Polyphenols and Metabolites Enhance Survival in Rodents and Nematodes—Impact of Mitochondria

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and RR
2.2. Characterization of Phenolic Compounds in RR
2.2.1. Sample Preparation
2.2.2. UHPLC and MS Settings
2.3. Nematode (Caenorhabditis elegans)
2.3.1. C. elegans and Bacterial Strains
2.3.2. Cultivation and Treatment
2.3.3. Lifespan
2.3.4. Heat-stress Resistance
2.3.5. Quantitative Real-time PCR
2.3.6. Isolation of Mitochondria
2.3.7. High-resolution Respirometry
2.3.8. Mitochondrial Membrane Potential (ΔΨm)
2.3.9. Citrate-Synthase Activity
2.3.10. Nematode Homogenization
2.3.11. Determination of ATP
2.3.12. Determination of Lactate and Pyruvate Content
2.3.13. Protein Quantification
2.4. Rodents
2.4.1. Animals and Treatment
2.4.2. Survival
2.5. Statistics
3. Results
3.1. Characterization of Phenolic Compounds in RR
3.1.1. Hydroxybenzoic and Hydroxycinnamic Acids
3.1.2. Flavanones
3.1.3. Flavones
3.1.4. Flavonols
3.1.5. Further Phenolic Compounds
3.2. Effect on Lifespan in Mice and Nematodes
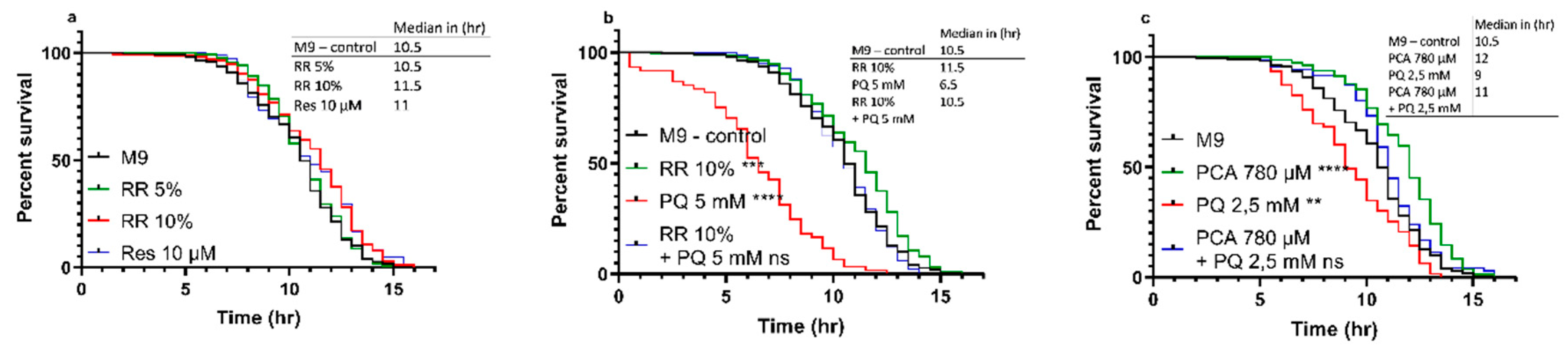
3.2.1. Physiological Lifespan
3.2.2. Heat-stress Resistance
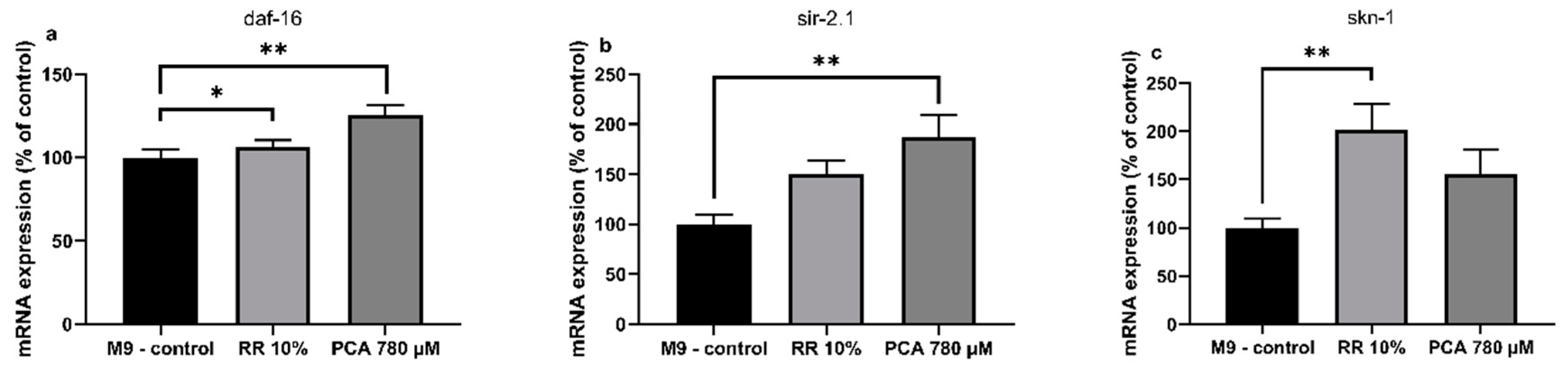
3.3. Quantitatice real-time PCR
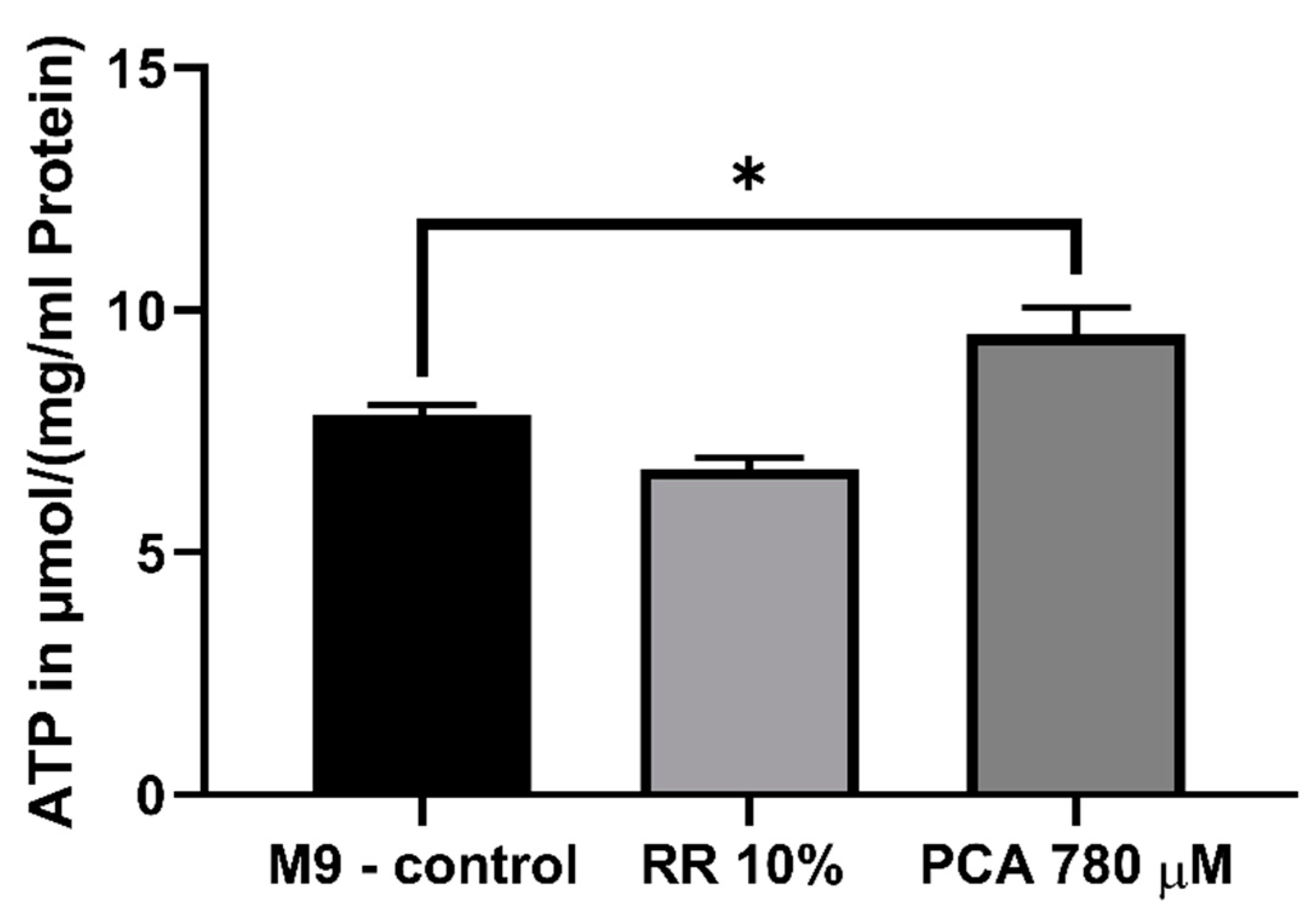
3.4. Mitochondrial Function
3.5. Glycolysis
4. Discussion
4.1. RR and PCA Improve Life and Health-span
4.2. Ameliorated Mitochondrial Function Through RR and PCA
4.3. Phenolic Compounds in RR
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pray, L. Nutrition across the Lifespan for Healthy Aging. In Proceedings of the National Academies of Sciences, Engineering, and Medicine’s Food Forum, Washington DC, USA, 12 May 2017. [Google Scholar]
- Tzioras, M.; Davies, C.; Newman, A.; Jackson, R.; Spires-Jones, T. APOE at the interface of inflammation, neurodegeneration and pathological protein spread in Alzheimer’s disease. Neuropathol. Appl. Neurobiol. 2018, 45, 327–346. [Google Scholar] [CrossRef]
- Mico, V.; Berninches, L.; Tapia, J.; Daimiel, L. NutrimiRAging: Micromanaging Nutrient Sensing Pathways through Nutrition to Promote Healthy Aging. Int. J. Mol. Sci. 2017, 18, E915. [Google Scholar] [CrossRef]
- Gonzalez-Freire, M.; de Cabo, R.; Bernier, M.; Sollott, S.J.; Fabbri, E.; Navas, P.; Ferrucci, L. Reconsidering the Role of Mitochondria in Aging. J. Gerontol. A Biol. Sci. Med. Sci. 2015, 70, 1334–1342. [Google Scholar] [CrossRef] [Green Version]
- Sebastian, D.; Palacin, M.; Zorzano, A. Mitochondrial Dynamics: Coupling Mitochondrial Fitness with Healthy Aging. Trends Mol. Med. 2017, 23, 201–215. [Google Scholar] [CrossRef]
- Van Giau, V.; An, S.S.A.; Hulme, J.P. Mitochondrial therapeutic interventions in Alzheimer’s disease. J. Neurol. Sci. 2018, 395, 62–70. [Google Scholar] [CrossRef]
- Macdonald, R.; Barnes, K.; Hastings, C.; Mortiboys, H. Mitochondrial abnormalities in Parkinson’s disease and Alzheimer’s disease: Can mitochondria be targeted therapeutically? Biochem. Soc. Trans. 2018, 46, 891–909. [Google Scholar] [CrossRef]
- Stockburger, C.; Eckert, S.; Eckert, G.P.; Friedland, K.; Muller, W.E. Mitochondrial Function, Dynamics, and Permeability Transition: A Complex Love Triangle as A Possible Target for the Treatment of Brain Aging and Alzheimer’s Disease. J. Alzheimers Dis. 2018, 64, S455–S467. [Google Scholar] [CrossRef]
- Kauppila, T.E.S.; Kauppila, J.H.K.; Larsson, N.-G. Mammalian Mitochondria and Aging: An Update. Cell Metab. 2017, 25, 57–71. [Google Scholar] [CrossRef]
- Yusufov, M.; Weyandt, L.L.; Piryatinsky, I. Alzheimer’s disease and diet: A systematic review. Int. J. Neurosci. 2017, 127, 161–175. [Google Scholar] [CrossRef]
- Petersson, S.D.; Philippou, E. Mediterranean Diet, Cognitive Function, and Dementia: A Systematic Review of the Evidence. Adv. Nutr. 2016, 7, 889–904. [Google Scholar] [CrossRef] [Green Version]
- Feart, C.; Samieri, C.; Barberger-Gateau, P. Mediterranean diet and cognitive health: An update of available knowledge. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 51–62. [Google Scholar] [CrossRef]
- Kim, H.G.; Oh, M.S. Herbal medicines for the prevention and treatment of Alzheimer’s disease. Curr. Pharm. Des. 2012, 18, 57–75. [Google Scholar]
- Bohn, T. Dietary factors affecting polyphenol bioavailability. Nutr. Rev. 2014, 72, 429–452. [Google Scholar] [CrossRef]
- Marin, L.; Miguelez, E.M.; Villar, C.J.; Lombo, F. Bioavailability of dietary polyphenols and gut microbiota metabolism: antimicrobial properties. Biomed. Res. Int. 2015, 2015, 905215. [Google Scholar] [CrossRef]
- Bors, W.; Michel, C.; Stettmaier, K. Structure-activity relationships governing antioxidant capacities of plant polyphenols. Methods Enzymol. 2001, 335, 166–180. [Google Scholar]
- Watzl, B.; Leitzmann, C. Bioaktive Substanzen in Lebensmitteln: 61 Tabellen, 3rd ed.; Hippokrates-Verl: Stuttgart, Germany, 2005; p. S1528. [Google Scholar]
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligne, B.; Ganzle, M.; Kort, R.; Pasin, G.; Pihlanto, A.; et al. Health benefits of fermented foods: Microbiota and beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar] [CrossRef]
- Septembre-Malaterre, A.; Remize, F.; Poucheret, P. Fruits and vegetables, as a source of nutritional compounds and phytochemicals: Changes in bioactive compounds during lactic fermentation. Food Res. Int. 2018, 104, 86–99. [Google Scholar] [CrossRef]
- Hippeli, S.; Janisch, K.; Kern, S.; Ölschläger, C.; Treutter, D.; May, C.; Elstner, E.F. Antioxidant and immune modulatory activities of fruit and vegetable extracts after “Cascade Fermentation”. Curr. Top. Biochem. 2007, 9, 83–97. [Google Scholar]
- Krzysztoforska, K.; Mirowska-Guzel, D.; Widy-Tyszkiewicz, E. Pharmacological effects of protocatechuic acid and its therapeutic potential in neurodegenerative diseases: Review on the basis of in vitro and in vivo studies in rodents and humans. Nutr. Neurosci. 2019, 22, 1–11. [Google Scholar] [CrossRef]
- Maulik, N.; McFadden, D.; Otani, H.; Thirunavukkarasu, M.; Parinandi, N.L. Antioxidants in Longevity and Medicine. Oxid. Med. Cell Longev. 2013, 2013, 820679. [Google Scholar] [CrossRef]
- Semaming, Y.; Sripetchwandee, J.; Sa-Nguanmoo, P.; Pintana, H.; Pannangpetch, P.; Chattipakorn, N.; Chattipakorn, S.C. Protocatechuic acid protects brain mitochondrial function in streptozotocin-induced diabetic rats. Appl. Physiol. Nutr. Metab. 2015, 40, 1078–1081. [Google Scholar] [CrossRef]
- Kakkar, S.; Bais, S. A review on protocatechuic acid and its pharmacological potential. ISRN Pharmacol. 2014, 2014, 952943. [Google Scholar] [CrossRef]
- Procédé de PréParation D’un Produit Naturel Par Fermentation. Available online: https://patents.google.com/patent/EP1153549A3/en (accessed on 8 August 2019).
- Katiki, L.M.; Ferreira, J.F.S.; Zajac, A.M.; Masler, C.; Lindsay, D.S.; Chagas, A.C.S.; Amarante, A.F.T. Caenorhabditis elegans as a model to screen plant extracts and compounds as natural anthelmintics for veterinary use. Vet. Parasitol. 2011, 182, 264–268. [Google Scholar] [CrossRef]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar]
- Stiernagle, T. Maintenance of C. elegans. WormBook 2006, 11, 1–11. [Google Scholar] [CrossRef]
- Amrit, F.R.G.; Ratnappan, R.; Keith, S.A.; Ghazi, A. The C. elegans lifespan assay toolkit. Methods 2014, 68, 465–475. [Google Scholar] [CrossRef]
- Fitzenberger, E.; Deusing, D.J.; Marx, C.; Boll, M.; Lüersen, K.; Wenzel, U. The polyphenol quercetin protects the mev-1 mutant of Caenorhabditis elegans from glucose-induced reduction of survival under heat-stress depending on SIR-2.1, DAF-12, and proteasomal activity. Mol. Nutr. Food Res. 2014, 58, 984–994. [Google Scholar] [CrossRef]
- Schmitt, S.; Saathoff, F.; Meissner, L.; Schropp, E.-M.; Lichtmannegger, J.; Schulz, S.; Eberhagen, C.; Borchard, S.; Aichler, M.; Adamski, J.; et al. A semi-automated method for isolating functionally intact mitochondria from cultured cells and tissue biopsies. Anal. Biochem. 2013, 443, 66–74. [Google Scholar] [CrossRef]
- Balch, W.E.; Rothman, J.E. Characterization of protein transport between successive compartments of the Golgi apparatus: Asymmetric properties of donor and acceptor activities in a cell-free system. Arch. Biochem. Biophys. 1985, 240, 413–425. [Google Scholar] [CrossRef]
- Stadlmann, S.; Renner, K.; Pollheimer, J.; Moser, P.L.; Zeimet, A.G.; Offner, F.A.; Gnaiger, E. Preserved coupling of oxidative phosphorylation but decreased mitochondrial respiratory capacity in IL-1beta-treated human peritoneal mesothelial cells. Cell Biochem. Biophys. 2006, 44, 179–186. [Google Scholar] [CrossRef]
- Hagl, S.; Kocher, A.; Schiborr, C.; Eckert, S.H.; Ciobanu, I.; Birringer, M.; El-Askary, H.; Helal, A.; Khayyal, M.T.; Frank, J.; et al. Rice bran extract protects from mitochondrial dysfunction in guinea pig brains. Pharmacol. Res. 2013, 76, 17–27. [Google Scholar] [CrossRef]
- Kuznetsov, A.V.; Strobl, D.; Ruttmann, E.; Königsrainer, A.; Margreiter, R.; Gnaiger, E. Evaluation of mitochondrial respiratory function in small biopsies of liver. Anal. Biochem. 2002, 305, 186–194. [Google Scholar] [CrossRef]
- Hutter, E.; Renner, K.; Pfister, G.; Stöckl, P.; Jansen-Dürr, P.; Gnaiger, E. Senescence-associated changes in respiration and oxidative phosphorylation in primary human fibroblasts. Biochem. J. 2004, 380, 919–928. [Google Scholar] [CrossRef] [Green Version]
- Larsen, S.; Nielsen, J.; Hansen, C.N.; Nielsen, L.B.; Wibrand, F.; Stride, N.; Schroder, H.D.; Boushel, R.; Helge, J.W.; Dela, F.; et al. Biomarkers of mitochondrial content in skeletal muscle of healthy young human subjects. J. Physiol. 2012, 590, 3349–3360. [Google Scholar] [CrossRef]
- Rabol, R.; Larsen, S.; Hojberg, P.M.V.; Almdal, T.; Boushel, R.; Haugaard, S.B.; Andersen, J.L.; Madsbad, S.; Dela, F. Regional anatomic differences in skeletal muscle mitochondrial respiration in type 2 diabetes and obesity. J. Clin. Endocrinol. Metab. 2010, 95, 857–863. [Google Scholar] [CrossRef]
- Asseburg, H.; Schafer, C.; Muller, M.; Hagl, S.; Pohland, M.; Berressem, D.; Borchiellini, M.; Plank, C.; Eckert, G.P. Effects of Grape Skin Extract on Age-Related Mitochondrial Dysfunction, Memory and Life Span in C57BL/6J Mice. Neuromolecular. Med. 2016, 18, 378–395. [Google Scholar] [CrossRef]
- Gower, A.J.; Lamberty, Y. The aged mouse as a model of cognitive decline with special emphasis on studies in NMRI mice. Behav. Brain. Res. 1993, 57, 163–173. [Google Scholar] [CrossRef]
- Fitzenberger, E.; Deusing, D.J.; Wittkop, A.; Kler, A.; Kriesl, E.; Bonnlander, B.; Wenzel, U. Effects of plant extracts on the reversal of glucose-induced impairment of stress-resistance in Caenorhabditis elegans. Plant Foods Hum. Nutr. 2014, 69, 78–84. [Google Scholar] [CrossRef]
- Castelein, N.; Muschol, M.; Dhondt, I.; Cai, H.; de Vos, W.H.; Dencher, N.A.; Braeckman, B.P. Mitochondrial efficiency is increased in axenically cultured Caenorhabditis elegans. Exp. Gerontol. 2014, 56, 26–36. [Google Scholar] [CrossRef]
- Zheng, J.; Meenu, M.; Xu, B. A systematic investigation on free phenolic acids and flavonoids profiles of commonly consumed edible flowers in China. J. Pharmaceut. Biomed. 2019, 172, 268–277. [Google Scholar] [CrossRef]
- Chen, W.; Wang, D.; Wang, L.-S.; Di, B.; Wang, J.; See, W.A.; Mallery, S.R.; Stoner, G.D.; Liu, Z. Pharmacokinetics of protocatechuic acid in mouse and its quantification in human plasma using LC-tandem mass spectrometry. J. Chromatogr. B. Analyt. Technol. Biomed. Life Sci. 2012, 908, 39–44. [Google Scholar] [CrossRef]
- Jardim, F.R.; de Rossi, F.T.; Nascimento, M.X.; da Silva, B.; Borges, P.A.; Prescilio, I.C.; de Oliveira, M.R. Resveratrol and Brain Mitochondria: A Review. Mol. Neurobiol. 2018, 55, 2085–2101. [Google Scholar] [CrossRef]
- Sarubbo, F.; Moranta, D.; Asensio, V.J.; Miralles, A.; Esteban, S. Effects of Resveratrol and Other Polyphenols on the Most Common Brain Age-Related Diseases. Curr. Med. Chem. 2017, 24, 4245–4266. [Google Scholar] [CrossRef]
- Sawikr, Y.; Yarla, N.S.; Peluso, I.; Kamal, M.A.; Aliev, G.; Bishayee, A. Neuroinflammation in Alzheimer’s Disease: The Preventive and Therapeutic Potential of Polyphenolic Nutraceuticals. Adv. Protein Chem. Struct. Biol. 2017, 108, 33–57. [Google Scholar]
- Casamenti, F.; Stefani, M. Olive polyphenols: New promising agents to combat aging-associated neurodegeneration. Expert. Rev. Neurother. 2017, 17, 345–358. [Google Scholar] [CrossRef]
- Hagl, S.; Asseburg, H.; Heinrich, M.; Sus, N.; Blumrich, E.-M.; Dringen, R.; Frank, J.; Eckert, G.P. Effects of Long-Term Rice Bran Extract Supplementation on Survival, Cognition and Brain Mitochondrial Function in Aged NMRI Mice. Neuromolecular. Med. 2016, 18, 347–363. [Google Scholar] [CrossRef]
- Cocheme, H.M.; Murphy, M.P. Complex I is the major site of mitochondrial superoxide production by paraquat. J. Biol. Chem. 2008, 283, 1786–1798. [Google Scholar] [CrossRef]
- Possik, E.; Pause, A. Measuring oxidative stress resistance of Caenorhabditis elegans in 96-well microtiter plates. J. Vis. Exp. 2015, 9, e52746. [Google Scholar] [CrossRef]
- Navarro, A.; Gomez, C.; Sanchez-Pino, M.-J.; Gonzalez, H.; Bandez, M.J.; Boveris, A.D.; Boveris, A. Vitamin E at high doses improves survival, neurological performance, and brain mitochondrial function in aging male mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R1392–R1399. [Google Scholar] [CrossRef] [Green Version]
- Kitani, K.; Osawa, T.; Yokozawa, T. The effects of tetrahydrocurcumin and green tea polyphenol on the survival of male C57BL/6 mice. Biogerontology 2007, 8, 567–573. [Google Scholar] [CrossRef]
- Kim, Y.S.; Seo, H.W.; Lee, M.-H.; Kim, D.K.; Jeon, H.; Cha, D.S. Protocatechuic acid extends lifespan and increases stress resistance in Caenorhabditis elegans. Arch. Pharm. Res. 2014, 37, 245–252. [Google Scholar] [CrossRef]
- Sunagawa, T.; Shimizu, T.; Kanda, T.; Tagashira, M.; Sami, M.; Shirasawa, T. Procyanidins from apples (Malus pumila Mill.) extend the lifespan of Caenorhabditis elegans. Planta. Med. 2011, 77, 122–127. [Google Scholar] [CrossRef]
- Abbas, S.; Wink, M. Epigallocatechin gallate from green tea (Camellia sinensis) increases lifespan and stress resistance in Caenorhabditis elegans. Planta. Med. 2009, 75, 216–221. [Google Scholar] [CrossRef]
- Yoon, D.S.; Cha, D.S.; Choi, Y.; Lee, J.W.; Lee, M.-H. MPK-1/ERK is required for the full activity of resveratrol in extended lifespan and reproduction. Aging Cell. 2018, 18, e12867. [Google Scholar] [CrossRef] [Green Version]
- Park, H.-E.H.; Jung, Y.; Lee, S.-J.V. Survival assays using Caenorhabditis elegans. Mol. Cells. 2017, 40, 90–99. [Google Scholar] [CrossRef]
- Labbadia, J.; Morimoto, R.I. The biology of proteostasis in aging and disease. Annu. Rev. Biochem. 2015, 84, 435–464. [Google Scholar] [CrossRef]
- Kourtis, N.; Nikoletopoulou, V.; Tavernarakis, N. Small heat-shock proteins protect from heat-stroke-associated neurodegeneration. Nature 2012, 490, 213–218. [Google Scholar] [CrossRef]
- Pietsch, K.; Saul, N.; Chakrabarti, S.; Sturzenbaum, S.R.; Menzel, R.; Steinberg, C.E.W. Hormetins, antioxidants and prooxidants: defining quercetin-, caffeic acid- and rosmarinic acid-mediated life extension in C. elegans. Biogerontology 2011, 12, 329–347. [Google Scholar] [CrossRef]
- Wilson, M.A.; Shukitt-Hale, B.; Kalt, W.; Ingram, D.K.; Joseph, J.A.; Wolkow, C.A. Blueberry polyphenols increase lifespan and thermotolerance in Caenorhabditis elegans. Aging Cell 2006, 5, 59–68. [Google Scholar] [CrossRef]
- Guha, S.; Natarajan, O.; Murbach, C.G.; Dinh, J.; Wilson, E.C.; Cao, M.; Zou, S.; Dong, Y. Supplement timing of cranberry extract plays a key role in promoting Caenorhabditis elegans healthspan. Nutrients 2014, 6, 911–921. [Google Scholar] [CrossRef]
- Saier, C.; Gommlich, I.; Hiemann, V.; Baier, S.; Koch, K.; Horn, G.; Kowalewsky, T.; Bartelt, J.; Seemann, M.; Watjen, W. Agrimonia. Procera. Wallr. Extract Increases Stress Resistance and Prolongs Life Span in Caenorhabditis elegans via Transcription Factor DAF-16 (FoxO Orthologue). Antioxidants 2018, 7, 12. [Google Scholar] [CrossRef]
- Viswanathan, M.; Kim, S.K.; Berdichevsky, A.; Guarente, L. A role for SIR-2.1 regulation of ER stress response genes in determining C. elegans life span. Dev. Cell 2005, 9, 605–615. [Google Scholar] [CrossRef]
- Xiong, L.-G.; Chen, Y.-J.; Tong, J.-W.; Gong, Y.-S.; Huang, J.-A.; Liu, Z.-H. Epigallocatechin-3-gallate promotes healthy lifespan through mitohormesis during early-to-mid adulthood in Caenorhabditis elegans. Redox. Biol. 2018, 14, 305–315. [Google Scholar] [CrossRef]
- Lee, J.; Kwon, G.; Park, J.; Kim, J.-K.; Lim, Y.-H. Brief Communication: SIR-2.1-dependent lifespan extension of Caenorhabditis elegans by oxyresveratrol and resveratrol. Exp. Biol. Med. 2016, 241, 1757–1763. [Google Scholar] [CrossRef]
- Zheng, S.-Q.; Huang, X.-B.; Xing, T.-K.; Ding, A.-J.; Wu, G.-S.; Luo, H.-R. Chlorogenic Acid Extends the Lifespan of Caenorhabditis elegans via Insulin/IGF-1 Signaling Pathway. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 464–472. [Google Scholar]
- Li, H.; Su, L.; Su, X.; Liu, X.; Wang, D.; Li, H.; Ba, X.; Zhang, Y.; Lu, J.; Huang, B.; et al. Arginine methylation of SKN-1 promotes oxidative stress resistance in Caenorhabditis elegans. Redox. Biol. 2019, 21, 101111. [Google Scholar] [CrossRef]
- Link, P.; Wink, M. Isoliquiritigenin exerts antioxidant activity in Caenorhabditis elegans via insulin-like signaling pathway and SKN-1. Phytomedicine 2018, 55, 119–124. [Google Scholar] [CrossRef]
- Lee, E.H.-C.; Lim, S.S.-C.; Yuen, K.-H.; Lee, C.-Y. Curcumin and a hemi-analogue with improved blood-brain barrier permeability protect against amyloid-beta toxicity in Caenorhabditis elegans via SKN-1/Nrf activation. J Pharm. Pharmacol. 2019, 71, 860–868. [Google Scholar] [CrossRef]
- Kong, Y.; Trabucco, S.E.; Zhang, H. Oxidative stress, mitochondrial dysfunction and the mitochondria theory of aging. J. Signal Transduct. 2014, 39, 86–107. [Google Scholar]
- Grimm, A.; Friedland, K.; Eckert, A. Mitochondrial dysfunction: The missing link between aging and sporadic Alzheimer’s disease. Biogerontology 2016, 17, 281–296. [Google Scholar] [CrossRef]
- Wang, X.; Wang, W.; Li, L.; Perry, G.; Lee, H.-g.; Zhu, X. Oxidative stress and mitochondrial dysfunction in Alzheimer’s disease. Biochim. Biophys. Acta 2014, 1842, 1240–1247. [Google Scholar] [CrossRef]
- Hagl, S.; Berressem, D.; Grewal, R.; Sus, N.; Frank, J.; Eckert, G.P. Rice bran extract improves mitochondrial dysfunction in brains of aged NMRI mice. Nutr. Neurosci. 2016, 19, 1–10. [Google Scholar] [CrossRef]
- Teixeira, J.; Chavarria, D.; Borges, F.; Wojtczak, L.; Wieckowski, M.R.; Karkucinska-Wieckowska, A.; Oliveira, P.J. Dietary Polyphenols and Mitochondrial Function: Role in Health and Disease. Curr Med Chem. 2017. [Google Scholar] [CrossRef]
- Friedman, J.R.; Nunnari, J. Mitochondrial form and function. Nature 2014, 505, 335–343. [Google Scholar] [CrossRef] [Green Version]
- Palikaras, K.; Lionaki, E.; Tavernarakis, N. Coordination of mitophagy and mitochondrial biogenesis during ageing in C. elegans. Nature 2015, 521, 525–528. [Google Scholar] [CrossRef]
- Berdichevsky, A.; Viswanathan, M.; Horvitz, H.R.; Guarente, L. C. elegans SIR-2.1 interacts with 14-3-3 proteins to activate DAF-16 and extend life span. Cell 2006, 125, 1165–1177. [Google Scholar] [CrossRef]
- Van der Bliek, A.M.; Sedensky, M.M.; Morgan, P.G. Cell Biology of the Mitochondrion. Genetics 2017, 207, 843–871. [Google Scholar] [CrossRef] [Green Version]
- Kanno, S.; Hirano, S.; Mukai, T.; Ro, A.; Kato, H.; Fukuta, M.; Aoki, Y. Cellular uptake of paraquat determines subsequent toxicity including mitochondrial damage in lung epithelial cells. Leg. Med. 2018, 37, 7–14. [Google Scholar] [CrossRef]
- Nicholls, D.G. Mitochondrial membrane potential and aging. Aging Cell 2004, 3, 35–40. [Google Scholar] [CrossRef]
- Moreno, A.J.; Santos, D.L.; Magalhaes-Novais, S.; Oliveira, P.J. Measuring Mitochondrial Membrane Potential with a Tetraphenylphosphonium-Selective Electrode. Curr. Protoc. Toxicol. 2015, 65, 1–16. [Google Scholar]
- Perry, S.W.; Norman, J.P.; Barbieri, J.; Brown, E.B.; Gelbard, H.A. Mitochondrial membrane potential probes and the proton gradient: A practical usage guide. BioTechniques 2011, 50, 98–115. [Google Scholar] [CrossRef]
- Qi, G.; Mi, Y.; Fan, R.; Zhao, B.; Ren, B.; Liu, X. Tea polyphenols ameliorates neural redox imbalance and mitochondrial dysfunction via mechanisms linking the key circadian regular Bmal1. Food Chem. Toxicol. 2017, 110, 189–199. [Google Scholar] [CrossRef]
- Fang, W.-J.; Wang, C.-J.; He, Y.; Zhou, Y.-L.; Peng, X.-D.; Liu, S.-K. Resveratrol alleviates diabetic cardiomyopathy in rats by improving mitochondrial function through PGC-1alpha deacetylation. Acta Pharmacol. Sin. 2018, 39, 59–73. [Google Scholar] [CrossRef]
- Kumar, S.; Stokes, J.; Singh, U.P.; Scissum-Gunn, K.; Singh, R.; Manne, U.; Mishra, M.K. Prolonged exposure of resveratrol induces reactive superoxide species-independent apoptosis in murine prostate cells. Tumour. Biol. 2017, 39, 1010428317715039. [Google Scholar] [CrossRef]
- Liu, Y.-M.; Jiang, B.; Bao, Y.-M.; An, L.-J. Protocatechuic acid inhibits apoptosis by mitochondrial dysfunction in rotenone-induced PC12 cells. Toxicol. In Vitro 2008, 22, 430–437. [Google Scholar] [CrossRef]
- Guan, S.; Jiang, B.; Bao, Y.M.; An, L.J. Protocatechuic acid suppresses MPP+ -induced mitochondrial dysfunction and apoptotic cell death in PC12 cells. Food Chem. Toxicol. 2006, 44, 1659–1666. [Google Scholar] [CrossRef]
- Rafiei, H.; Omidian, K.; Bandy, B. Dietary Polyphenols Protect Against Oleic Acid-Induced Steatosis in an in Vitro Model of NAFLD by Modulating Lipid Metabolism and Improving Mitochondrial Function. Nutrients 2019, 11, 541. [Google Scholar] [CrossRef]
- Reutzel, M.; Grewal, R.; Silaidos, C.; Zotzel, J.; Marx, S.; Tretzel, J.; Eckert, G.P. Effects of Long-Term Treatment with a Blend of Highly Purified Olive Secoiridoids on Cognition and Brain ATP Levels in Aged NMRI Mice. Oxid. Med. Cell Longev. 2018, 2018, 4070935. [Google Scholar] [CrossRef]
- Ulicna, O.; Vancova, O.; Kucharska, J.; Janega, P.; Waczulikova, I. Rooibos tea (Aspalathus linearis) ameliorates the CCl4-induced injury to mitochondrial respiratory function and energy production in rat liver. Gen. Physiol. Biophys. 2019, 38, 15–25. [Google Scholar] [CrossRef]
- Yanase, S.; Suda, H.; Yasuda, K.; Ishii, N. Impaired p53/CEP-1 is associated with lifespan extension through an age-related imbalance in the energy metabolism of C. elegans. Genes Cells 2017, 22, 1004–1010. [Google Scholar] [CrossRef]
- Wadsworth, W.G.; Riddle, D.L. Developmental regulation of energy metabolism in Caenorhabditis elegans. Dev. Biol. 1989, 132, 167–173. [Google Scholar] [CrossRef]
- Van Voorhies, W.A.; Ward, S. Broad oxygen tolerance in the nematode Caenorhabditis elegans. J. Exp. Biol. 2000, 203, 2467–2478. [Google Scholar]
- Du Plessis, S.S.; Agarwal, A.; Mohanty, G.; van der Linde, M. Oxidative phosphorylation versus glycolysis: What fuel do spermatozoa use? Asian J. Androl. 2015, 17, 230–235. [Google Scholar] [CrossRef]
- Stevens, J.F.; Revel, J.S.; Maier, C.S. Mitochondria-Centric Review of Polyphenol Bioactivity in Cancer Models. Antioxid. Redox. Signal. 2018, 29, 1589–1611. [Google Scholar] [CrossRef]
- Campbell, N.K.; Fitzgerald, H.K.; Fletcher, J.M.; Dunne, A. Plant-Derived Polyphenols Modulate Human Dendritic Cell Metabolism and Immune Function via AMPK-Dependent Induction of Heme Oxygenase-1. Front. Immunol. 2019, 10, 345. [Google Scholar] [CrossRef]
- Yan, B.; Cheng, L.; Jiang, Z.; Chen, K.; Zhou, C.; Sun, L.; Cao, J.; Qian, W.; Li, J.; Shan, T.; et al. Resveratrol Inhibits ROS-Promoted Activation and Glycolysis of Pancreatic Stellate Cells via Suppression of miR-21. Oxid Med Cell Longev. 2018, 2018, 1346958. [Google Scholar] [CrossRef]
- Tyszka-Czochara, M.; Bukowska-Strakova, K.; Kocemba-Pilarczyk, K.A.; Majka, M. Caffeic Acid Targets AMPK Signaling and Regulates Tricarboxylic Acid Cycle Anaplerosis while Metformin Downregulates HIF-1alpha-Induced Glycolytic Enzymes in Human Cervical Squamous Cell Carcinoma Lines. Nutrients 2018, 10, 841. [Google Scholar] [CrossRef]
- Rechner, A.R.; Smith, M.A.; Kuhnle, G.; Gibson, G.R.; Debnam, E.S.; Srai, S.K.S.; Moore, K.P.; Rice-Evans, C.A. Colonic metabolism of dietary polyphenols: influence of structure on microbial fermentation products. Free Radic. Biol. Med. 2004, 36, 212–225. [Google Scholar] [CrossRef]
- Rodríguez, H.; Curiel, J.A.; Landete, J.M.; de las Rivas, B.; de Felipe, F.L.; Gómez-Cordovés, C.; Mancheño, J.M.; Muñoz, R. Food phenolics and lactic acid bacteria. Int. J. Food Microbiol. 2009, 132, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Fabre, N.; Rustan, I.; de Hoffmann, E.; Quetin-Leclercq, J. Determination of flavone, flavonol, and flavanone aglycones by negative ion liquid chromatography electrospray ion trap mass spectrometry. J. Am. Soc. Mass. Spectrom. 2001, 12, 707–715. [Google Scholar] [CrossRef] [Green Version]
- Beekwilder, J.; Marcozzi, D.; Vecchi, S.; Vos, R.d.; Janssen, P.; Francke, C.; van Vlieg, J.H.; Hall, R.D. Characterization of Rhamnosidases from Lactobacillus plantarum and Lactobacillus acidophilus. Appl. Environ. Microbiol. 2009, 75, 3447–3454. [Google Scholar] [CrossRef]
- Barreca, D.; Bellocco, E.; Caristi, C.; Leuzzi, U.; Gattuso, G. Distribution of C- and O-glycosyl flavonoids, (3-hydroxy-3-methylglutaryl)glycosyl flavanones and furocoumarins in Citrus aurantium L. juice. Food Chem. 2011, 124, 576–582. [Google Scholar] [CrossRef]
- Colaric, M.; Veberic, R.; Solar, A.; Hudina, M.; Stampar, F. Phenolic acids, syringaldehyde, and juglone in fruits of different cultivars of Juglans regia L. J. Agric. Food Chem. 2005, 53, 6390–6396. [Google Scholar] [CrossRef]
- Dall’Asta, M.; Calani, L.; Tedeschi, M.; Jechiu, L.; Brighenti, F.; Del Rio, D. Identification of microbial metabolites derived from in vitro fecal fermentation of different polyphenolic food sources. Nutrition 2012, 28, 197–203. [Google Scholar] [CrossRef]
- Vitaglione, P.; Donnarumma, G.; Napolitano, A.; Galvano, F.; Gallo, A.; Scalfi, L.; Fogliano, V. Protocatechuic Acid Is the Major Human Metabolite of Cyanidin-Glucosides. J. Nutr. 2007, 137, 2043–2048. [Google Scholar] [CrossRef]
- Pereira-Caro, G.; Fernández-Quirós, B.; Ludwig, I.A.; Pradas, I.; Crozier, A.; Moreno-Rojas, J.M. Catabolism of citrus flavanones by the probiotics Bifidobacterium longum and Lactobacillus rhamnosus. Eur. J. Nutr. 2018, 57, 231–242. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Product Size (bp) | Conc. (µM) |
|---|---|---|---|
| ama-1 | 5′-ccaggaacttcggctcagta-3′ 5′-tgtatgatggtgaagctggcg-3′ | 85 | 0.1 |
| act-2 | 5′-cccactcaatccaaaggcta-3′ 5′-gggactgtgtgggraacacc-3′ | 168 | 0.1 |
| daf-16 | 5′-tcctcattcactcccgattc-3′ 5′-ccggtgtattcatgaacgtg-3′ | 175 | 0.1 |
| sir-2.1 | 5′-tggctgacgattcgatggat-3′ 5′-atgagcagaaatcgcgacac-3′ | 179 | 0.1 |
| skn-1 | 5′-acagggtggaaaaagcaagg-3′ 5′-caggccaaacgccaatgac-3′ | 246 | 0.1 |
| Peak | tR (min) | UVmax (nm) | [M−H]− | [M+H]+ | MSn m/z | Compound |
|---|---|---|---|---|---|---|
| 1 | 3.23 | 294 | 169 | MS2: 125, 151 MS3[125]: 125, 81, 97 | Gallic acid | |
| 2 | 5.37 | 260/295 | 153 | MS2:109 MS3[109]: 109 | Protocatechuic acid | |
| 3 | 5.81 | 276 | 127 | MS2: 127, 109 MS3[127]: 127, 109 | Pyrogallol | |
| 4 | 6.67 | 277 | 181 | MS2: 163, 135 MS3[163]: 119 | Dihydroxyhydrocinnamic acid | |
| 5 | 7.41 | 209/256 | 137 | MS2: 93, 137 MS3[93]: 93 | 4-Hydroxybenzoic acid | |
| 6 | 8.60 | 282 | 181 | MS2: 137, 163, 119, 109 MS3[137]: 137, 119, 109, 81, 93,95 | 3,4-Dihydroxyhydrocinnamic acid | |
| 7 | 11.47 | 272/331 | 593 | MS2: 473, 503, 353, 575 MS3[473]: 353, 383 | Apigenin 6,8-di-C-glucoside | |
| 8 | 11.71 | 218/311 | 163 | MS2: 119, 163 MS3[119]: 119 | p-Coumaric acid | |
| 9 | 12.04 | 272/349 | 623 | MS2: 503, 383, 533, 413 MS3[503]: 383, 413 | Lucenin-2 methyl ether Isomer 2 | |
| 10 | 12.41 | 272/349 | 623 | MS2: 503, 383, 533, 413 MS3[503]: 383, 413 | Lucenin-2 methyl ether Isomer 1 | |
| 11 | 13.81 | 285/330 | 595 | MS2: 287 MS3[287]: 151 | Eriodictyol-7-O-rutinoside | |
| 12 | 14.44 | 269/349 | 593 | MS2: 285 MS3[285]: 285, 151 | Kampferol rutinoside | |
| 13 | 16.46 | 260/349 | 447 | MS2: 301, 300 MS3[301]: 179, 151, 273 | Quercetin rhamnoside | |
| 14 | 16.63 | 285 | 609 | MS2: 301 MS3[301]: 286, 242, 283, 257, 125 | Hesperetin-7-O-rutinoside | |
| 15 | 18.70 | 304 | 253 | MS2: 253 MS3[253]: 253 | Daidzein | |
| 16 | 19.68 | 372 A | 285 | MS2: 285, 241, 217, 199, 175, 257, 151 MS3[285]: 241, 243, 213 | Luteolin | |
| 17 | 19.68 | 372 | 301 | MS2: 179, 151, 257, 273 MS3[179]: 151, 179 | Quercetin | |
| 18 | 20.67 | 285/330 sh | 271 | MS2: 151, 177, 165, 107 MS3[151]: 107, 151 | Naringenin | |
| 19 | 20.82 | 261/330 | 269 | MS2: 269, 201, 181, 225, 169 MS3[269]: 269 | Apigenin | |
| 20 | 21.03 | 363 | 285 | MS2: 285, 151 MS3[285]: 285 | Kaempferol | |
| 21 | 21.17 | 288/330 sh | 301 | MS2: 286, 242, 283 257, 125, 199, 151 MS3[286]: 258, 242, 199, 174, 268, 215 | Hesperetin |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dilberger, B.; Passon, M.; Asseburg, H.; Silaidos, C.V.; Schmitt, F.; Schmiedl, T.; Schieber, A.; Eckert, G.P. Polyphenols and Metabolites Enhance Survival in Rodents and Nematodes—Impact of Mitochondria. Nutrients 2019, 11, 1886. https://doi.org/10.3390/nu11081886
Dilberger B, Passon M, Asseburg H, Silaidos CV, Schmitt F, Schmiedl T, Schieber A, Eckert GP. Polyphenols and Metabolites Enhance Survival in Rodents and Nematodes—Impact of Mitochondria. Nutrients. 2019; 11(8):1886. https://doi.org/10.3390/nu11081886
Chicago/Turabian StyleDilberger, Benjamin, Maike Passon, Heike Asseburg, Carmina V. Silaidos, Fabian Schmitt, Tommy Schmiedl, Andreas Schieber, and Gunter P. Eckert. 2019. "Polyphenols and Metabolites Enhance Survival in Rodents and Nematodes—Impact of Mitochondria" Nutrients 11, no. 8: 1886. https://doi.org/10.3390/nu11081886
APA StyleDilberger, B., Passon, M., Asseburg, H., Silaidos, C. V., Schmitt, F., Schmiedl, T., Schieber, A., & Eckert, G. P. (2019). Polyphenols and Metabolites Enhance Survival in Rodents and Nematodes—Impact of Mitochondria. Nutrients, 11(8), 1886. https://doi.org/10.3390/nu11081886