Influence of Growth Hormone and Glutamine on Intestinal Stem Cells: A Narrative Review


Abstract
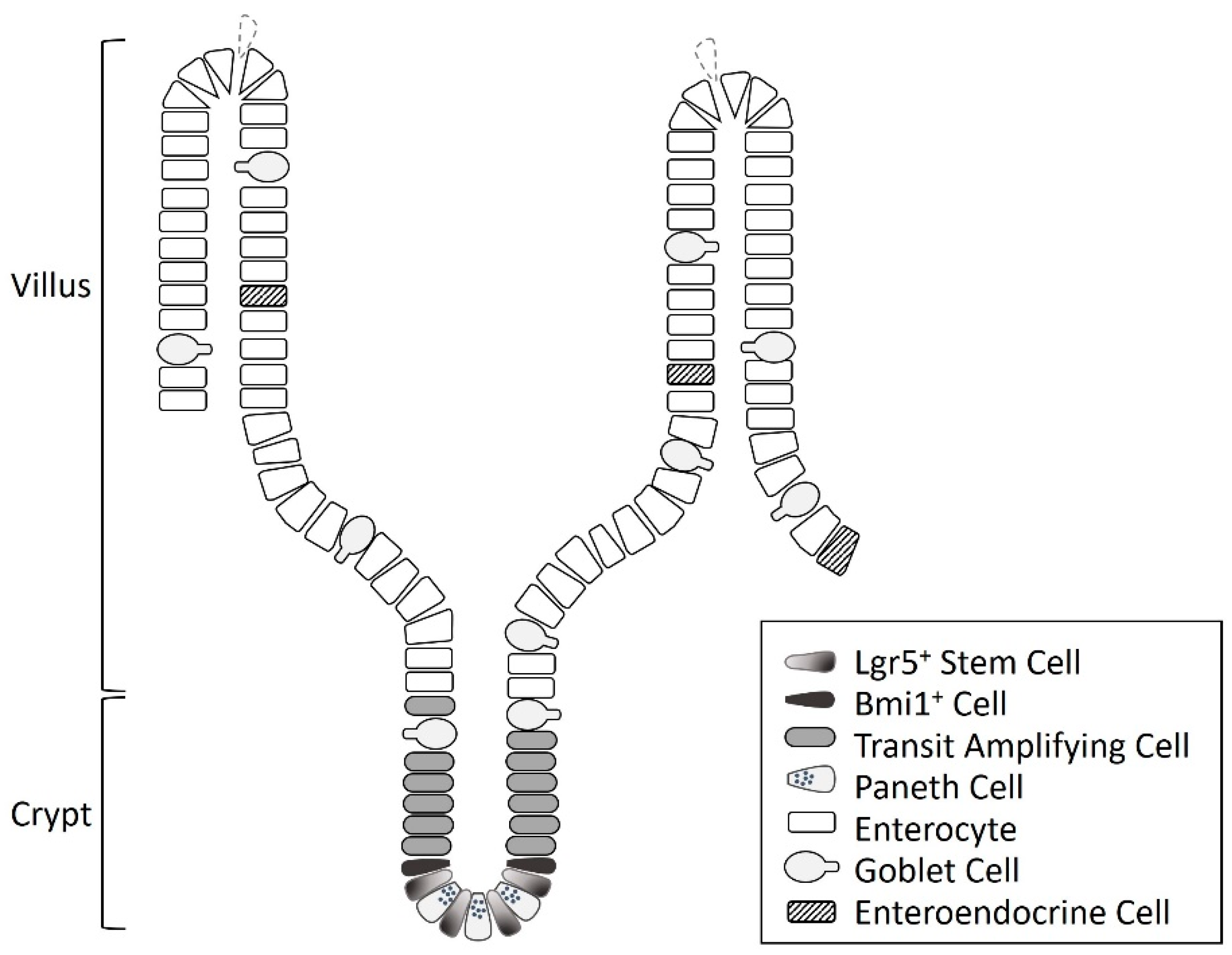
:1. Intestinal Stem Cells
2. Growth Hormone and Intestinal Stem Cells
3. Glutamine and Intestinal Stem Cells
4. Influence of Combined Growth Hormone and Glutamine on the Intestines
5. Influence of Combined Growth Hormone and Glutamine on Intestinal Stem Cells
6. Factors Affecting the Effects of Growth Hormone and/or Glutamine on the Intestines and Intestinal Stem Cells
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Barker, N.; van de Wetering, M.; Clevers, H. The intestinal stem cell. Genes Dev. 2008, 22, 1856–1864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Wu, J.M.; Lin, T.Y.; Wu, C.C.; Chiu, K.M.; Chang, B.F.; Tseng, S.H.; Chu, S.H. Tetrandrine ameliorated reperfusion injury of small bowel transplantation. J. Pediatr. Surg. 2009, 44, 2145–2152. [Google Scholar] [CrossRef] [PubMed]
- Yeung, T.M.; Chia, L.A.; Kosinski, C.M.; Kuo, C.J. Regulation of self-renewal and differentiation by the intestinal stem cell niche. Cell. Mol. Life Sci. 2011, 68, 2513–2523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mah, A.T.; van Landeghem, L.; Gavin, H.E.; Magness, S.T.; Lund, P.K. Impact of diet-induced obesity on intestinal stem cells: Hyperproliferation but impaired intrinsic function that requires insulin/IGF1. Endocrinology 2014, 155, 3302–3314. [Google Scholar] [CrossRef] [PubMed]
- McLeod, C.J.; Wang, L.; Wong, C.; Jones, D.L. Stem cell dynamics in response to nutrient availability. Curr. Biol. 2010, 20, 2100–2105. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, O.H.; Katajisto, P.; Lamming, D.W.; Gultekin, Y.; Bauer-Rowe, K.E.; Sengupta, S.; Birsoy, K.; Dursun, A.; Yilmaz, V.O.; Selig, M.; et al. mTORC1 in the Paneth cell niche couples intestinal stem-cell function to calorie intake. Nature 2012, 486, 490–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, N. Adult intestinal stem cells: Critical drivers of epithelial homeostasis and regeneration. Nat. Rev. Mol. Cell Biol. 2014, 15, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xia, Y.; Zhu, G.; Yan, J.; Tan, C.; Deng, B.; Deng, J.; Yin, Y.; Ren, W. Glutamine supplementation improves intestinal cell proliferation and stem cell differentiation in weanling mice. Food Nutr. Res. 2018, 62. [Google Scholar] [CrossRef]
- Kim, M.H.; Kim, H. The Roles of Glutamine in the Intestine and Its Implication in Intestinal Diseases. Int. J. Mol. Sci. 2017, 18, 1051. [Google Scholar] [CrossRef]
- Heuberger, J.; Kosel, F.; Qi, J.; Grossmann, K.S.; Rajewsky, K.; Birchmeier, W. Shp2/MAPK signaling controls goblet/paneth cell fate decisions in the intestine. Proc. Natl. Acad. Sci. USA 2014, 111, 3472–3477. [Google Scholar] [CrossRef] [Green Version]
- Schuijers, J.; Clevers, H. Adult mammalian stem cells: The role of Wnt, Lgr5 and R-spondins. EMBO J. 2012, 31, 2685–2696. [Google Scholar] [CrossRef] [PubMed]
- Pearce, S.C.; Al-Jawadi, A.; Kishida, K.; Yu, S.; Hu, M.; Fritzky, L.F.; Edelblum, K.L.; Gao, N.; Ferraris, R.P. Marked differences in tight junction composition and mac–Romolecular permeability among different intestinal cell types. BMC Biol. 2018, 16, 19. [Google Scholar] [CrossRef] [PubMed]
- Barker, N.; van Es, J.H.; Kuipers, J.; Kujala, P.; van den Born, M.; Cozijnsen, M.; Haegebarth, A.; Korving, J.; Begthel, H.; Peters, P.J.; et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature 2007, 449, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Basak, O.; Beumer, J.; Wiebrands, K.; Seno, H.; van Oudenaarden, A.; Clevers, H. Induced quiescence of Lgr5+ stem cells in intestinal organoids enables differentiation of hormone-producing enteroendocrine cells. Cell Stem Cell 2017, 20, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Tseng, S.H.; Yao, C.L.; Li, C.; Tsai, Y.H. Distinct Effects of Growth Hormone and Glutamine on Activation of Intestinal Stem Cells. J. Parenter. Enteral. Nutr. 2018, 42, 642–651. [Google Scholar] [CrossRef]
- Bloemendaal, A.L.; Buchs, N.C.; George, B.D.; Guy, R.J. Intestinal stem cells and intestinal homeostasis in health and in inflamma–Tion: A review. Surgery 2016, 159, 1237–1248. [Google Scholar] [CrossRef]
- Beer, V.J.; Warren, M.A.; Cope, G.H.; Baillie, H.S. Effects of growth hormone on intestinal morphology of genetically dwarf rats. J. Anat. 1995, 186, 253–259. [Google Scholar]
- Peng, H.S.; Poovaiah, N.; Forrester, M.; Cochran, E.; Wang, Q. Ex vivo culture of primary intestinal stem cells in collagen gels and foams. ACS Biomater. Sci. Eng. 2015, 1, 37–42. [Google Scholar] [CrossRef]
- Benhamou, P.H.; Canarelli, J.P.; Richard, S.; Cordonnier, C.; Postel, J.P.; Grenier, E.; Leke, A.; Dupont, C. Human recombinant growth hormone increases small bowel lengthening after massive small bowel resection in piglets. J. Pediatr. Surg. 1997, 32, 1332–1336. [Google Scholar] [CrossRef]
- Buhler, C.; Hammon, H.; Rossi, G.L.; Blum, J.W. Small intestinal morphology in eight-day-old calves fed colostrum for different durations or only milk replacer and treated with long-R3-insulin-like growth factor I and growth hormone. J. Anim. Sci. 1998, 76, 758–765. [Google Scholar] [CrossRef]
- Challacombe, D.N.; Wheeler, E.E. The trophic action of human growth hormone on human duodenal mucosa cultured in vitro. J. Pediatr. Gastroenterol. Nutr. 1995, 21, 50–53. [Google Scholar] [CrossRef]
- Guo, M.; Li, Y.; Wang, Z.; Wu, B.; Wang, J.; Li, J. Morphological adaptation in adult short bowel syndrome undergoing intestinal rehabilitation. J. Invest. Surg. 2013, 26, 1–5. [Google Scholar] [CrossRef]
- Chen, Y.; Lee, S.H.; Tsai, Y.H.; Tseng, S.H. Ischemic preconditioning increased the intestinal stem cell activities in the intestinal crypts in mice. J. Surg. Res. 2014, 187, 85–93. [Google Scholar] [CrossRef]
- Chen, Y.; Li, C.; Tsai, Y.H.; Tseng, S.H. Intestinal crypt organoid: Isolation of intestinal stem cells, In vitro culture, and optical observation. Methods Mol. Biol. 2017. [Google Scholar] [CrossRef]
- Kuo, W.T.; Lee, T.C.; Yang, H.Y.; Chen, C.Y.; Au, Y.C.; Lu, Y.Z.; Wu, L.L.; Wei, S.C.; Ni, Y.H.; Lin, B.R.; et al. PS receptor subunits have antagonistic roles in epithelial apoptosis and colonic carcinogenesis. Cell Death Differ. 2015, 22, 1590–1604. [Google Scholar] [CrossRef]
- Mohamed, M.S.; Chen, Y.; Yao, C.L. Intestinal stem cells and stem cell-based therapy for intestinal diseases. Cytotechnology 2015, 67, 177–189. [Google Scholar] [CrossRef]
- Moore, S.R.; Guedes, M.M.; Costa, T.B.; Vallance, J.; Maier, E.A.; Betz, K.J.; Aihara, E.; Mahe, M.M.; Lima, A.A.; Oria, R.B.; et al. Glutamine and alanyl-glutamine promote crypt expansion and mTOR signaling in murine enteroids. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, 831–839. [Google Scholar] [CrossRef]
- Rouch, J.D.; Scott, A.; Lei, N.Y.; Solorzano-Vargas, R.S.; Wang, J.; Hanson, E.M.; Kobayashi, M.; Lewis, M.; Stelzner, M.G.; Dunn, J.C.; et al. Development of Functional Microfold (M) Cells from Intestinal Stem Cells in Primary Human Enteroids. PLoS ONE 2016, 11, e0148216. [Google Scholar] [CrossRef]
- Wallach, T.E.; Bayrer, J.R. Intestinal Organoids: New Frontiers in the Study of Intestinal Disease and Physiology. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 180–185. [Google Scholar] [CrossRef] [Green Version]
- Cai, T.; Qi, Y.; Jergens, A.; Wannemuehler, M.; Barrett, T.A.; Wang, Q. Effects of six common dietary nutrients on murine intestinal organoid growth. PLoS ONE 2018, 13, e0191517. [Google Scholar] [CrossRef]
- Byrne, T.A.; Wilmore, D.W.; Iyer, K.; Dibaise, J.; Clancy, K.; Robinson, M.K.; Chang, P.; Gertner, J.M.; Lautz, D. Growth hormone, glutamine, and an optimal diet reduces parenteral nutrition in patients with short bowel syndrome: A prospective, randomized, placebo-controlled, double-blind clinical trial. Ann. Surg. 2005, 242, 655–661. [Google Scholar] [CrossRef]
- Matarese, L.E.; Seidner, D.L.; Steiger, E. Growth hormone, glutamine, and modified diet for intestinal adaptation. J. Am. Diet Assoc. 2004, 104, 1265–1272. [Google Scholar] [CrossRef]
- Iannoli, P.; Miller, J.H.; Ryan, C.K.; Gu, LH.; Ziegler, T.R.; Sax, H.C. Epidermal growth factor and human growth hormone accelerate adaptation after massive enterectomy in an additive, nutrient-dependent, and site-specific fashion. Surgery 1997, 122, 721–728. [Google Scholar] [CrossRef]
- Zhou, X.; Li, Y.X.; Li, N.; Li, J.S. Glutamine enhances the gut-trophic effect of growth hormone in rat after massive small bowel resection. J. Surg. Res. 2001, 99, 47–52. [Google Scholar] [CrossRef]
- Ribeiro, S.R.; Pinto, P.E.; de Miranda, A.C.; Bromberg, S.H.; Lopasso, F.P.; Irya, K. Weight loss and morphometric study of intestinal mucosa in rats after massive intestinal resection: Influence of a glutamine-enriched diet. Rev. Hosp. Clin. Fac. Med. Sao Paulo 2004, 59, 349–356. [Google Scholar] [CrossRef]
- Spadoni, J.M.; Aguilar-Nascimento, J.E.; Silva, M.H.; Spadoni-Neto, B.; Costa, P.A.; Alessio, D.M. Effects of the combined use of glutamine and growth hormone in the intestinal adaptation after massive resection of the small bowel in rats. Acta Cir. Bras. 2005, 20, 382–389. [Google Scholar] [CrossRef] [Green Version]
- Fadrique, B.; Lopez, J.M.; Bermudez, R.; Gomez de Segura, I.A.; Vazquez, I.; de Miguel, E. Growth hormone plus high protein diet promotes adaptation after massive bowel resection in aged rats. Exp. Gerontol. 2001, 36, 1727–1737. [Google Scholar] [CrossRef]
- Zhou, X.; Li, Y.X.; Li, N.; Li, J.S. Effect of bowel rehabilitative therapy on structural adaptation of remnant small intestine: Animal experiment. World J. Gastroenterol. 2001, 7, 66–73. [Google Scholar] [CrossRef]
- Shaw, D.; Gohil, K.; Basson, M.D. Intestinal mucosal atrophy and adaptation. World J. Gastroenterol. 2012, 18, 6357–6375. [Google Scholar] [CrossRef]
- Eyarefe, O.D.; Emikpe, B.O.; Akinloye, S.O.; Alonge, T.O.; Fayemi, O.E. Effects of honey, glutamine and their combination on canine small bowel epithelial cell proliferation following massive resection. Niger. J. Physiol. Sci. 2012, 27, 189–193. [Google Scholar]
- Brooks, A.J.; Waters, M.J. The growth hormone receptor: Mechanism of activation and clinical implications. Nat. Rev. Endocrinol. 2010, 6, 515–525. [Google Scholar] [CrossRef]
- Kawa, M.P.; Stecewicz, I.; Piecyk, K.; Pius-Sadowska, E.; Paczkowska, E.; Roginska, D.; Sobus, A.; Łuczkowska, K.; Gawrych, E.; Petriczko, E.; et al. Effects of growth hormone therapeutic supplementation on hematopoietic stem/progenitor cells in children with growth hormone deficiency: Focus on proliferation and differentiation capabilities. Endocrine 2015, 50, 162–175. [Google Scholar] [CrossRef]
- Waters, M.J.; Brooks, A.J. Growth hormone and cell growth. Endocr. Dev. 2012, 23, 86–95. [Google Scholar]
- Gu, Y.; Wu, Z.H. The anabolic effects of recombinant human growth hormone and glutamine on parenterally fed, short bowel rats. World J. Gastroenterol. 2002, 8, 752–757. [Google Scholar] [CrossRef]
- McLenachan, S.; Lum, M.G.; Waters, M.J.; Turnley, A.M. Growth hormone promotes proliferation of adult neurosphere cultures. Growth Horm. IGF Res. 2009, 19, 212–218. [Google Scholar] [CrossRef]
- Slot, K.A.; Kastelijn, J.; Bachelot, A.; Kelly, P.A.; Binart, N.; Teerds, K.J. Reduced recruitment and survival of primordial and growing follicles in GH receptor-deficient mice. Reproduction 2006, 131, 525–532. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhou, J.; Cheng, C.M.; Kopchick, J.J.; Bondy, C.A. Evidence supporting dual, IGF-I-independent and IGF-I-dependent, roles for GH in promoting longitudinal bone growth. J. Endocrinol. 2004, 180, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Merchav, S.; Tatarsky, I.; Hochberg, Z. Enhancement of erythropoiesis in vitro by human growth hormone is mediated by insulin-like growth factor I. Br. J. Haematol. 1988, 70, 267–271. [Google Scholar] [CrossRef]
- Merchav, S.; Tatarsky, I.; Hochberg, Z. Enhancement of human granulopoiesis in vitro by biosynthetic insulin-like growth factor I/somatomedin C and human growth hormone. J. Clin. Invest. 1988, 81, 791–797. [Google Scholar] [CrossRef]
- Welniak, L.A.; Sun, R.; Murphy, W.J. The role of growth hormone in T-cell development and reconstitution. J. Leukoc. Biol. 2002, 71, 381–387. [Google Scholar]
- Chen, B.J.; Deoliveira, D.; Spasojevic, I.; Sempowski, G.D.; Jiang, C.; Owzar, K.; Wang, X.; Gesty-Palmer, D.; Cline, J.M. Growth hormone mitigates against lethal irradiation and enhances hematologic and immune recovery in mice and nonhuman primates. PLoS ONE 2010, 5, e11056. [Google Scholar] [CrossRef]
- French, R.A.; Broussard, S.R.; Meier, W.A.; Minshall, C.; Arkins, S.; Zachary, J.F.; Dantzer, R.; Kelley, K.W. Age-associated loss of bone marrow hematopoietic cells is reversed by GH and accompanies thymic constitution. Endocrinology 2002, 143, 690–699. [Google Scholar] [CrossRef]
- Chung, J.Y.; Sunwoo, J.S.; Kim, M.W.; Kim, M. The neuroprotective effects of human growth hormone as a potential treatment for amyotrophic lateral sclerosis. Neural Regen. Res. 2015, 10, 1201–1203. [Google Scholar]
- Pathipati, P.; Gorba, T.; Scheepens, A.; Goffin, V.; Sun, Y.; Fraser, M. Growth hormone and prolactin regulate human neural stem cell regenerative activity. Neuroscience 2011, 190, 409–427. [Google Scholar] [CrossRef]
- Ulshen, M.H.; Dowling, R.H.; Fuller, C.R.; Zimmermann, E.M.; Lund, P.K. Enhanced growth of small bowel in transgenic mice overexpressing bovine growth hormone. Gastroenterology 1993, 104, 973–980. [Google Scholar] [CrossRef]
- Wheeler, E.E.; Challacombe, D.N. The trophic action of growth hormone, insulin-like growth factor-I, and insulin on human duodenal mucosa cultured in vitro. Gut 1997, 40, 57–60. [Google Scholar] [CrossRef]
- Garcia-Sancho Tellez, L.; Gomez de Segura, I.A.; Vazquez, I.; de Miguel, E.; Garcia-Sancho, L. Growth hormone effects in intestinal adaptation after massive bowel resection in the suckling rat. J. Pediatr. Gastroenterol. Nutr. 2001, 33, 477–482. [Google Scholar] [CrossRef]
- Cetinbas, F.; Yelken, B.; Gulbas, Z. Role of glutamine administration on cellular immunity after total parenteral nutrition enriched with glutamine in patients with systemic inflammatory response syndrome. J. Crit. Care 2010, 25, 661. [Google Scholar] [CrossRef]
- Venoji, R.; Amirtharaj, G.J.; Kini, A.; Vanaparthi, S.; Venkatraman, A.; Ramachandran, A. Enteral glutamine differentially regulates Nrf 2 along the villus-crypt axis of the intestine to enhance glutathione levels. J. Gastroenterol. Hepatol. 2015, 30, 1740–1747. [Google Scholar] [CrossRef]
- Zhong, X.; Zhang, X.H.; Li, X.M.; Zhou, Y.M.; Li, W.; Huang, X.X.; Zhang, L.L.; Wang, T. Intestinal growth and morphology is associated with the increase in heat shock protein 70 expression in weaning piglets through supplementation with glutamine. J. Anim. Sci. 2011, 89, 3634–3642. [Google Scholar] [CrossRef]
- Beaufrere, A.M.; Neveux, N.; Patureau Mirand, P.; Buffiere, C.; Marceau, G.; Sapin, V.; Cynober, L.; Meydinal-Denis, D. Long-term intermittent glutamine supplementation repairs intestinal damage (structure and functional mass) with advanced age: Assessment with plasma citrulline in a rodent model. J. Nutr. Health Aging 2014, 18, 814–819. [Google Scholar] [CrossRef]
- Horvath, K.; Jami, M.; Hill, I.D.; Papadimitriou, J.C.; Magder, L.S.; Chanasongcram, S. Isocaloric glutamine-free diet and the morphology and function of rat small intestine. JPEN J. Parenter. Enteral Nutr. 1996, 20, 128–134. [Google Scholar] [CrossRef]
- Scheppach, W.; Loges, C.; Bartram, P.; Christl, S.U.; Richter, F.; Dusel, G.; Stehle, P.; Fuerst, P.; Kasper, H. Effect of free glutamine and alanyl-glutamine dipeptide on mucosal proliferation of the human ileum and colon. Gastroenterology 1994, 107, 429–434. [Google Scholar] [CrossRef]
- Balteskard, L.; Unneberg, K.; Mjaaland, M.; Jenssen, T.G.; Revhaug, A. Growth hormone and insulinlike growth factor 1 promote intestinal uptake and hepatic release of glutamine in sepsis. Ann. Surg. 1998, 228, 131–139. [Google Scholar] [CrossRef]
- Tannuri, U.; Carrazza, F.R.; Iriya, K. The effects of glutamine-supplemented diet on the intestinal mucosa of the malnourished growing rat. Rev. Hosp. Clin. Fac. Med. 2000, 55, 87–92. [Google Scholar] [CrossRef]
- Zhang, W.; Frankel, W.L.; Bain, A.; Choi, D.; Klurfeld, D.M.; Rombeau, J.L. Glutamine reduces bacterial translocation after small bowel transplantation in cyclosporine-treated rats. J. Surg. Res. 1995, 58, 159–164. [Google Scholar] [CrossRef]
- Ren, W.; Duan, J.; Yin, J.; Liu, G.; Cao, Z.; Xiong, X.; Chen, S.; Li, T.; Yin, Y.; Hou, Y.; et al. Dietary L-glutamine supplementation modulates microbial community and activates innate immunity in the mouse intestine. Amino Acids 2014, 46, 2403–2413. [Google Scholar] [CrossRef]
- Ren, W.; Wang, K.; Yin, J.; Chen, S.; Liu, G.; Tan, B.; Wu, G.; Bazer, F.W.; Peng, Y.; Yin, Y. Glutamine-induced secretion of intestinal secretory immunoglobulin A: A mechanistic perspective. Front Immunol. 2016, 7, 503. [Google Scholar] [CrossRef]
- Jiang, Q.; Chen, J.; Liu, S.; Liu, G.; Yao, K.; Yin, Y. L-glutamine at-tenuates apoptosis induced by endoplasmic reticulum stress by activating the IRE1alpha-XBP1 axis in IPEC-J2: A novel mech–Anism of L-glutamine in promoting intestinal health. Int. J. Mol. Sci. 2017, 18, 2617. [Google Scholar] [CrossRef]
- Wang, B.; Wu, Z.; Ji, Y.; Sun, K.; Dai, Z.; Wu, G. L-glutamine enhances tight junction integrity by activating CaMK kinase 2-AMP-activated protein kinase signaling in intestinal porcine epithelial cells. J. Nutr. 2016, 146, 501–508. [Google Scholar] [CrossRef]
- Yi, D.; Hou, Y.; Wang, L.; Ouyang, W.; Long, M.; Zhao, D.; Ding, B.; Liu, Y.; Wu, G. L-glutamine enhances enterocyte growth via activation of the mTOR signaling pathway independently of AMPK. Amino Acids 2015, 47, 65–78. [Google Scholar] [CrossRef]
- Smith, R.J. Glutamine metabolism and its physiologic importance. J. Parenteral. Enteral. Nutr. 1990, 14, 40–44. [Google Scholar] [CrossRef]
- Swaid, F.; Sukhotnik, I.; Matter, I.; Berkowitz, D.; Hadjittofi, C.; Pollak, Y.; Lavy, A. Dietary glutamine supplementation prevents mucosal injury and modulates intestinal epithelial restitution following acetic acid induced intestinal injury in rats. Nutr. Metab. (Lond.) 2013, 10, 53. [Google Scholar] [CrossRef]
- Ueno, P.M.; Oria, R.B.; Maier, E.A.; Guedes, M.; de Azevedo, O.G.; Wu, D.; Willson, T.; Hogan, S.P.; Lima, A.A.; Guerrant, R.L.; et al. Alanyl-glutamine promotes intestinal epithelial cell homeostasis in vitro and in a murine model of weanling undernutrition. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, 612–622. [Google Scholar] [CrossRef]
- Wang, J.; Chen, L.; Li, P.; Li, X.; Zhou, H.; Wang, F.; Li, D.; Yin, Y.; Wu, G. Gene expres–sion is altered in piglet small intestine by weaning and dietary glutamine supplementation. J. Nutr. 2008, 138, 1025–1032. [Google Scholar] [CrossRef]
- Li, Y.; Xu, B.; Liu, F.; Tan, L.; Li, J. The effect of glutamine-supplemented total parenteral nutrition on nutrition and intestinal absorptive function in a rat model. Pediatr. Surg. Int. 2006, 22, 508–513. [Google Scholar] [CrossRef]
- Zhang, W.; Frankel, W.L.; Singh, A.; Laitin, E.; Klurfeld, D.; Rombeau, J.L. Improvement of structure and function in orthotopic small bowel transplantation in the rat by glutamine. Transplantation 1993, 56, 512–517. [Google Scholar] [CrossRef]
- Frankel, W.L.; Zhang, W.; Afonso, J.; Klurfeld, D.M.; Don, S.H.; Laitin, E.; Deaton, D.; Furth, E.E.; Pietra, G.G.; Naji, A.; et al. Glutamine enhancement of structure and function in transplanted small intestine in the rat. JPEN J. Parenter. Enteral Nutr. 1993, 17, 47–55. [Google Scholar] [CrossRef]
- Domeneghini, C.; Di Giancamillo, A.; Bosi, G.; Arrighi, S. Can nutraceuticals affect the structure of intestinal mucosa? Qualitative and quantitative microanatomy in L-glutamine diet-supplemented weaning piglets. Vet. Res. Commun. 2006, 30, 331–342. [Google Scholar] [CrossRef]
- Mandir, N.; Goodlad, R.A. The effects of glutamine on intestinal epithelial cell proliferation in parenterally fed rats. Gut 1999, 44, 608–614. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Ren, W.; Fang, J.; Hu, C.A.; Guan, G.; Al-Dhabi, N.A.; Yin, J.; Duraipandiyan, V.; Chen, S.; Peng, Y.; et al. L-glutamine and L-arginine protect against enterotoxigenic Escherichia coli infection via intestinal innate immunity in mice. Amino Acids 2017, 49, 1945–1954. [Google Scholar] [CrossRef]
- Zhang, X.Q.; Li, J.S.; Li, N.; Li, Y.S.; Fan, X.H. Trophic effect of enteral rehabilitative therapy in rat small bowel transplantation. Transplant. Proc. 2005, 37, 2351–2353. [Google Scholar] [CrossRef]
- Marc Rhoads, J.; Wu, G. Glutamine, arginine, and leucine signaling in the intestine. Amino Acids 2009, 37, 111–122. [Google Scholar] [CrossRef]
- Weiming, Z.; Ning, L.; Jieshou, L. Effect of recombinant human growth hormone and enteral nutrition on short bowel syndrome. J. Parenter. Enteral. Nutr. 2004, 28, 377–381. [Google Scholar] [CrossRef]
- Byrne, T.A.; Morrissey, T.B.; Nattakom, T.V.; Ziegler, T.R.; Wilmore, D.W. Growth hormone, glutamine, and a modified diet enhance nutrient absorption in patients with severe short bowel syndrome. J. Parenter. Enteral. Nutr. 1995, 19, 296–302. [Google Scholar] [CrossRef]
- Seguy, D.; Darmaun, D.; Duhamel, A.; Thuillier, F.; Cynober, L.; Cortot, A.; Gottrand, F.; Messing, B. Growth hormone enhances fat-free mass and glutamine availability in patients with short-bowel syndrome: An ancillary double-blind, randomized crossover study. Am. J. Clin. Nutr. 2014, 100, 850–858. [Google Scholar] [CrossRef]
- Darmaun, D.; Hayes, V.; Schaeffer, D.; Welch, S.; Mauras, N. Effects of glutamine and recombinant human growth hormone on protein metabolism in prepubertal children with cystic fibrosis. J. Clin. Endocrinol. Metab. 2004, 89, 1146–1152. [Google Scholar] [CrossRef]
- Donmez, R.; Oren, D.; Ozturk, G.; Kisaoglu, A.; Ozogul, B.; Atamanalp, S.S. The combined effects of glutamine and growth hormone on intestinal anastomosis in the rat intra-abdominal sepsis model. J. Surg. Res. 2013, 182, 142–145. [Google Scholar] [CrossRef]
- Duska, F.; Fric, M.; Waldauf, P.; Pazout, J.; Andel, M.; Mokrejs, P.; Tůma, P.; Pachl, J. Frequent intravenous pulses of growth hormone together with glutamine supplementation in prolonged critical illness after multiple trauma: Effects on nitrogen balance, insulin resistance, and substrate oxidation. Crit. Care Med. 2008, 36, 1707–1713. [Google Scholar] [CrossRef]
- Hammarqvist, F.; Sandgren, A.; Andersson, K.; Essen, P.; McNurlan, M.A.; Garlick, P.J.; Wernerman, J. Growth hormone together with glutamine-containing total parenteral nutrition maintains muscle glutamine levels and results in a less negative nitrogen balance after surgical trauma. Surgery 2001, 129, 576–586. [Google Scholar] [CrossRef]
- Tang, Z.F.; Ling, Y.B.; Lin, N.; Hao, Z.; Xu, R.Y. Glutamine and recombinant human growth hormone protect intestinal barrier function following portal hypertension surgery. World J. Gastroenterol. 2007, 13, 2223–2228. [Google Scholar] [CrossRef]
- Byrne, T.A.; Persinger, R.L.; Young, L.S.; Ziegler, T.R.; Wilmore, D.W. A new treatment for patients with short-bowel syndrome. Growth hormone, glutamine, and a modified diet. Ann. Surg. 1995, 222, 243–255. [Google Scholar] [CrossRef]
- Scolapio, J.S.; Camilleri, M.; Fleming, C.R.; Oenning, L.V.; Burton, D.D.; Sebo, T.J.; Batts, K.P.; Kelly, D.G. Effect of growth hormone, glutamine, and diet on adaptation in short-bowel syndrome: A randomized, controlled study. Gastroenterology 1997, 113, 1074–1081. [Google Scholar] [CrossRef]
- Jung, S.E.; Youn, Y.K.; Lim, Y.S.; Song, H.G.; Rhee, J.E.; Suh, G.J. Combined administration of glutamine and growth hormone synergistically reduces bacterial translocation in sepsis. J. Korean Med. Sci. 2003, 18, 17–22. [Google Scholar] [CrossRef]
- Wales, P.W.; Nasr, A.; de Silva, N.; Yamada, J. Human growth hormone and glutamine for patients with short bowel syndrome. Cochrane Database Syst. Rev. 2010, 6, CD006321. [Google Scholar] [CrossRef]
- Gu, Y.; Wu, Z.H.; Xie, J.X.; Jin, D.Y.; Zhuo, H.C. Effects of growth hormone (rhGH) and glutamine supplemented parenteral nutrition on intestinal adaptation in short bowel rats. Clin. Nutr. 2001, 20, 159–166. [Google Scholar] [CrossRef]
- Waitzberg, D.L.; Cukier, C.; Mucerino, D.R.; Logulo, A.F.; Torrinhas, R.S.; de Castro, I. Small bowel adaptation with growth hormone and glutamine after massive resection of rat’s small bowel. Nutr. Hosp. 1999, 14, 81–90. [Google Scholar]
- Zhou, Y.; Wu, X.T.; Yang, G.; Zhuang, W.; Wei, M.L. Clinical evidence of growth hormone, glutamine and a modified diet for short bowel syndrome: Meta-analysis of clinical trials. Asia Pac. J. Clin. Nutr. 2005, 14, 98–102. [Google Scholar]
- Vanderhoof, J.A.; Kollman, K.A.; Griffin, S.; Adrian, T.E. Growth hormone and glutamine do not stimulate intestinal adaptation following massive small bowel resection in the rat. J. Pediatr. Gastroenterol. Nutr. 1997, 25, 327–331. [Google Scholar] [CrossRef]
- Seguy, D.; Vahedi, K.; Kapel, N.; Souberbielle, J.C.; Messing, B. Low-dose growth hormone in adult home parenteral nutrition-dependent short bowel syndrome patients: A positive study. Gastroenterology 2003, 124, 293–302. [Google Scholar] [CrossRef]
- Szkudlarek, J.; Jeppesen, P.B.; Mortensen, P.B. Effect of high dose growth hormone with glutamine and no change in diet on intestinal absorption in short bowel patients: A randomised, double blind, crossover, placebo controlled study. Gut 2000, 47, 199–205. [Google Scholar] [CrossRef]
- Weale, A.R.; Edwards, A.G.; Bailey, M.; Lear, P.A. Intestinal adaptation after massive intestinal resection. Postgrad Med. J. 2005, 81, 178–184. [Google Scholar] [CrossRef] [Green Version]
- Gibson, P.R.; Anderson, R.P.; Mariadason, J.M.; Wilson, A.J. Protective role of the epithelium of the small intestine and colon. Inflamm. Bowel Dis. 1996, 2, 279–302. [Google Scholar] [CrossRef]


{kind=link}
| Model | GH Doses | Treatment Duration | Effects | Reference Number |
|---|---|---|---|---|
| Cultured ISCs from mice | 10 ng/ml | 7 days | ↑Lgr5, Bmi1, Msi1, EphB3, Ki67 ↑lysozyme, villin, ↓chromogranin A  mucin 2 mucin 2 ↑crypt organoids | [15] |
| Cultured ISCs from GH-treated mice | 5 mg/kg/day, sc | 7 days | ↑Lgr5, Msi1, EphB3 Bmi 1 ↑lysozyme, villin ↓mucin 2 chromogranin A ↑crypt organoids | [15] |
| Rats genetically deficient in GH | 1.6 IU/day, ip | 7 days | ↑villus volume, surface area, crypt volume, epithelial cell height | [18] |
| Newborn piglets with 80% intestinal resection | 0.1 IU/kg/day, sc | 28 days | ↑intestinal lengtheningvillus height, villus diameter, intestinal muscular layer, wall thickness, mitosis number per filed, crypt/villus ratio | [19] |
| Newborn calves | 1 mg/kg, bid, sc | 7 days | ↓villus size ↑crypt depth, crypt/villus ratio | [20] |
| Cultured explants of human duodenal mucosa | 0.004 IU/ml | 22 h | ↑number of metaphase arrests per crypt | [21] |
| Aged rats with 80% intestinal resection | 1 mg/kg/day, sc + high protein diet | 7 days | ↑mucosal height, crypt proliferation | [37] |
| Cultured explants of human duodenal mucosa | 0.004 IU/ml | 1 day | ↑crypt cell proliferation | [56] |
| Suckling rats with 80% intestinal resection | 1 mg/kg, sc, qod | 16 days (8 doses in total) | ↑intestinal lengthening, ↑crypt height, mucosa mass slightly | [57] |
| Model | Gln Doses | Treatment Duration | Effects | Reference Number |
|---|---|---|---|---|
| 3-week-old weaning mice, ileum | 10 mg/mL + basal diet | 14 days | ↓crypt depth ↔villus height ↑villus/crypt ratio ↑Ki67-positive cells in the crypt ↔sucrase, lysozyme, angiogenin 4, mucin 2, trefoil factor 3, peptide YY, chromogranin A, | [8] |
| Cultured ISCs from mice | 10 mM | 7 days | ↑Msi1 ↔Lgr5, Bmi1, EphB3 ↑mucin 2, chromogranin A ↔lysozyme, villin ↑crypt organoids | [15] |
| Cultured ISCs from Gln-treated mice | 1 g/kg/day, ip | 7 days | ↑Lgr5 expression ↔Bmi1, Msi 1, EphB3 ↑mucin 2 ↔villin, chromogranin A ↓lysozyme | [15] |
| Midjejunal crypts of mice | 2 mM | 1–4 days | ↑epithelial cell proliferation, enteroid expansion | [27] |
| Rats with massive intestinal resection (25 cm jejunum remnant) | 12% in diet | 20 days | ↑villus height, crypt depth, mucosal thickness | [35] |
| Dogs with 70% intestinal resection | 33 g/5 kg/day, oral | 15 days | ↑villus height and width, crypt depth | [40] |
| Rats | 4% or 8% in diet | 28 days | ↑mucosal weight, protein and DNA content, mitosis number per crypt ↓villus height, villus/crypt ratio | [62] |
| Biopsy samples from normal human ileum | Incubation with 2 mM Gln | 4 h | ↑crypt cell proliferation, BrdU labeling in the crypt | [63] |
| Rats with 15 days of malnutrition, jejunum | 2% in diet | 15 days | Correction of malnutrition-induced increased crypt depth | [65] |
| Rats with 60% intestinal resection + allograft transplantation | 2% + isocaloric polymeric diet, infusion via gastrostomy | 10 days | ↑crypt depth | [66] |
| Weaning piglets | 2% in diet | 28 days | ↑villus height, crypt depth ↓villus/crypt ratio ↑PCNA staining in crypt cells, number of mitotic mucosal cells, Lgr5 mRNA | [79] |
| Rats | 0.5%, 1%, or 2% in TPN | 10 days | ↔mitotic activity in the crypts, BrdU labeling | [80] |
| Model | GH and Gln Doses | Treatment Duration | Effects | Reference Number |
|---|---|---|---|---|
| Cultured ISCs from mice | GH: 10 ng/mL Gln: 10 mM | 7 days | ↔organoid formation ↑Lgr5, Bmi1, Msi1, EphB3 ↑mucin 2 ↔lysozyme, villin, chromogranin A | [15] |
| Cultured ISCs from GH and Gln-treated mice | GH: 5 mg/kg/day, sc Gln: 1 g/kg/day, ip | 7 days | ↑organoid formation ↑Ki67 staining ↑Lgr5, Bmi1, Msi1, EphB3 ↔lysozyme, villin, mucin 2, chromogranin A | [15] |
| Humans with short bowel syndrome | GH: 0.05 mg/kg/day, sc Gln: 30 g/day, enteral | 28 days | ↑crypt depth, ↑villus height ↑Ki67 staining | [22] |
| Rats with 80% intestinal resection | GH: 0.6 IU/day (2 g/day), sc Gln: 4%, enteral | 14 days | ↓crypt depth, ↔villus height ↓intestinal wall width | [36] |
| Rats with 85% intestinal resection | GH: 0.3 IU, bid, sc Gln: 20 g/l, enteral + soybean fiber | 8 days | ↑crypt depth, ↑villus height ↑mucosal thickness | [38] |
| Rats with allogenic heterotopic small bowel transplantation | GH: 1 U/kg/day, sc Gln: 2 g/100 mL parenteral solution | 14 days | ↑crypt depth, ↑villus height ↑villus width | [82] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Tsai, Y.-H.; Tseng, B.-J.; Tseng, S.-H. Influence of Growth Hormone and Glutamine on Intestinal Stem Cells: A Narrative Review. Nutrients 2019, 11, 1941. https://doi.org/10.3390/nu11081941
Chen Y, Tsai Y-H, Tseng B-J, Tseng S-H. Influence of Growth Hormone and Glutamine on Intestinal Stem Cells: A Narrative Review. Nutrients. 2019; 11(8):1941. https://doi.org/10.3390/nu11081941
Chicago/Turabian StyleChen, Yun, Ya-Hui Tsai, Bor-Jiun Tseng, and Sheng-Hong Tseng. 2019. "Influence of Growth Hormone and Glutamine on Intestinal Stem Cells: A Narrative Review" Nutrients 11, no. 8: 1941. https://doi.org/10.3390/nu11081941
APA StyleChen, Y., Tsai, Y. -H., Tseng, B. -J., & Tseng, S. -H. (2019). Influence of Growth Hormone and Glutamine on Intestinal Stem Cells: A Narrative Review. Nutrients, 11(8), 1941. https://doi.org/10.3390/nu11081941