Effects of Moderate Chronic Food Restriction on the Development of Postprandial Dyslipidemia with Ageing

 , , , and
, , , and
Abstract
:1. Introduction
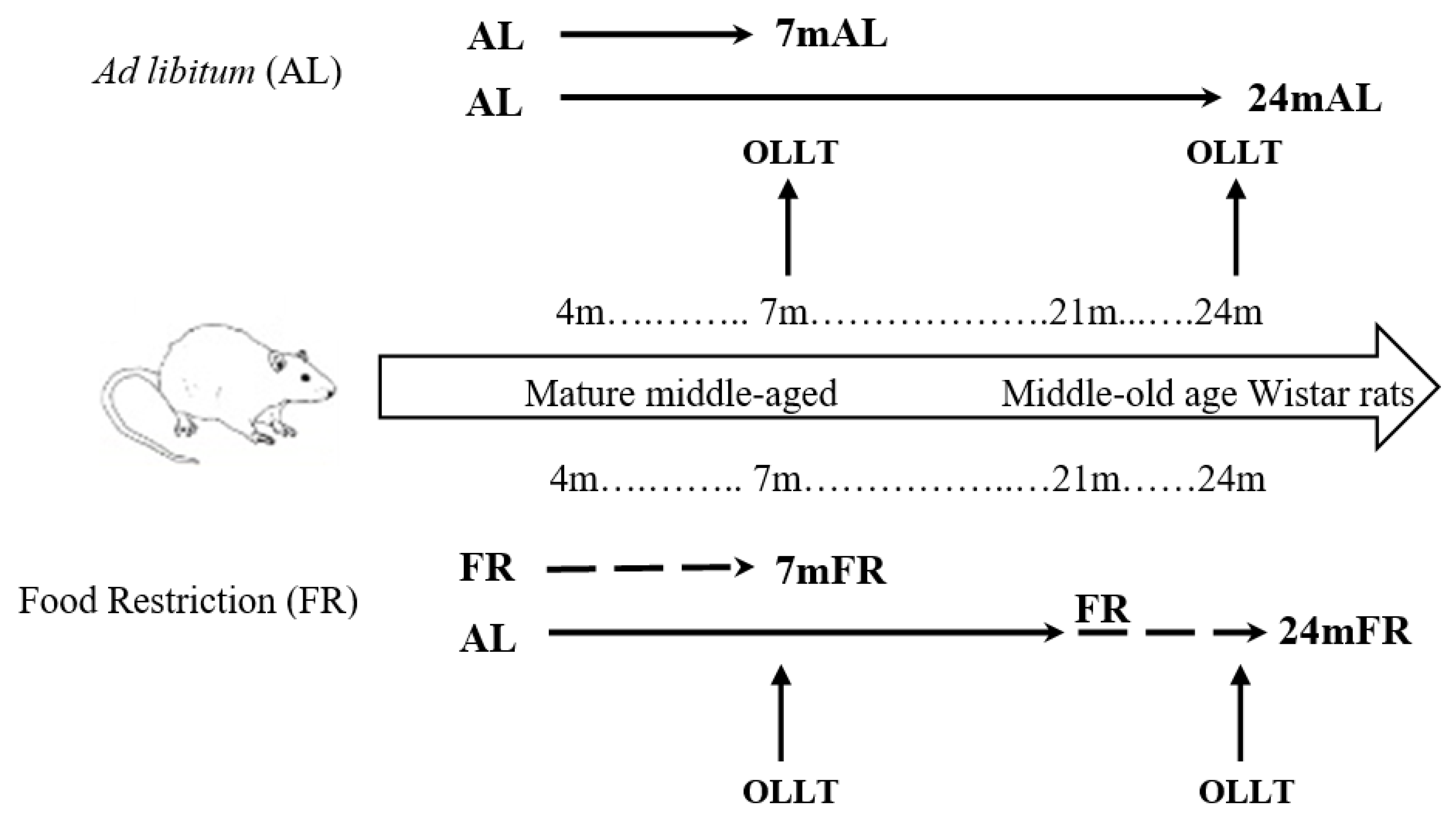
2. Materials and Methods
2.1. Animals
2.2. Food Restriction (FR) Protocol
2.3. Oral Lipid Loading Test (OLLT)
2.4. Serum and Tissue Measurements of Hormones and Metabolites
2.5. Real-Time RT-PCR
2.6. Calculations and Statistical Analysis
3. Results
3.1. Physiological and Metabolic Parameters in the Fasting State
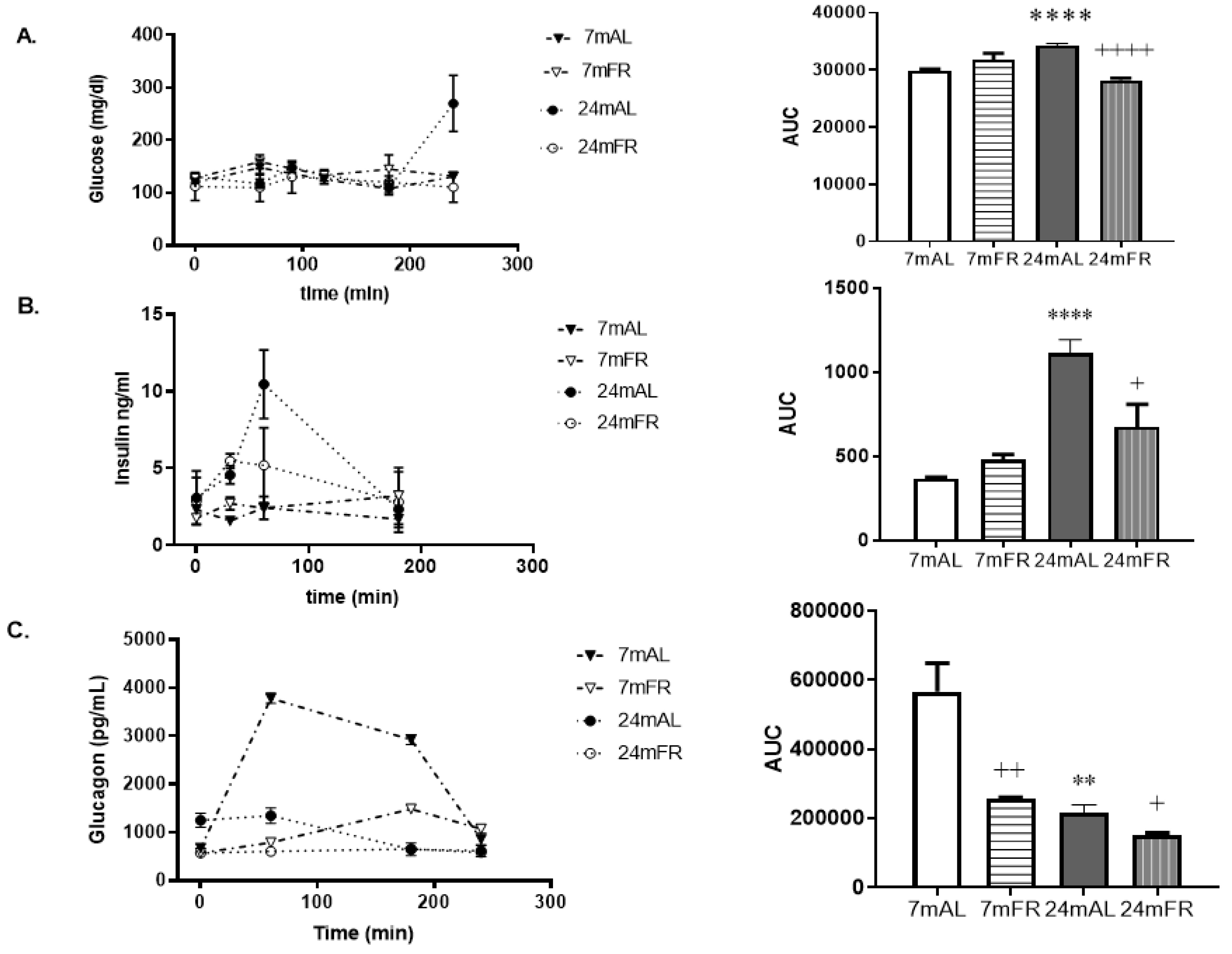
3.2. Effect of Food Restriction in Serum Glucose, Insulin, and Glucagon Profiles during the OLLT
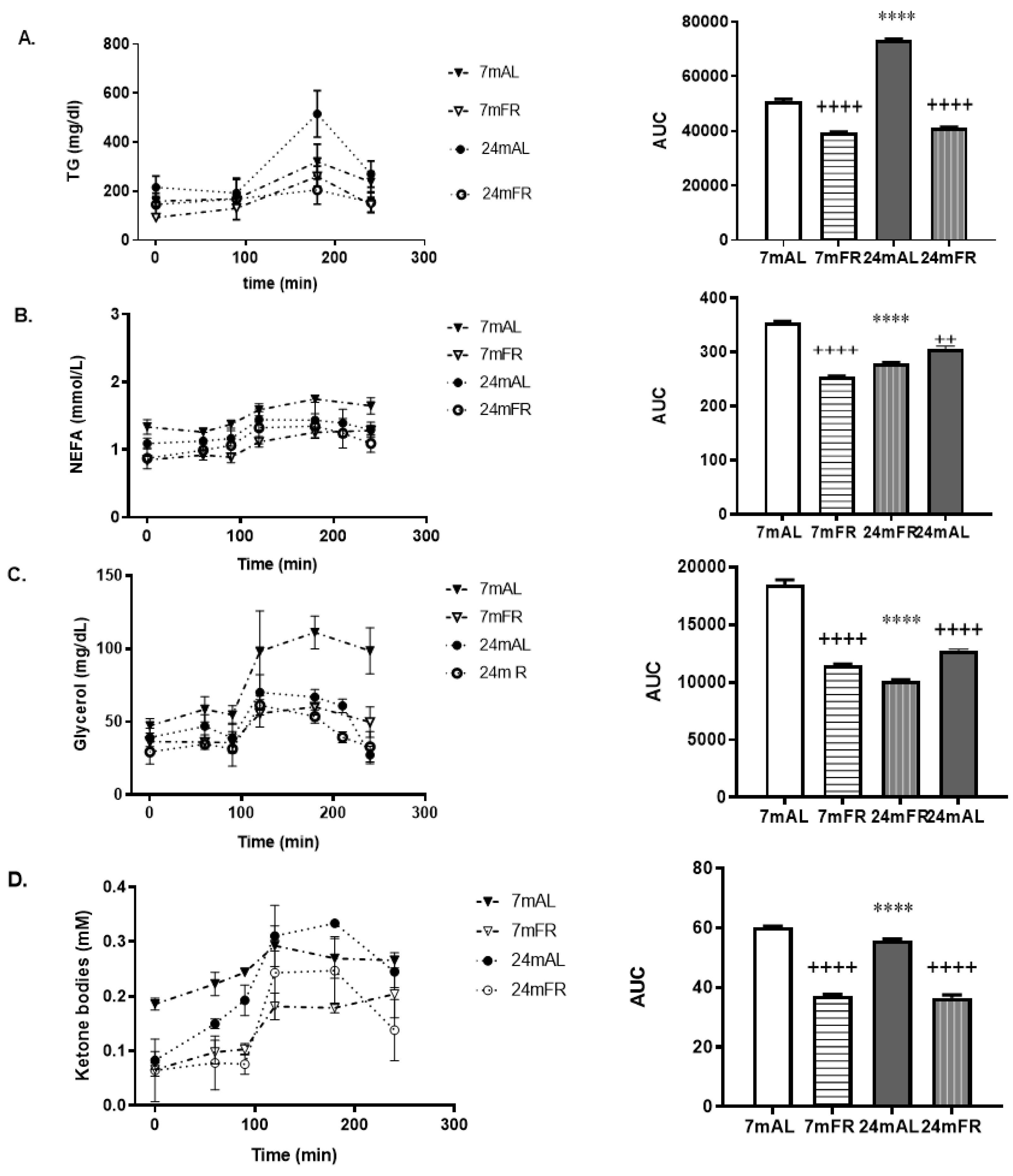
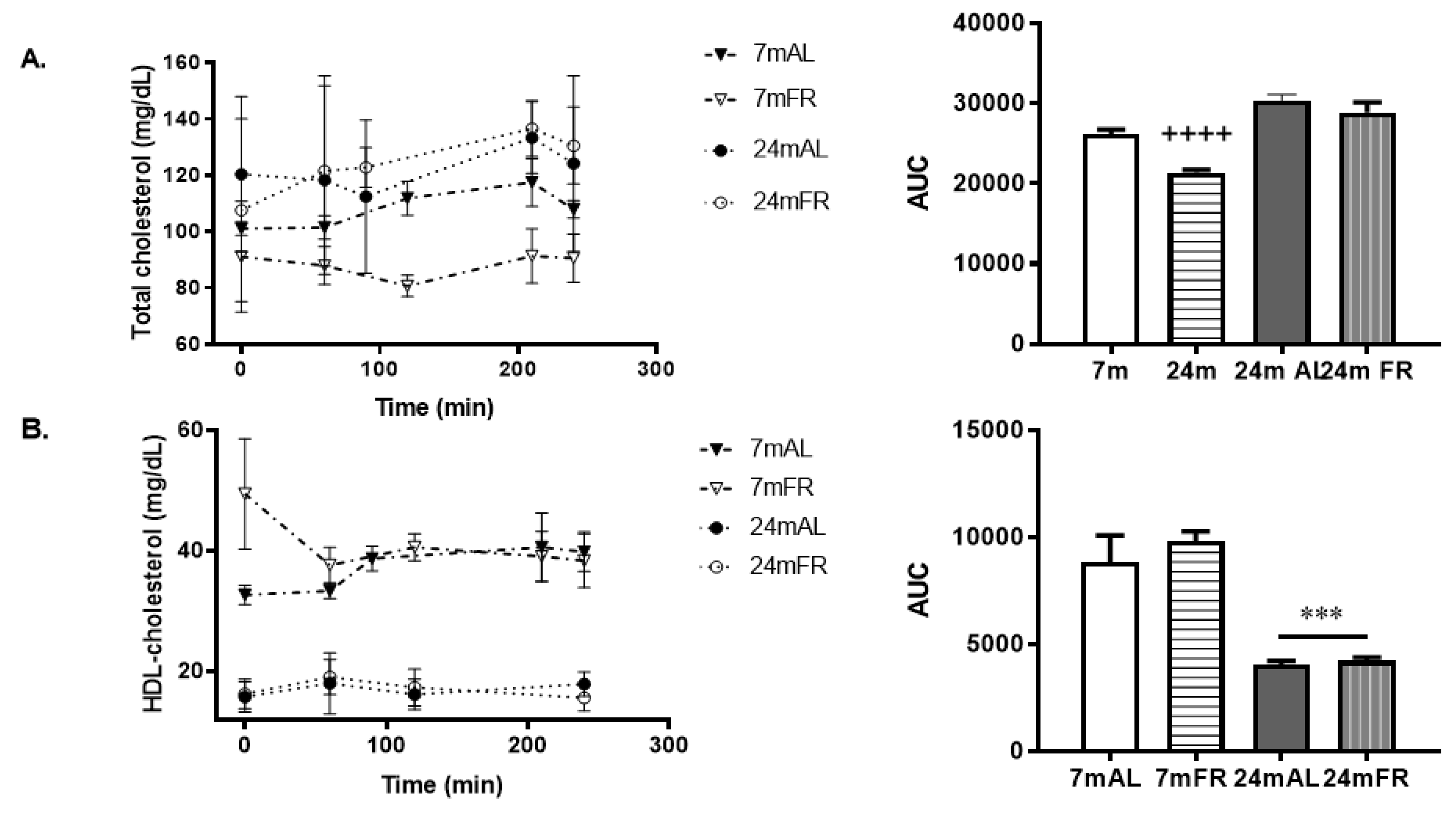
3.3. Serum Lipid Profiles during the OLLT
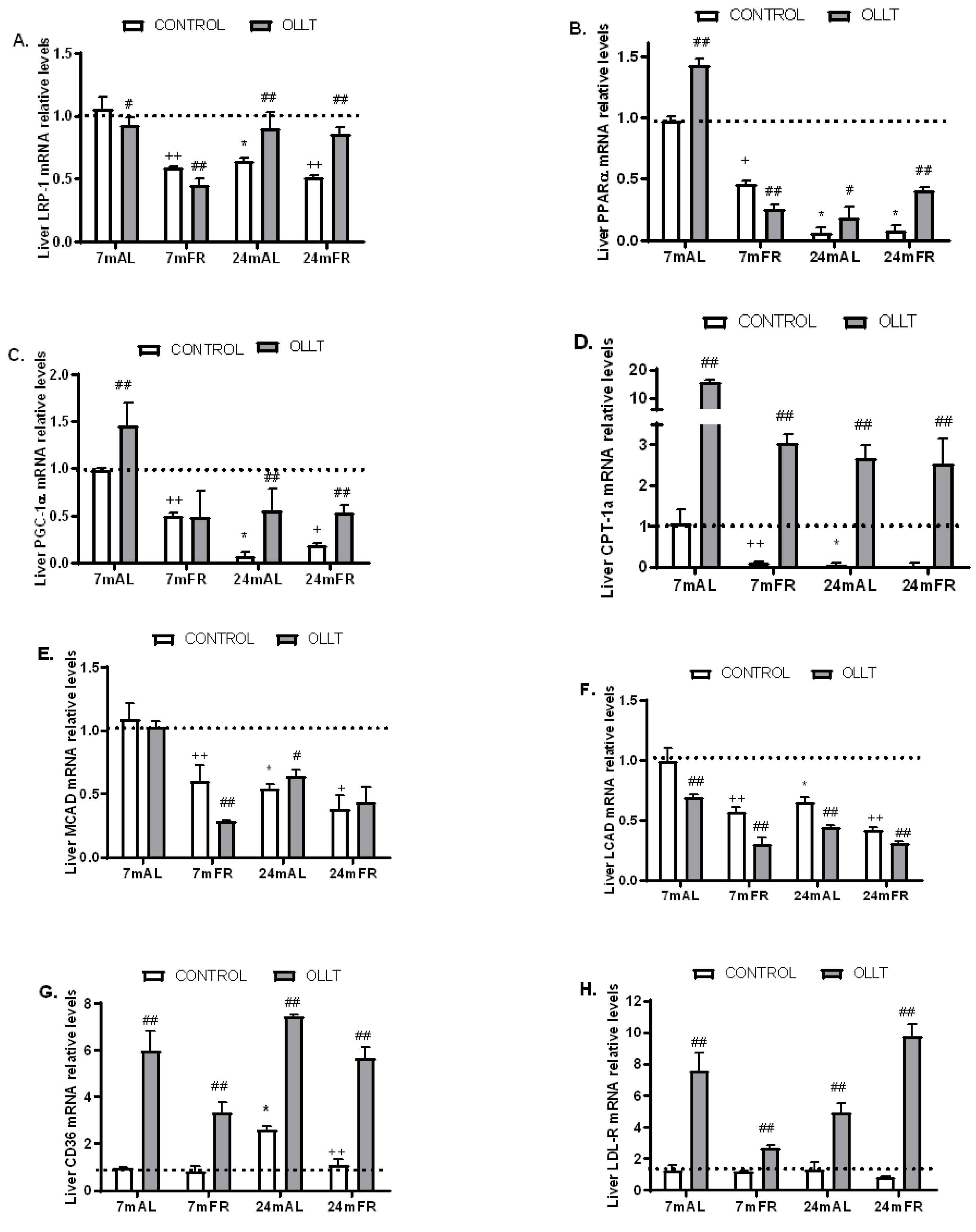
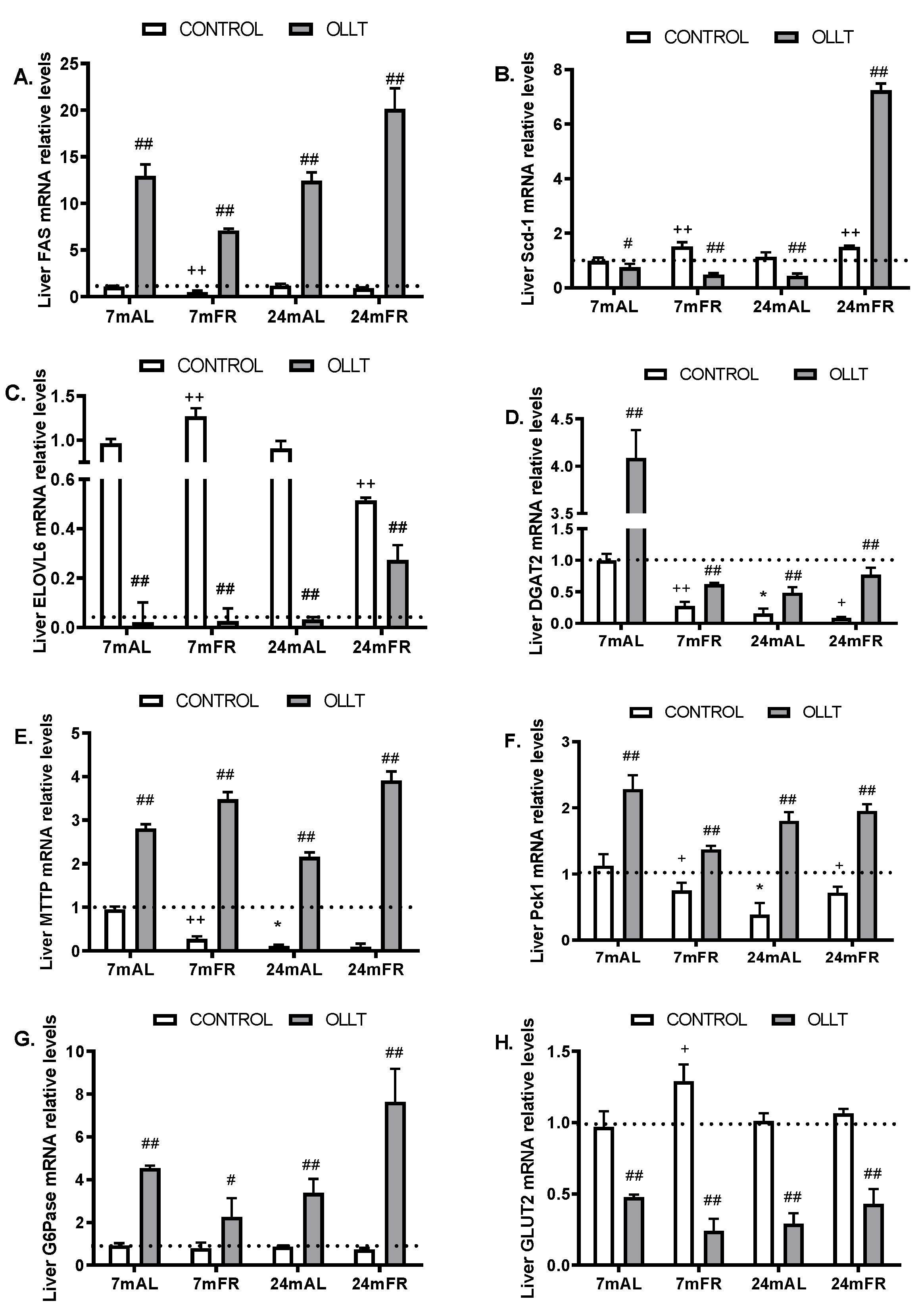
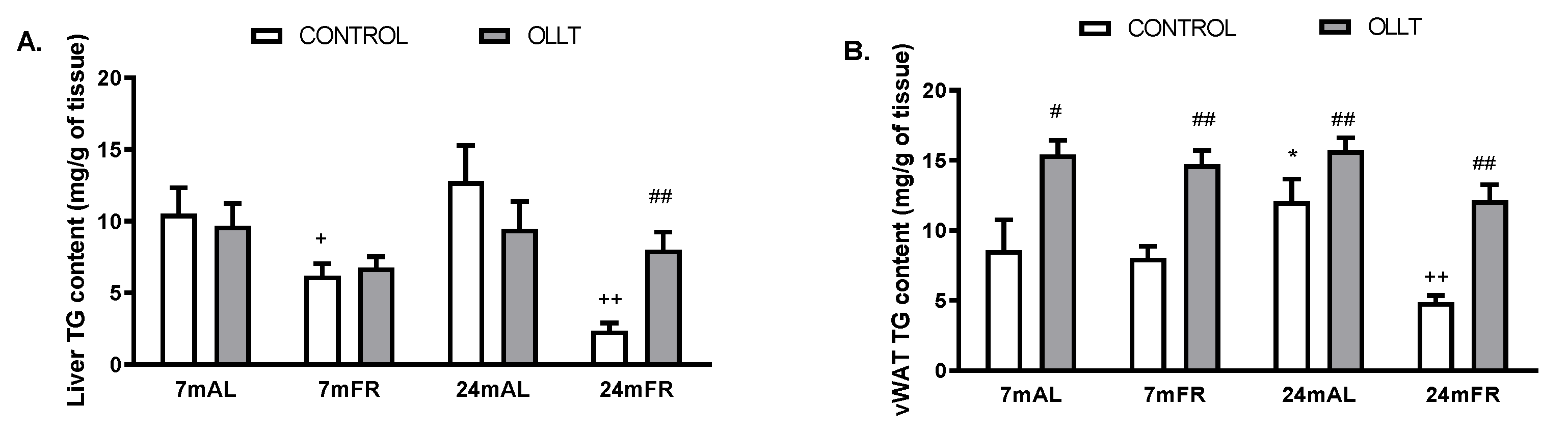
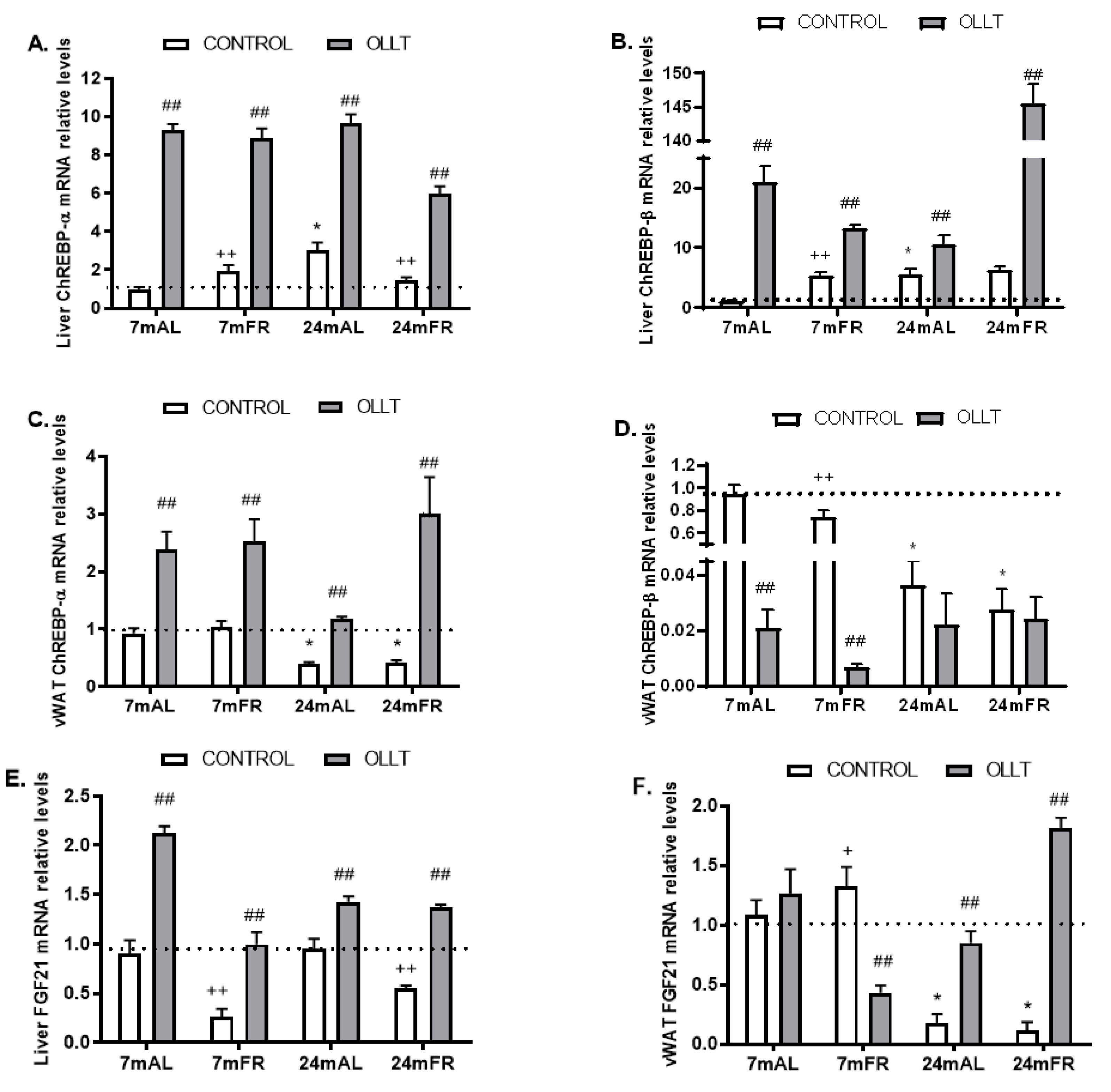
3.4. Changes in the Metabolic Capacity of Liver after the OLLT
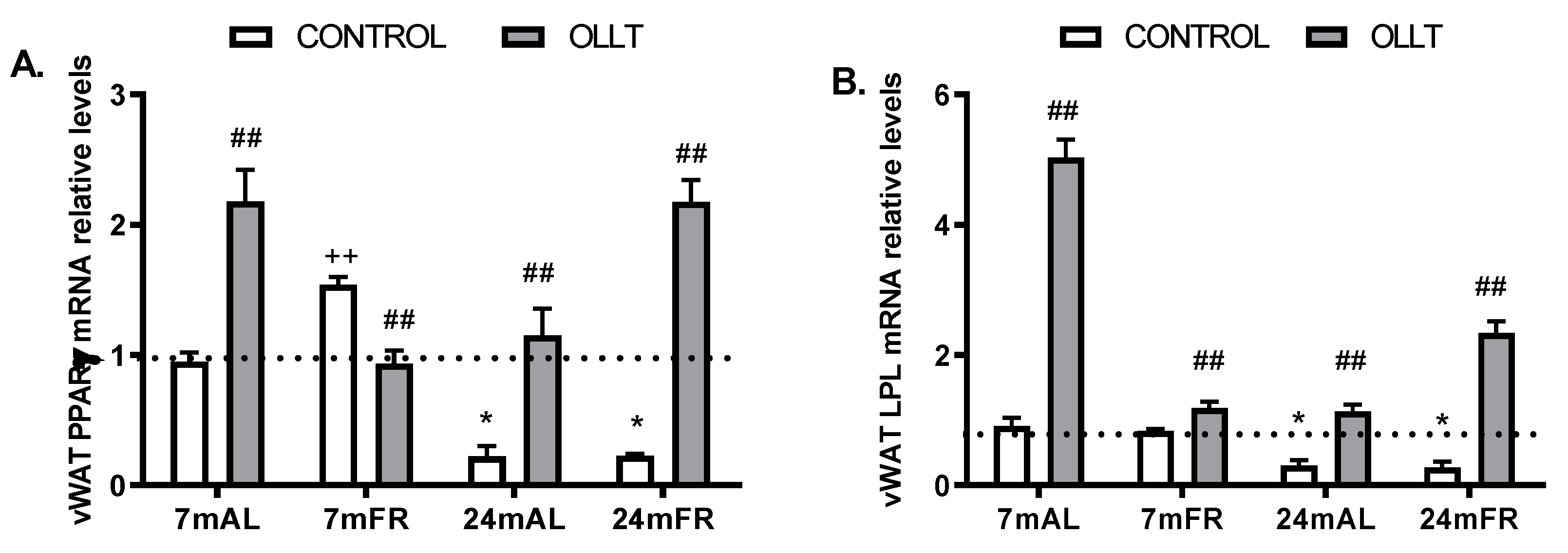
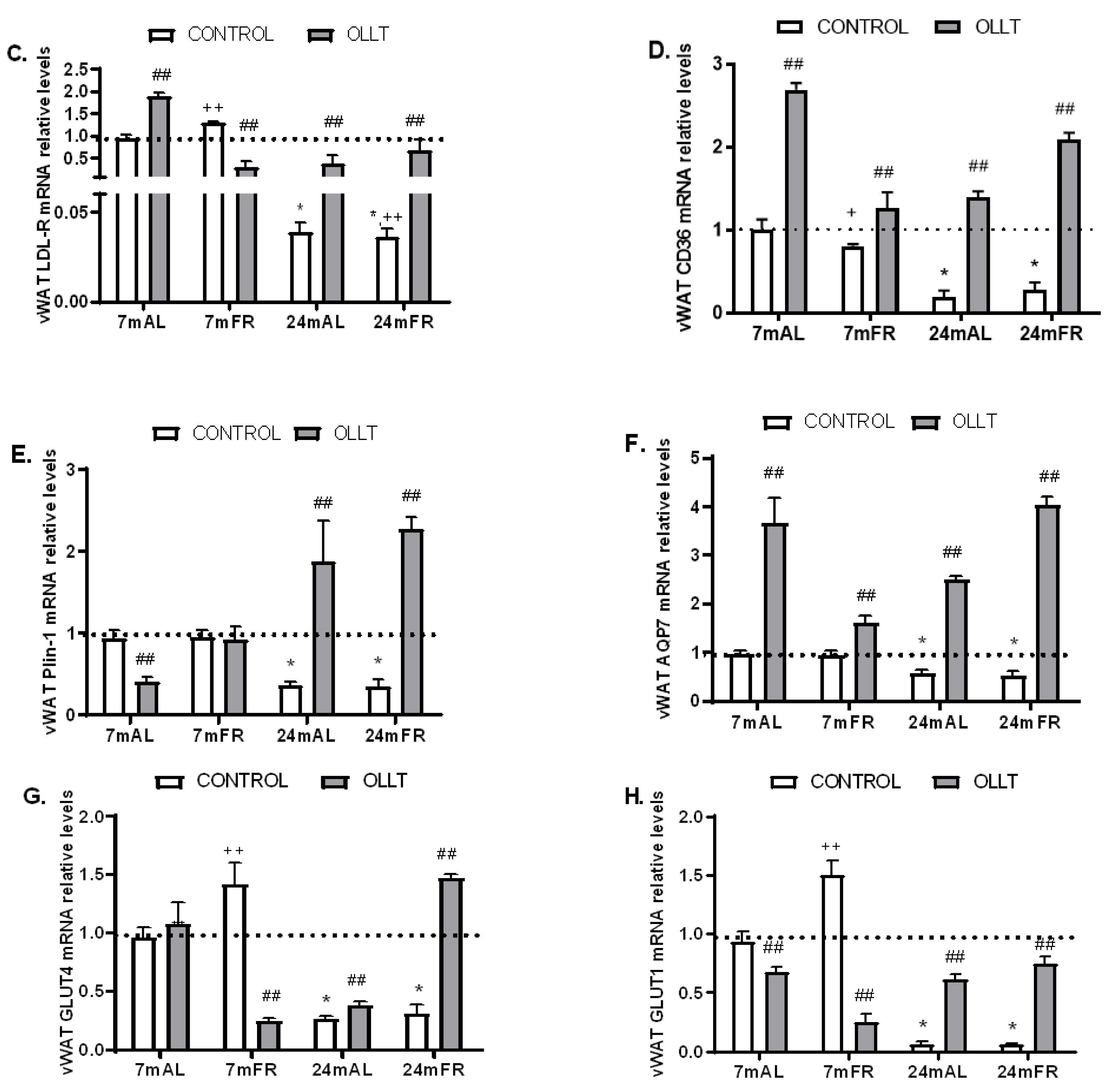
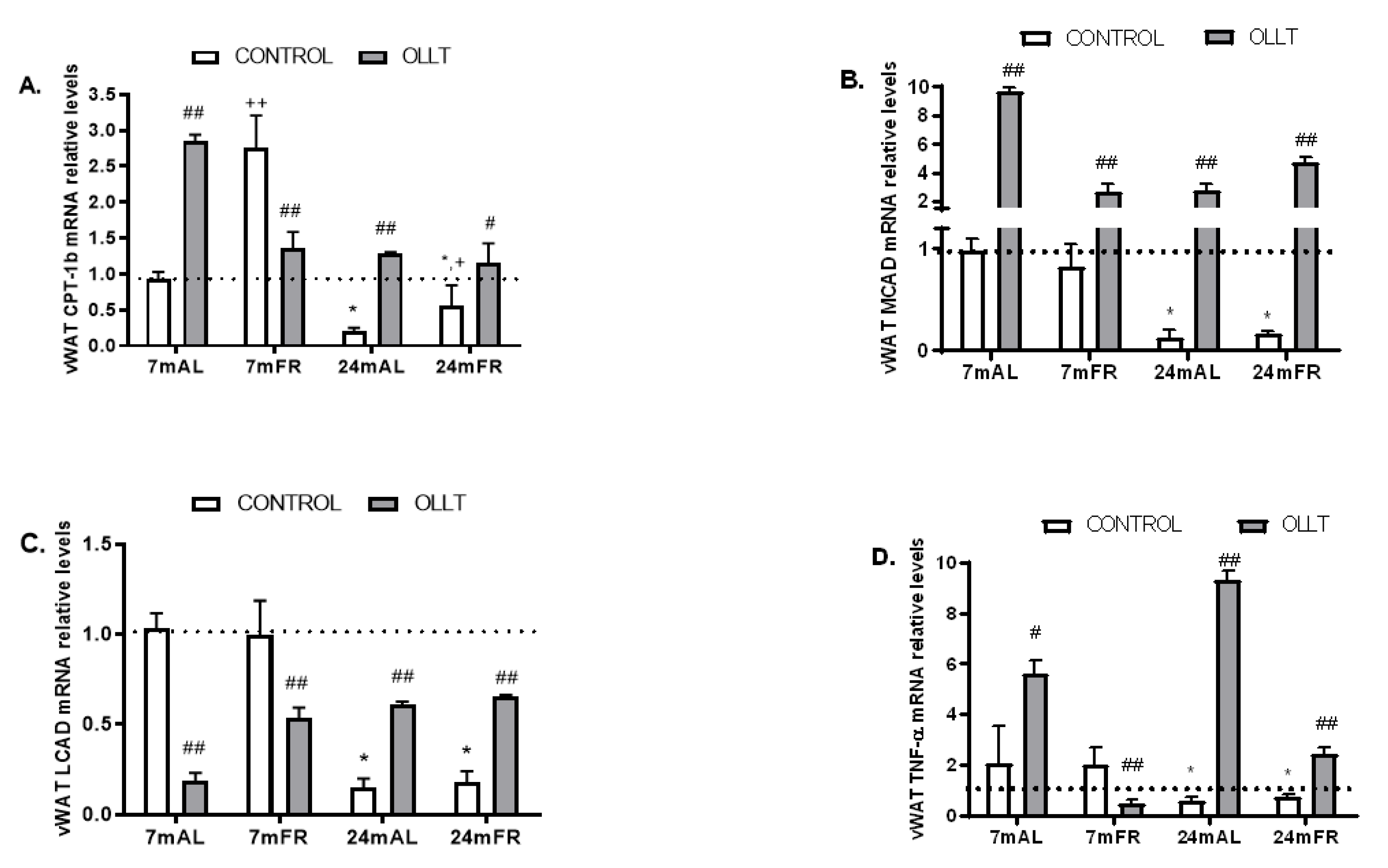
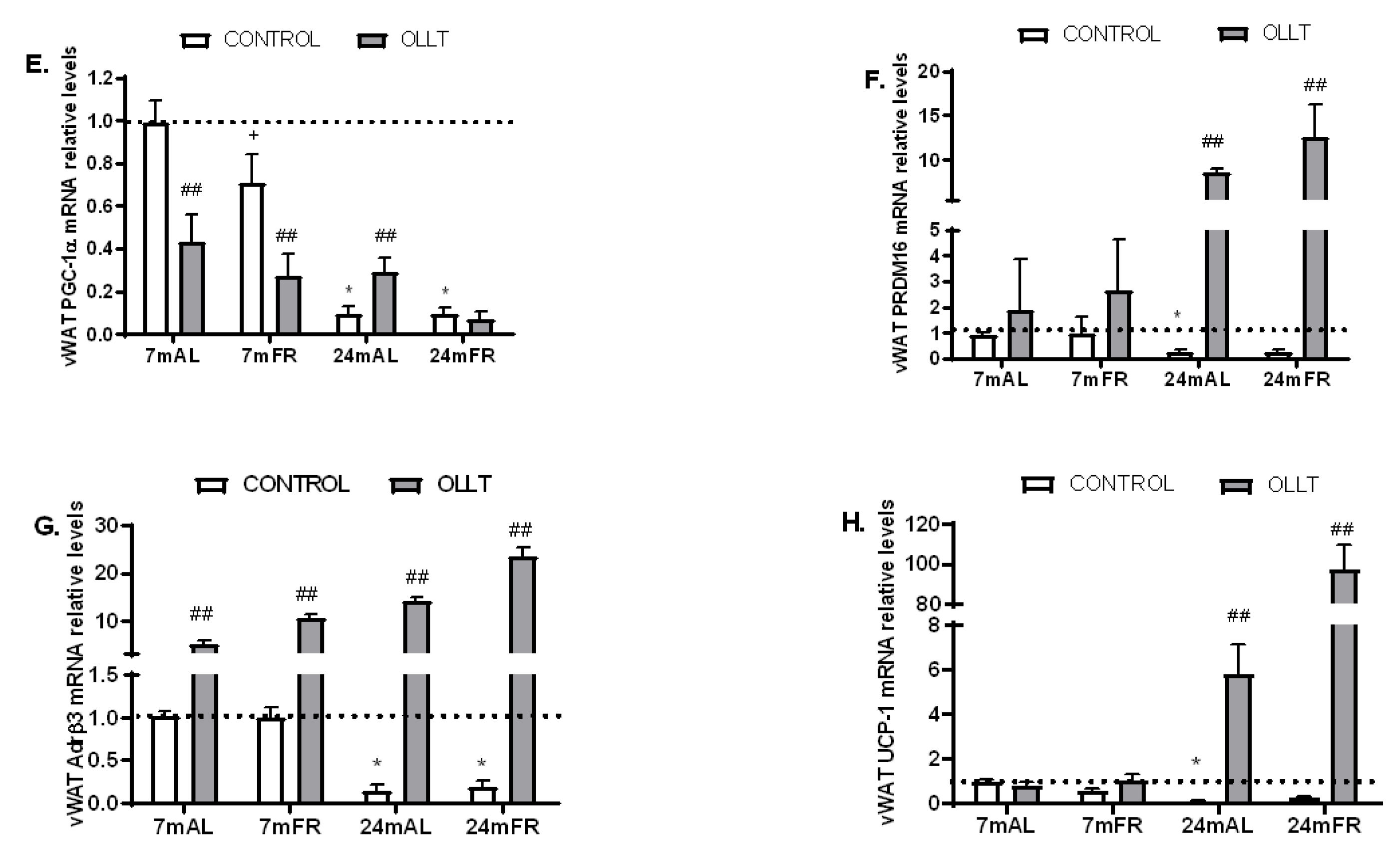
3.5. Changes in the Metabolic Capacity of vWAT after the OLLT
3.6. Changes in Hepatic and vWAT ChREBP Activity after the OLLT
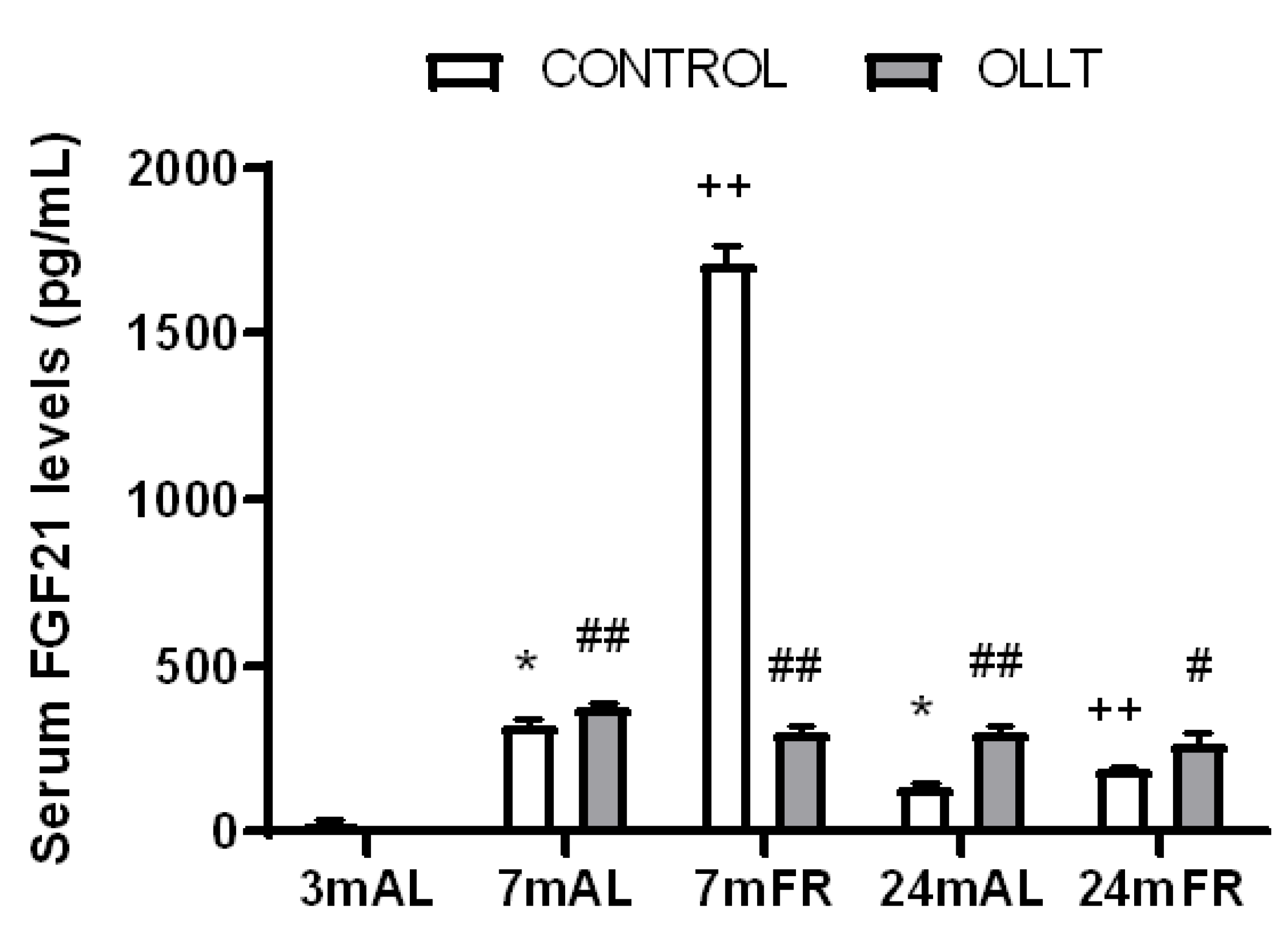
3.7. Changes in the Basal and Postprandial Serum Levels of FGF21 after the OLLT
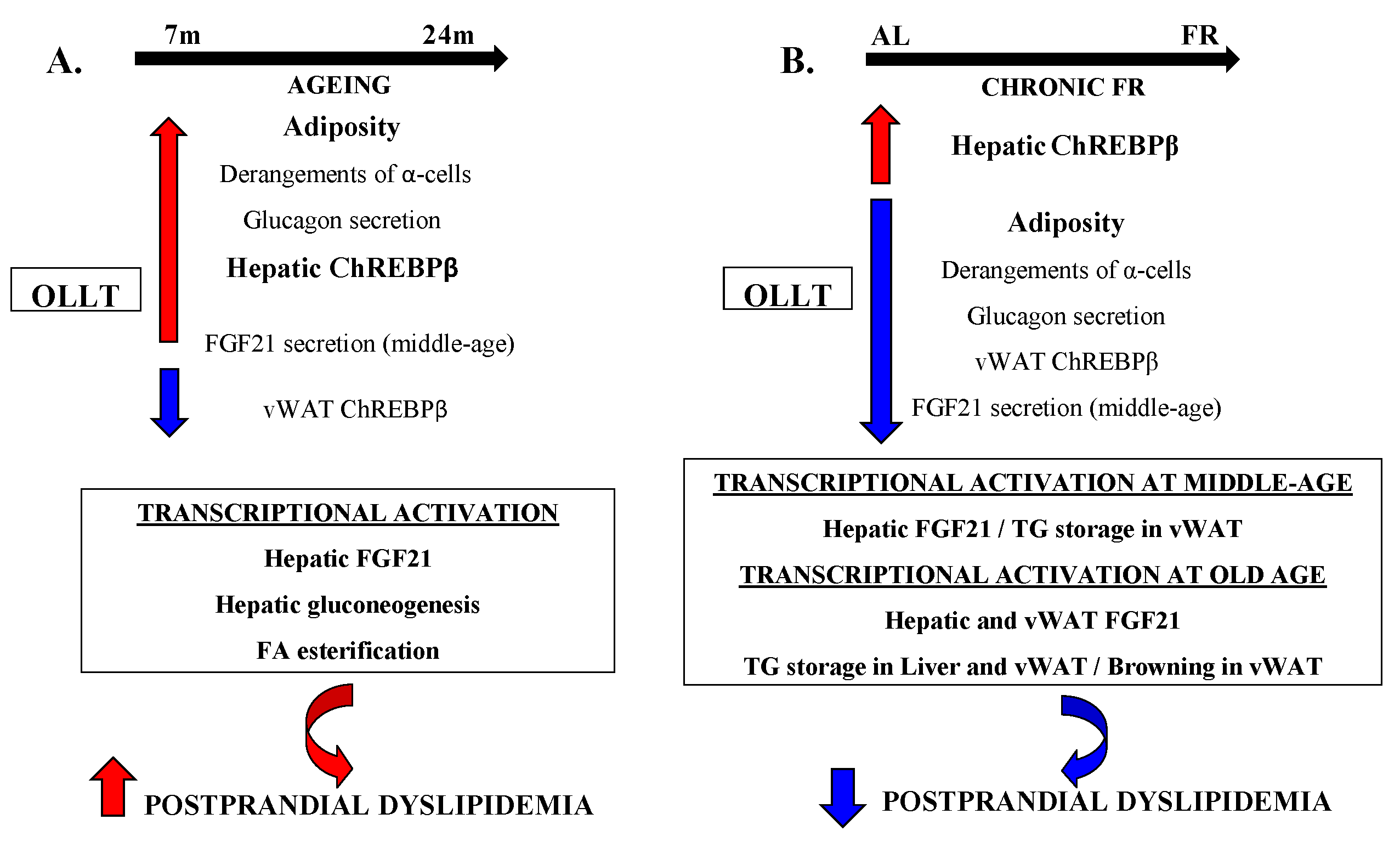
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bennett, B.J.; Hall, K.D.; Hu, F.B.; McCarteny, A.L.; Roberto, C. Nutrition and the science of disease prevention: A systems approach to support metabolic health. Ann. N.Y. Acad. Sci. 2015, 1352, 1–12. [Google Scholar] [CrossRef] [PubMed]
- McPhail, S.J. Multimorbidity in chronic disease: Impact on health care resources and costs. Risk Manag. Healthc. Policy 2016, 9, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Reiner, Z.; Catapano, A.L.; De Backer, G.; Graham, I.; Taskinen, M.R.; Wiklund, O.; Agewall, S.; Alegria, E.; Chapman, M.J.; Durrington, P.; et al. ESC/EAS Guidelines for the management of dyslipidaemias: The Task Force for the management of dyslipidaemias of the European Society of Cardiology (ESC) and the European Atherosclerosis Society (EAS). Eur. Heart J. 2011, 32, 1769–1818. [Google Scholar] [PubMed] [Green Version]
- Krasinski, S.D.; Cohn, J.S.; Schaefer, E.J.; Russell, R.M. Postprandial plasma retinyl ester response is greater in older subjects compared with younger subjects. Evidence for delayed plasma clearance of intestinal lipoproteins. J. Clin. Invest. 1990, 85, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Zilversmit, D.B. Atherogenesis: A post-prandial phenomenon. Circulation 1979, 60, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Cohn, J.S.; Johnson, E.J.; Millar, J.S.; Cohn, S.D.; Milne, R.W.; Marcel, Y.L.; Russell, R.M.; Schaefer, E.J. Contribution of apoB-48 and apoB-100 triglyceride-rich lipoproteins (TRL) to postprandial increases in the plasma concentration of TRL triglycerides and retinyl esters. J. Lipid Res. 1993, 34, 2033–2040. [Google Scholar]
- Havel, R.J. Remnant lipoproteins as therapeutic targets. Curr. Opin. Lipidol. 2000, 11, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, K.; Nakano, T.; Tokita, Y.; Nagamine, T.; Inazu, A.; Kobayashi, J.; Mabuchi, H.; Stanhope, K.L.; Havel, P.J.; Okazaki, M.; et al. Postprandial lipoprotein metabolism: VLDL vs chylomicrons. Clin. Chim. Acta 2011, 412, 1306–1318. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, K.; Nakano, T.; Tokita, Y.; Nagamine, T.; Yatsuzuka, S.; Shimomura, Y.; Tanaka, A.; Sumino, H.; Nara, M.; Machida, T.; et al. The characteristics of remnant lipoproteins in the fasting and postprandial plasma. Clin. Chim. Acta 2012, 413, 1077–1086. [Google Scholar] [CrossRef]
- Jackson, K.G.; Minihane, A.M. Postprandial lipemia and cardiovascular disease: Impact of age and gender on nonfasting triacylglycerol levels. Clin. Lipidol. 2010, 5, 1–4. [Google Scholar] [CrossRef]
- Nakajima, K.; Tanaka, A. Atherogenic postprandial RLP, VLDL remnants as a causal factor in atherosclerosis. Clin. Chim. Acta 2018, 478, 200–215. [Google Scholar] [CrossRef] [PubMed]
- Bjornstad, P.; Eckel, R.H. Pathogenesis of Lipid Disorders in Insulin Resistance: A Brief Review. Curr. Diab. Rep. 2018, 18, 127. [Google Scholar] [CrossRef] [PubMed]
- Krauss, R.M. Lipids and lipoproteins in patients with type 2 diabetes. Diabetes Care 2004, 27, 1496–1504. [Google Scholar] [CrossRef] [PubMed]
- Glass, C.K.; Olefsky, J.M. Inflammation and lipid signaling in the etiology of insulin resistance. Cell Metab. 2012, 15, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, K.; Tokita, Y.; Tanaka, A. Hypothesis: Postprandial remnant lipoproteins are the causal factors that induce resistance associated with obesity. Clin. Chim. Acta 2018, 485, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Salamanca, A.; Bárcena, B.; Arribas, C.; Fernández-Agulló, T.; Martínez, C.; Carrascosa, J.M.; Ros, M.; Andrés, A.; Gallardo, N. Aging impairs the hepatic subcellular distribution of ChREBP in response to fasting/feeding in rats: Implications on hepatic steatosis. Exp. Gerontol. 2015, 69, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escrivá, F.; Gavete, M.L.; Fermín, Y.; Pérez, C.; Gallardo, N.; Alvarez, C.; Andrés, A.; Ros, M.; Carrascosa, J.M. Effect of age and moderate food restriction on insulin sensitivity in Wistar rats: Role of adiposity. J. Endocrinol. 2007, 194, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Horrillo, D.; Gallardo, N.; Lauzurica, N.; Barrus, M.T.; San Frutos, M.G.; Andrés, A.; Ros, M.; Fernandez-Agullo, T. Development of liver fibrosis during aging: Effects of caloric restriction. J. Biol. Regul. Homeost. Agents 2013, 27, 377–388. [Google Scholar]
- Benhamed, F.; Denechaud, P.D.; Lemoine, M.; Robichon, C.; Moldes, M.; Bertrand-Michel, J.; Ratziu, V.; Serfaty, L.; Housset, C.; Capeau, J.; et al. The lipogenic transcription factor ChREBP dissociates hepatic steatosis from insulin resistance in mice and humans. J. Clin. Invest. 2012, 122, 2176–2194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Yeung, D.C.; Karpisek, M.; Stejskal, D.; Zhou, Z.G.; Liu, F.; Wong, R.L.; Chow, W.S.; Tso, A.W.; Lam, K.S.; et al. Serum FGF21 levels are increased in obesity and are independently associated with the metabolic syndrome in humans. Diabetes 2008, 57, 1246–1253. [Google Scholar] [CrossRef] [PubMed]
- Mraz, M.; Bartlova, M.; Lacinova, Z.; Michalsky, D.; Kasalicky, M.; Haluzikova, D.; Matoulek, M.; Dostalova, I.; Humenanska, V.; Haluzik, M. Serum concentrations and tissue expression of a novel endocrine regulator fibroblast growth factor-21 in patients with type 2 diabetes and obesity. Clin. Endocrinol. 2009, 71, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, D.J.; Steele, T.; Wilding, J.P.; Halford, J.C.; Harrold, J.A.; Hamer, M.; Karpe, F. What have human experimental overfeeding studies taught us about adipose tissue expansion and susceptibility to obesity and metabolic complications? Int. J. Obes. 2017, 41, 853–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porta, E.A.; Joun, N.S.; Nitta, R.T. Effects of the type of dietary fat at two levels of vitamin E in Wistar male rats during development and ageing. I. Life span, serum biochemical parameters and pathological changes. Mech. Ageing Dev. 1980, 13, 1–39. [Google Scholar] [CrossRef]
- Yorke, A.; Kane, A.E.; Hancock Friesen, C.L.; Howlett, S.E.; O’Blenes, S. Development of a Rat Clinical Frailty Index. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 897–903. [Google Scholar] [CrossRef] [PubMed]
- López, V.; Bonzón-Kulichenko, E.; Moltó, E.; Fernández-Agulló, T.; Arribas, C.; Andrés, A.; Gallardo, N. Food restriction is required to preserve the anti-steatotic effects of central leptin in the liver of middle-aged rats. Obesity 2018, 26, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Eggstein, M.; Kuhlmann, E. Triglycerides and glycerol determination after alkaline hydrolysis. In Methods of Enzymatic Analysis, 2nd ed.; Bergmeyer, H.U., Ed.; Academic Press: New York, USA, 1974; Volume 4, pp. 1825–1831. [Google Scholar]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Overturf, M.L.; Loose-Mitchell, D.S. In vivo model systems: The choice of the experimental animal model for analysis of lipoproteins and atherosclerosis. Curr. Opin. Lipidol. 1992, 3, 179–185. [Google Scholar] [CrossRef]
- Kim, M.S.; Krawczyk, S.A.; Doridot, L.; Fowler, A.J.; Wang, J.X.; Trauger, S.A.; Noh, H.L.; Kang, H.J.; Meissen, J.K.; Blatnik, M.; et al. ChREBP regulates fructose-induced glucose production independently of insulin signaling. J. Clin. Invest. 2016, 126, 4372–4386. [Google Scholar] [CrossRef] [Green Version]
- Bonzón-Kulichenko, E.; Moltó, E.; Pintado, C.; Fernández, A.; Arribas, C.; Schwudke, D.; Gallardo, N.; Shevchenko, A.; Andrés, A. Changes in visceral adipose tissue plasma membrane lipid composition in old rats are associated with adipocyte hypertrophy with aging. J. Gerontol. A Biol. Sci. Med. Sci. 2018, 73, 1139–1146. [Google Scholar] [CrossRef]
- De Fronzo, R.A. Pathogenesis of type 2 diabetes mellitus. Med. Clin. North Am. 2004, 88, 787–835. [Google Scholar] [CrossRef]
- Færch, K.; Vistisen, D.; Pacini, G.; Torekov, S.S.; Johansen, N.B.; Witte, D.R.; Jonsson, A.; Pedersen, O.; Hansen, T.; Lauritzen, T.; et al. Insulin resistance is accompanied by increased fasting glucagon and delayed glucagon suppression in individuals with normal and impaired glucose regulation. Diabetes 2016, 65, 3473–3481. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.D.; Finan, B.; Clemmensen, C.; DiMarchi, R.D.; Tschöp, M.H. The new biology and pharmacology of glucagon. Physiol. Rev. 2017, 97, 721–766. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Wahed, A.; Guilmeau, S.; Postic, C. Sweet sixteenth for ChREBP: Established roles and future goals. Cell Metab. 2017, 26, 324–341. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.Y.; Repa, J.J. The Liver X Receptor (LXR) and hepatic lipogenesis. The carbohydrate-response element-binding protein is a target gene of LXR. J. Biol. Chem. 2007, 282, 743–751. [Google Scholar] [CrossRef] [PubMed]
- Bindesboll, C.; Fan, Q.; Norgaard, N.C.; MacPherson, L.; Ruan, H.B.; Wu, J.; Pedersen, T.A.; Steffensen, K.R.; Yang, X.; Matthews, J.; et al. Liver X receptor regulates hepatic nuclear O-GlcNAc signaling and carbohydrate responsive element-binding protein activity. J. Lip. Res. 2015, 56, 771–785. [Google Scholar] [CrossRef] [Green Version]
- Ducheix, S.; Montagner, A.; Polizzi, A.; Lasserre, F.; Regnier, M.; Marmugi, A.; Benhamed, F.; Bertrand-Michel, J.; Mselli-Lakhal, L.; Loiseau, N.; et al. Dietary oleic acid regulates hepatic lipogenesis through a liver X receptor-dependent signaling. PLoS ONE 2017, 12, e0181393. [Google Scholar] [CrossRef] [PubMed]
- Seyer, P.; Vallois, D.; Poitry-Yamate, C.; Schütz, F.; Metref, S.; Tarussio, D.; Maechler, P.; Staels, B.; Lanz, B.; Grueter, R.; et al. Hepatic glucose sensing is required to preserve β cell glucose competence. J. Clin. Invest. 2013, 123, 1662–1676. [Google Scholar] [CrossRef]
- Tang, Y.; Wallace, M.; Sanchez-Gurmaches, J.; Hsiao, W.Y.; Li, H.; Lee, P.L.; Vernia, S.; Metallo, C.M.; Guertin, D.A. Adipose tissue mTORC2 regulates ChREBP-driven de novo lipogenesis and hepatic glucose metabolism. Nat. Commun. 2016, 7, 11365. [Google Scholar] [CrossRef] [Green Version]
- Herman, M.A.; Peroni, O.D.; Villoria, J.; Schön, R.; Abumrad, N.A.; Blüher, M.; Klein, S.; Kahn, B.B. A novel ChREBP isoform in adipose tissue regulates systemic glucose metabolism. Nature 2012, 484, 333–338. [Google Scholar] [CrossRef] [Green Version]
- Santos, A.L.; Lindner, A.B. Protein posttranslational modifications: Roles in aging and age-related disease. Oxid. Med. Cell Longv. 2017, 2017, 5716409. [Google Scholar] [CrossRef]
- Schlein, C.; Talukdar, S.; Heine, M.; Fischer, A.W.; Krott, L.M.; Nilsson, S.K.; Brenner, M.B.; Heeren, J.; Scheja, L. FGF21 Lowers Plasma Triglycerides by Accelerating Lipoprotein Catabolism in White and Brown Adipose Tissues. Cell Metab. 2016, 23, 441–453. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Xiaoli, A.M.; Yang, F. Regulation and Metabolic Significance of De Novo Lipogenesis in Adipose Tissues. Nutrients 2018, 10, 1383. [Google Scholar] [CrossRef]
- Kharitonenkov, A.; Shiyanova, T.L.; Koester, A.; Ford, A.M.; Micanovic, R.; Galbreath, E.J.; Sandusky, G.E.; Hammond, L.J.; Moyers, J.S.; Owens, R.A.; et al. FGF-21 as a novel metabolic regulator. J. Clin. Invest. 2005, 115, 1627–1635. [Google Scholar] [CrossRef] [Green Version]
- Inagaki, T.; Dutchak, P.; Zhao, G.; Ding, X.; Gautron, L.; Parameswara, V.; Li, Y.; Goetz, R.; Mohammadi, M.; Esser, V.; et al. Endocrine regulation of the fasting response by PPARalpha-mediated induction of fibroblast growth factor 21. Cell Metab. 2007, 5, 415–425. [Google Scholar] [CrossRef]
- Badman, M.K.; Pissios, P.; Kennedy, A.R.; Koukos, G.; Flier, J.S.; Maratos-Flier, E. Hepatic fibroblast growth factor 21 is regulated by PPARalpha and is a key mediator of hepatic lipid metabolism in ketotic states. Cell Metab. 2007, 5, 426–437. [Google Scholar] [CrossRef]
- Nygaard, E.B.; Møller, C.L.; Kievit, P.; Grove, K.L.; Andersen, B. Increased fibroblast growth factor 21 expression in high-fat diet-sensitive non-human primates (Macaca mulatta). Int. J. Obesity 2014, 38, 183–191. [Google Scholar] [CrossRef]
- Matikainen, N.; Taskinen, M.R.; Stennabb, S.; Lundbom, N.; Hakkarainen, A.; Vaaralahti, K.; Raivio, T. Decrease in circulating fibroblast growth factor 21 after an oral fat load is related to postprandial triglyceride-rich lipoproteins and liver fat. Eur. J. Endocrinol. 2012, 166, 487–492. [Google Scholar] [CrossRef]
- Hanks, L.J.; Gutiérrez, O.M.; Bamman, M.M.; Ashraf, A.; McCormick, K.L.; Casazza, K. Circulating levels of fibroblast growth factor-21 increase with age independently of body composition indices among healthy individuals. J. Clin. Transl. Endocrinol. 2015, 2, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Villarroya, J.; Gallego-Escuredo, J.M.; Delgado-Anglés, A.; Cairó, M.; Moure, R.; Mateo, M.G.; Domingo, J.C.; Domingo, P.; Giralt, M.; Villarroya, F. Aging is associated with increased FGF21 levels but unaltered FGF21 responsiveness in adipose tissue. Aging Cell 2018, 17, e12822. [Google Scholar] [CrossRef]
- Lewis, J.E.; Samms, R.J.; Cooper, S.; Luckett, J.C.; Perkins, A.C.; Adams, A.C.; Tsintzas, K.; Ebling, F.J.P. Reduced adiposity attenuates FGF21 mediated metabolic improvements in the Siberian hamster. Sci. Rep. 2017, 7, 4238. [Google Scholar] [CrossRef] [Green Version]
- Keipert, S.; Kutschke, M.; Ost, M.; Schwarzmayr, T.; van Schothorst, E.M.; Lamp, D.; Brachthäuser, L.; Hamp, I.; Mazibuko, S.E.; Hartwig, S.; et al. Long-Term Cold Adaptation Does Not Require FGF21 or UCP1. Cell Metab. 2017, 26, 437–446. [Google Scholar] [CrossRef]
- Rogers, N.H.; Landa, A.; Park, S.; Smith, R.G. Aging leads to a programmed loss of brown adipocytes in murine subcutaneous white adipose tissue. Aging Cell 2012, 11, 1074–1083. [Google Scholar] [CrossRef] [Green Version]
- Sepa-Kishi, D.M.; Ceddia, R.B. Circulating fibroblast growth factor 21 is reduced, whereas its production is increased in a fat depot-specific manner in cold-acclimated rats. Adipocyte 2018, 7, 238–247. [Google Scholar] [CrossRef] [Green Version]
- Cousin, B.; Cinti, S.; Morroni, M.; Raimbault, S.; Ricquier, D.; Pénicaud, L.; Casteilla, L. Occurrence of brown adipocytes in rat white adipose tissue: Molecular and morphological characterization. J. Cell Sci. 1992, 103, 931–942. [Google Scholar]
- Shin, S.; Ajuwon, K.M. Effects of diets differing in composition of 18-C fatty acids on adipose tissue thermogenic gene expression in mice fed high-fat diets. Nutrients 2018, 10, 256. [Google Scholar] [CrossRef]
- Li, M.V.; Chang, B.; Imamura, M.; Poungvarin, N.; Chan, L. Glucose-dependent transcriptional regulation by an evolutionarily conserved glucose-sensing module. Diabetes 2006, 55, 1179–1189. [Google Scholar] [CrossRef]
- Iizuka, K.; Takao, K.; Kato, T.; Horikawa, Y.; Takeda, J. ChREBP Reciprocally Regulates Liver and Plasma Triacylglycerol Levels in Different Manners. Nutrients 2018, 10, 1699. [Google Scholar] [CrossRef]
- Hojna, S.; Jordan, M.D.; Kollias, H.; Pausova, Z. High-fat diet induces emergence of brown-like adipocytes in white adipose tissue of spontaneously hypertensive rats. Hypertens. Res. 2012, 35, 279–286. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wistar Rats | 3mAL | 7mAL | 7mFR | 24mAL | 24mFR |
|---|---|---|---|---|---|
| Body weight (g) | 325 ± 17 | 425 ± 7 & | 382 ± 4 + | 646 ±19 &,* | 490 ±12 &,+ |
| Mean daily food intake (g) | 22 ± 1.2 | 22 ± 0.4 | 17 ± 0.5 + | 23 ± 0.7 | 18 ± 0.3 + |
| Visceral adiposity index (%) | 2.1 ± 0.2 | 5.7 ± 0.3 & | 1.2 ± 0.04 + | 7.0 ± 0.5 &,* | 1.9 ± 0.1 + |
| Fasting serum leptin (ng/mL) | 4.7 ± 0.6 | 7.5 ± 0.9 & | 2.1 ± 0.1 + | 16.3 ± 2 &,* | 7.1 ± 1.2 + |
| HOMA-IR | 7.5 ± 1.5 | 7.8 ± 1.9 | 2.4 ± 0.5 + | 10.1 ± 1 &,* | 7.3 ± 1.2 |
| Wistar Rats | 7mAL | 7mFR | 24mAL | 24mFR |
|---|---|---|---|---|
| Fasting serum glucose (mg/dL) | 119 ± 5 | 128 ± 7 | 131 ± 5 | 112 ± 22 |
| Fasting serum insulin (ng/mL) | 2.0 ± 0.1 | 1.4 ± 0.9 | 2.8 ± 0.1 | 2.1 ± 1 |
| Fasting serum glucagon (pg/mL) | 671 ± 29 | 574 ± 5 | 1328 ± 62 * | 571 ± 8 + |
| Fasting serum TAG (mg/dl) | 156 ± 9 | 98 ± 14 + | 171 ± 8 * | 103 ± 4 + |
| Fasting serum NEFA (mmol/L) | 1.27 ± 0.1 | 0.84 ± 0.002 + | 1.08 ± 0.03 * | 0.87 ± 0.06 + |
| Fasting serum glycerol (mg/dl) | 47 ± 4 | 36 ± 0.9 + | 39 ± 3 * | 24 ± 7 + |
| Fasting serum KB (mM) | 0.18 ± 0.01 | 0.06 ± 0.01 + | 0.081 ± 0.008 * | 0.06 ± 0.002 + |
| Fasting serum Cho (mg/dL) | 101 ± 2 | 91 ± 19 | 120 ± 13 * | 107 ± 16 |
| Fasting serum cHDL(mg/dL) | 32.7 ± 1 | 53.0 ± 14 + | 15.8 ± 1 * | 16.2 ± 1 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández, A.; Mazuecos, L.; Pintado, C.; Rubio, B.; López, V.; de Solís, A.J.; Rodríguez, M.; Andrés, A.; Gallardo, N. Effects of Moderate Chronic Food Restriction on the Development of Postprandial Dyslipidemia with Ageing. Nutrients 2019, 11, 1865. https://doi.org/10.3390/nu11081865
Fernández A, Mazuecos L, Pintado C, Rubio B, López V, de Solís AJ, Rodríguez M, Andrés A, Gallardo N. Effects of Moderate Chronic Food Restriction on the Development of Postprandial Dyslipidemia with Ageing. Nutrients. 2019; 11(8):1865. https://doi.org/10.3390/nu11081865
Chicago/Turabian StyleFernández, Alejandro, Lorena Mazuecos, Cristina Pintado, Blanca Rubio, Virginia López, Alain J. de Solís, María Rodríguez, Antonio Andrés, and Nilda Gallardo. 2019. "Effects of Moderate Chronic Food Restriction on the Development of Postprandial Dyslipidemia with Ageing" Nutrients 11, no. 8: 1865. https://doi.org/10.3390/nu11081865
APA StyleFernández, A., Mazuecos, L., Pintado, C., Rubio, B., López, V., de Solís, A. J., Rodríguez, M., Andrés, A., & Gallardo, N. (2019). Effects of Moderate Chronic Food Restriction on the Development of Postprandial Dyslipidemia with Ageing. Nutrients, 11(8), 1865. https://doi.org/10.3390/nu11081865