejo-Pareja, F. Statins and cognitive functioning in the elderly: A population-based study. J. Alzheimers Dis. 2010, 21, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Richardson, K.; Schoen, M.; French, B.; Umscheid, C.A.; Mitchell, M.D.; Arnold, S.E.; Heidenreich, P.A.; Rader, D.J.; deGoma, E.M. Statins and cognitive function: A systematic review. Ann. Intern. Med. 2013, 159, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Wolozin, B.; Kellman, W.; Ruosseau, P.; Celesia, G.G.; Siegel, G. Decreased prevalence of Alzheimer disease associated with 3-hydroxy-3-methyglutaryl coenzyme A reductase inhibitors. Arch. Neurol. 2000, 57, 1439–1443. [Google Scholar] [CrossRef]
- Jick, H.; Zornberg, G.L.; Jick, S.S.; Seshadri, S.; Drachman, D.A. Statins and the risk of dementia. Lancet 2000, 356, 1627–1631. [Google Scholar] [CrossRef]
- Wong, W.B.; Lin, V.W.; Boudreau, D.; Devine, E.B. Statins in the prevention of dementia and Alzheimer’s disease: A meta-analysis of observational studies and an assessment of confounding. Pharmacoepidemiol. Drug Saf. 2013, 22, 345–358. [Google Scholar] [CrossRef]
- McGuinness, B.; Craig, D.; Bullock, R.; Passmore, P. Statins for the prevention of dementia. Cochrane Database Syst. Rev. 2016. [Google Scholar] [CrossRef] [PubMed]
- Appleton, J.P.; Scutt, P.; Sprigg, N.; Bath, P.M. Hypercholesterolaemia and vascular dementia. Clin. Sci. 2017, 131, 1561–1578. [Google Scholar] [CrossRef] [Green Version]
- Schultz, B.G.; Patten, D.K.; Berlau, D.J. The role of statins in both cognitive impairment and protection against dementia: A tale of two mechanisms. Transl. Neurodegener. 2018, 7. [Google Scholar] [CrossRef]
- Subczynski, W.K.; Pasenkiewicz-Gierula, M.; Widomska, J.; Mainali, L.; Raguz, M. High Cholesterol/Low Cholesterol: Effects in Biological Membranes: A Review. Cell Biochem. Biophys. 2017, 75, 369–385. [Google Scholar] [CrossRef] [PubMed]
- Nordestgaard, B.G.; Chapman, M.J.; Humphries, S.E.; Ginsberg, H.N.; Masana, L.; Descamps, O.S.; Wiklund, O.; Hegele, R.A.; Raal, F.J.; Defesche, J.C.; et al. Familial hypercholesterolaemia is underdiagnosed and undertreated in the general population: Guidance for clinicians to prevent coronary heart disease: Consensus statement of the European Atherosclerosis Society. Eur. Heart J. 2013, 34, 3478–3490a. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, E.D.; Watts, G.F.; Fatemi-Langroudi, B.; Gosling, R.G. Aortic compliance in young patients with heterozygous familial hypercholesterolaemia. Clin. Sci. 1992, 83, 717–721. [Google Scholar] [CrossRef] [PubMed]
- Tsoupras, A.; Lordan, R.; Zabetakis, I. Inflammation, not Cholesterol, Is a Cause of Chronic Disease. Nutrients 2018, 10, 604. [Google Scholar] [CrossRef]
- Solomonov, I.; Weygand, M.J.; Kjaer, K.; Rapaport, H.; Leiserowitz, L. Trapping Crystal Nucleation of Cholesterol Monohydrate: Relevance to Pathological Crystallization. Biophys. J. 2005, 88, 1809–1817. [Google Scholar] [CrossRef] [PubMed]
- Varsano, N.; Fargion, I.; Wolf, S.G.; Leiserowitz, L.; Addadi, L. Formation of 3D Cholesterol Crystals from 2D Nucleation Sites in Lipid Bilayer Membranes: Implications for Atherosclerosis. J. Am. Chem. Soc. 2015, 137, 1601–1607. [Google Scholar] [CrossRef]
- Ziblat, R.; Fargion, I.; Leiserowitz, L.; Addadi, L. Spontaneous formation of two-dimensional and three-dimensional cholesterol crystals in single hydrated lipid bilayers. Biophys. J. 2012, 103, 255–264. [Google Scholar] [CrossRef]
- Ong, D.S.; Anzinger, J.J.; Leyva, F.J.; Rubin, N.; Addadi, L.; Kruth, H.S. Extracellular cholesterol-rich microdomains generated by human macrophages and their potential function in reverse cholesterol transport. J. Lipid Res. 2010, 51, 2303–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troup, G.M.; Xie, Y.; Boesze-Battaglia, K.; Huang, Y.; Kirk, T.; Hanley, F.; Tulenko, T.N. Membrane Cholesterol and the Formation of Cholesterol Domains in the Pathogenesis of Cardiovascular Disease. Macromol. Symp. 2005, 219, 25–38. [Google Scholar] [CrossRef]
- Kellner-Weibel, G.; Yancey, P.G.; Jerome, W.G.; Walser, T.; Mason, R.P.; Phillips, M.C.; Rothblat, G.H. Crystallization of free cholesterol in model macrophage foam cells. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 1891–1898. [Google Scholar] [CrossRef] [PubMed]
- Bassnett, S. Lens organelle degradation. Exp. Eye Res. 2002, 74, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wride, M.A. Lens fibre cell differentiation and organelle loss: Many paths lead to clarity. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2011, 366, 1219–1233. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Widomska, J.; Subczynski, W.K. Why Is Very High Cholesterol Content Beneficial for the Eye Lens but Negative for Other Organs? Nutrients 2019, 11, 1083. https://doi.org/10.3390/nu11051083
Widomska J, Subczynski WK. Why Is Very High Cholesterol Content Beneficial for the Eye Lens but Negative for Other Organs? Nutrients. 2019; 11(5):1083. https://doi.org/10.3390/nu11051083
Chicago/Turabian StyleWidomska, Justyna, and Witold K. Subczynski. 2019. "Why Is Very High Cholesterol Content Beneficial for the Eye Lens but Negative for Other Organs?" Nutrients 11, no. 5: 1083. https://doi.org/10.3390/nu11051083
APA StyleWidomska, J., & Subczynski, W. K. (2019). Why Is Very High Cholesterol Content Beneficial for the Eye Lens but Negative for Other Organs? Nutrients, 11(5), 1083. https://doi.org/10.3390/nu11051083