Maternal Creatine Supplementation Positively Affects Male Rat Hippocampal Synaptic Plasticity in Adult Offspring

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Supplementation Protocol
2.2. Electrophysiological Experiment Preparation
2.3. Field Potential Recordings
2.4. Patch-Clamp Recordings
2.5. Morphological Analysis
2.6. Calcium Imaging
2.7. Statistical Analysis
3. Results
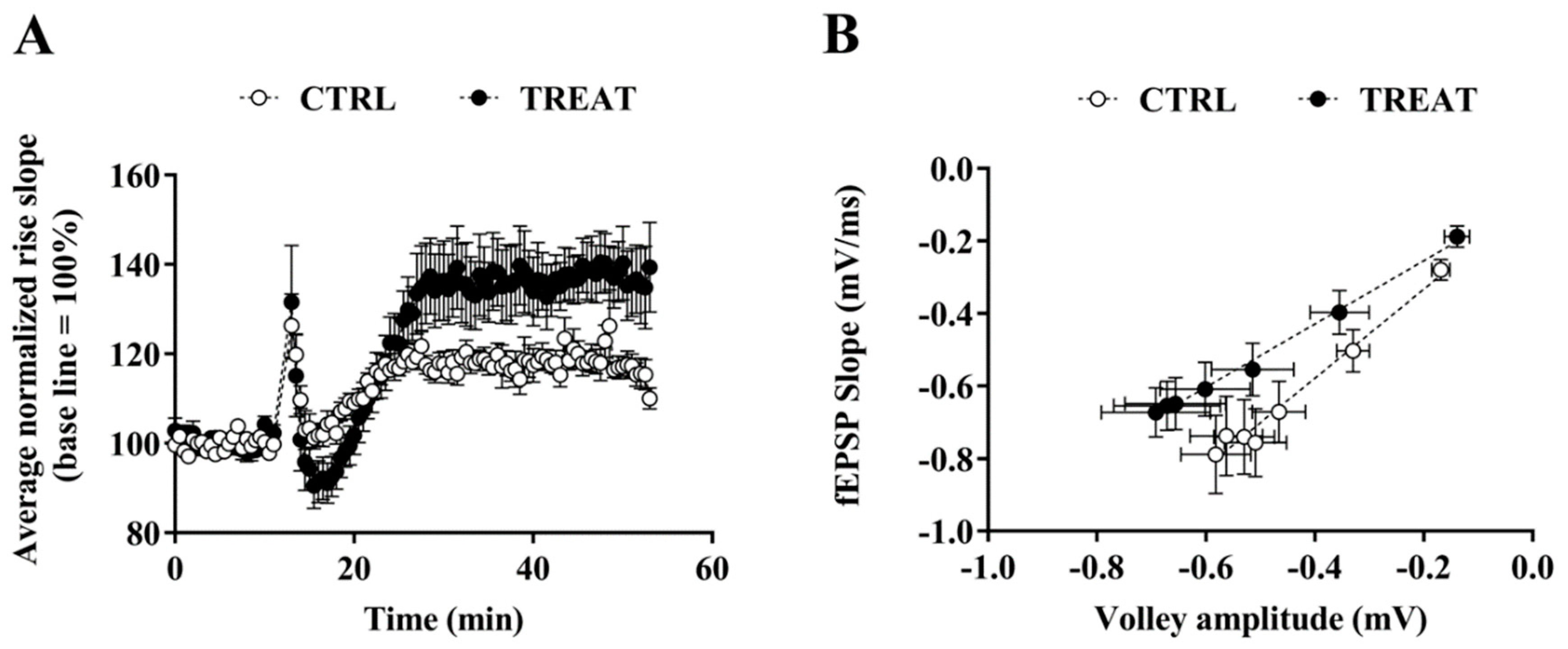
3.1. Field Recordings
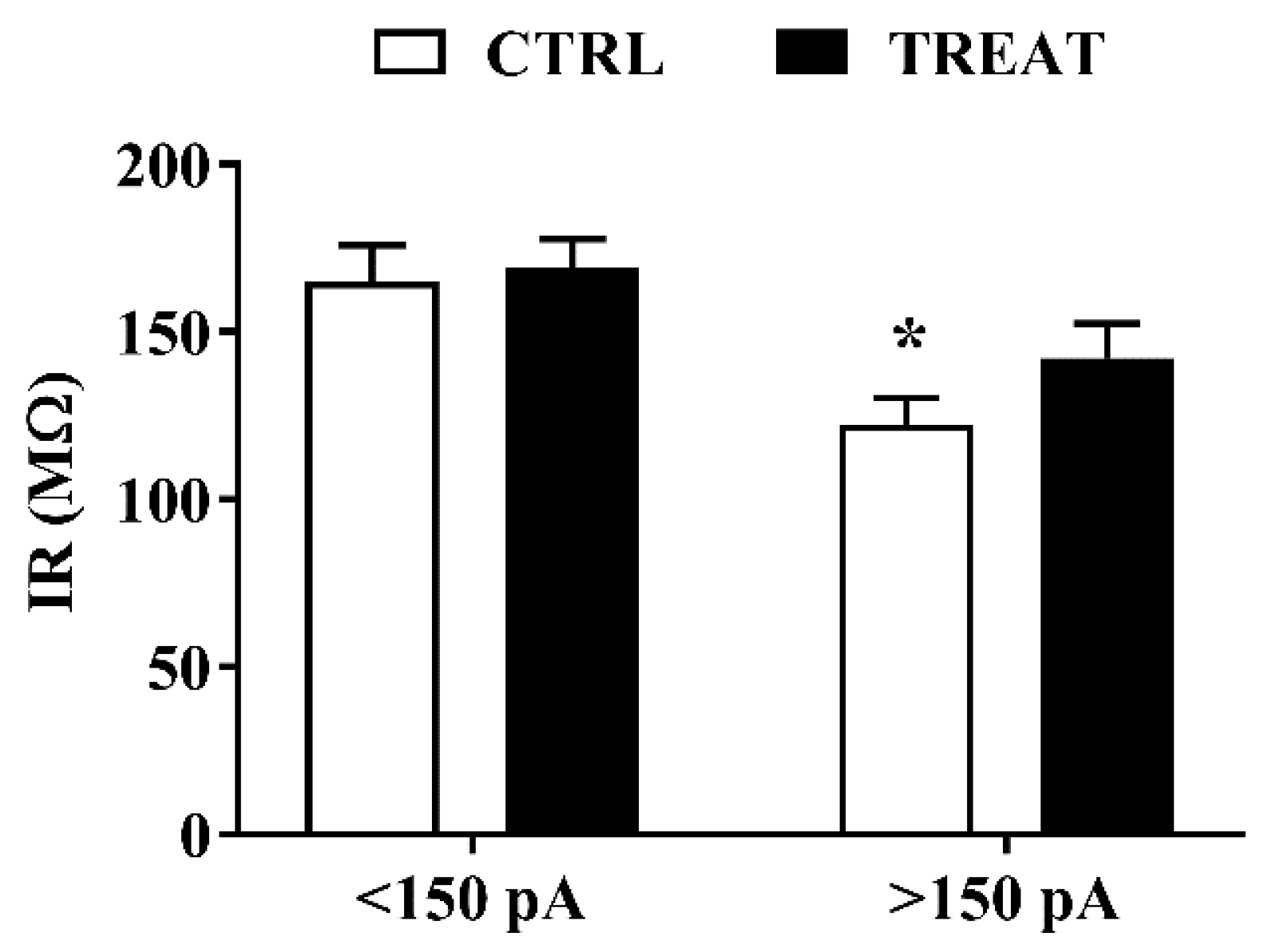
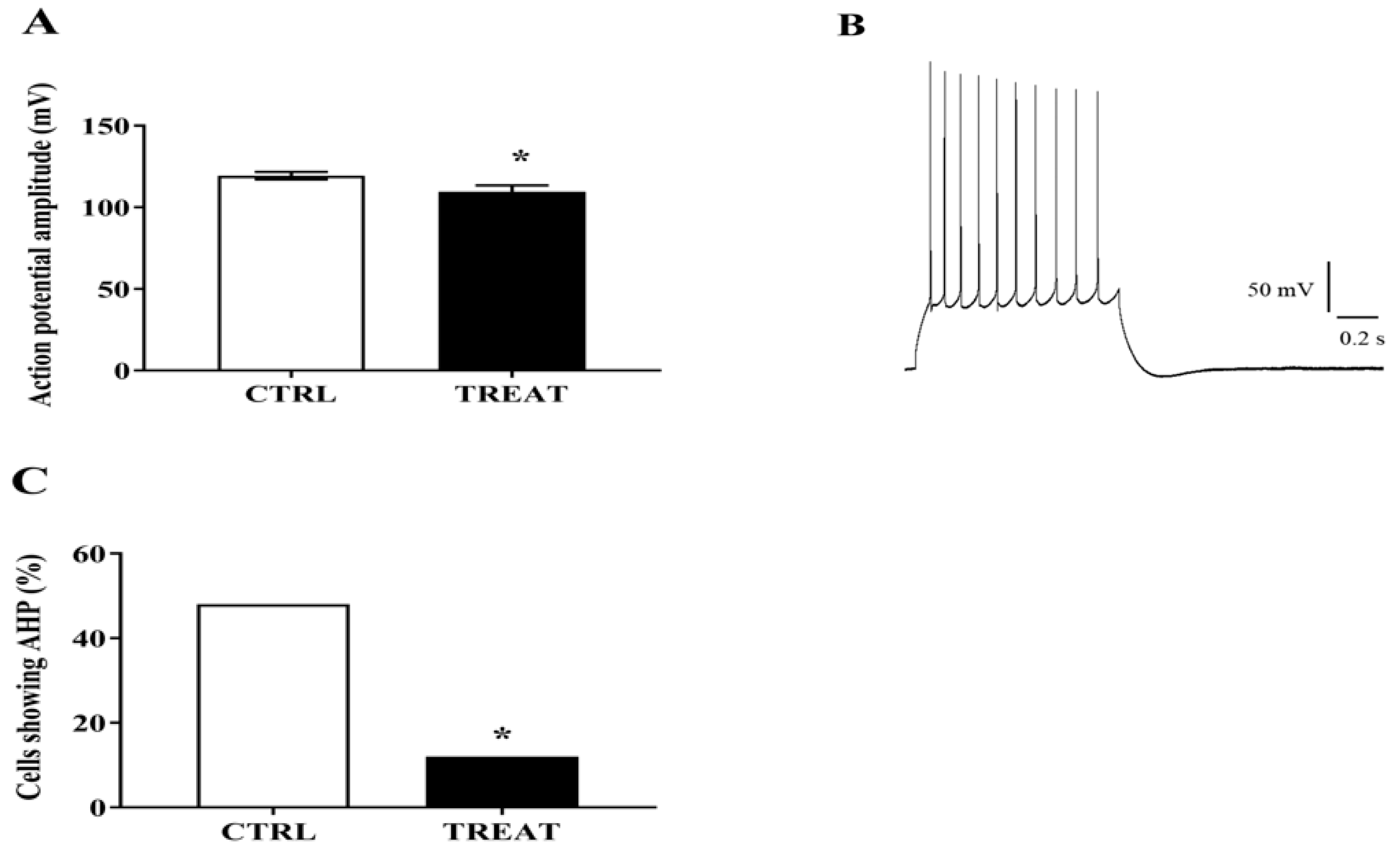
3.2. Whole-Cell Analysis
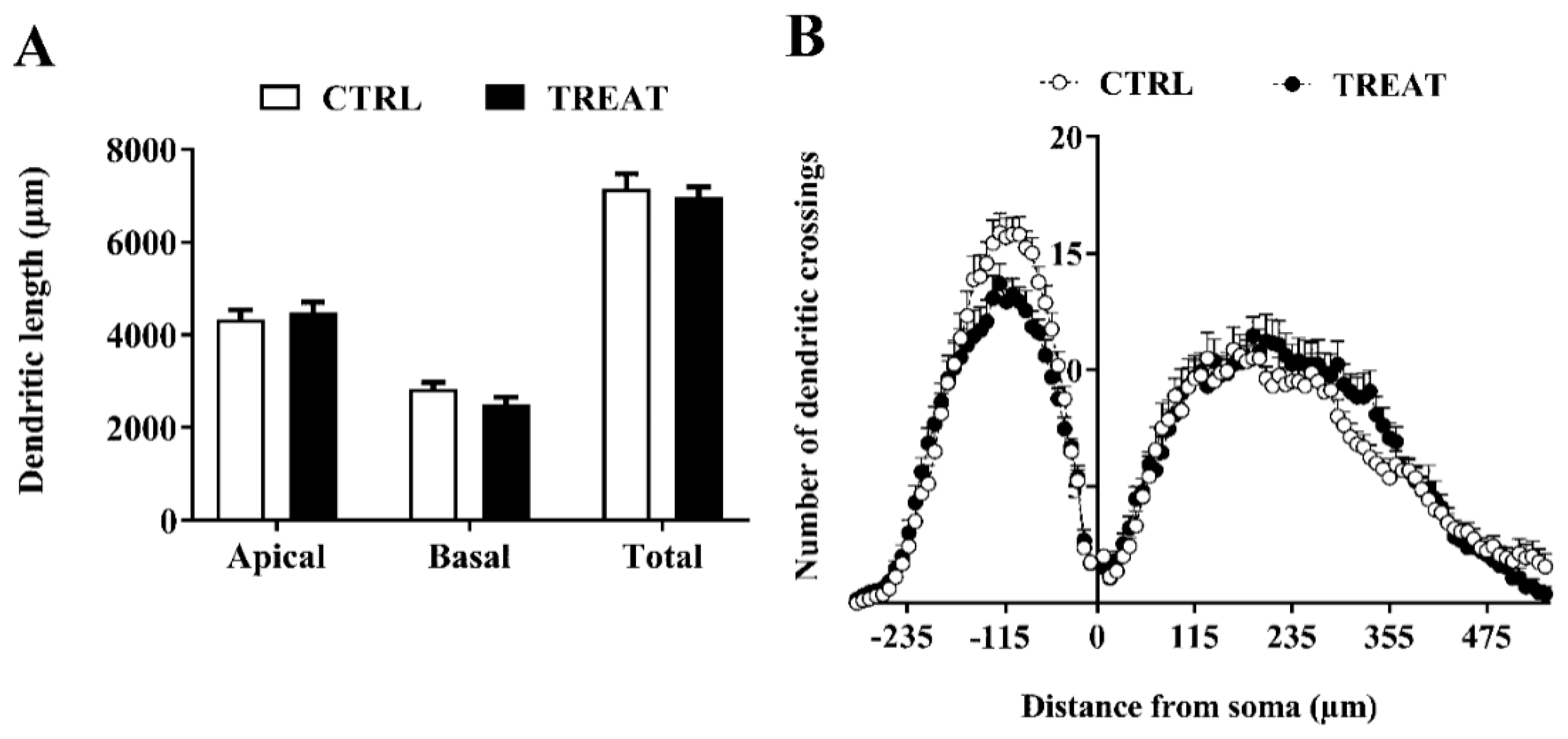
3.3. Morphological Analysis
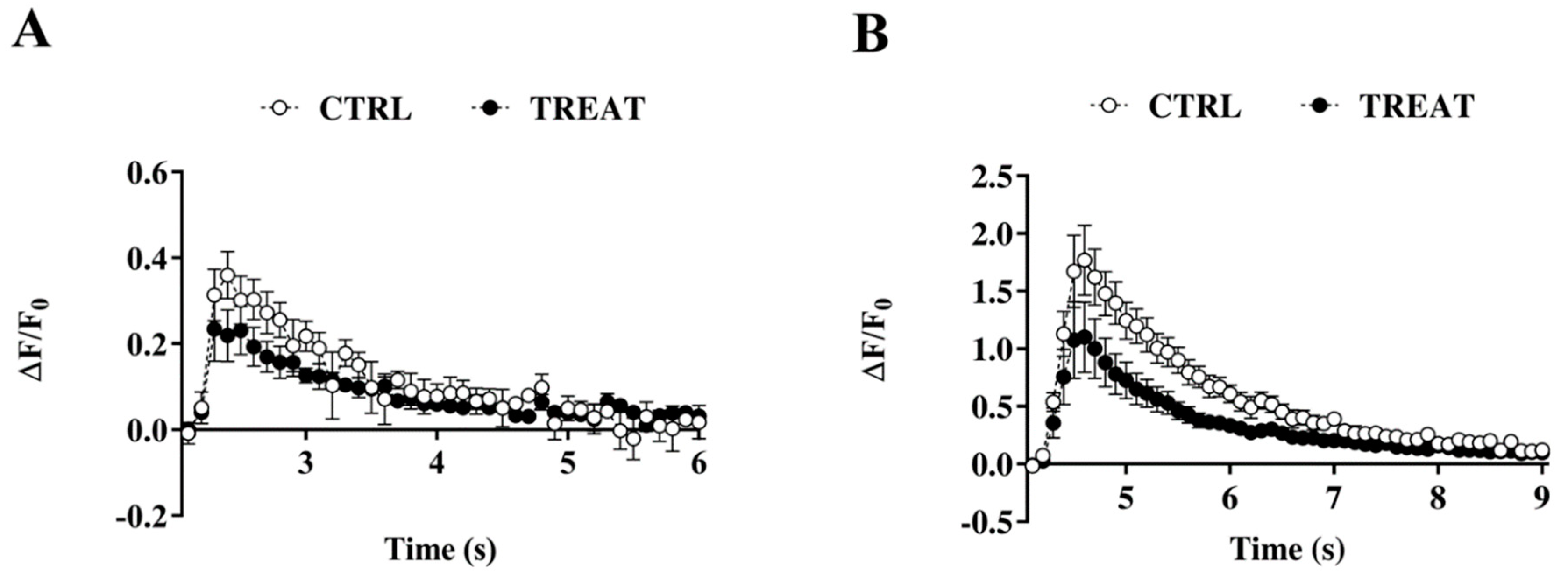
3.4. Intracellular Calcium Dynamics
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chung, Y.L.; Alexanderson, H.; Pipitone, N.; Morrison, C.; Dastmalchi, M.; Stahl-Hallengren, C.; Richards, S.; Thomas, E.L.; Hamilton, G.; Bell, J.D.; et al. Creatine supplements in patients with idiopathic inflammatory myopathies who are clinically weak after conventional pharmacologic treatment: Six-month, double-blind, randomized, placebo-controlled trial. Arthritis Rheum. 2007, 57, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Kley, R.A.; Tarnopolsky, M.A.; Vorgerd, M. Creatine treatment in muscle disorders: A meta-analysis of randomised controlled trials. J. Neurol. Neurosurg. Psychiatry 2008, 79, 366–367. [Google Scholar] [CrossRef] [PubMed]
- Candow, D.G.; Chilibeck, P.D.; Forbes, S.C. Creatine supplementation and aging musculoskeletal health. Endocrine 2014, 45, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Forbes, S.C.; Chilibeck, P.D.; Candow, D.G. Creatine Supplementation During Resistance Training Does Not Lead to Greater Bone Mineral Density in Older Humans: A Brief Meta-Analysis. Front. Nutr. 2018, 5, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze, A.; Battini, R. Pre-symptomatic treatment of creatine biosynthesis defects. Sub-Cell. Biochem. 2007, 46, 167–181. [Google Scholar]
- Joncquel-Chevalier Curt, M.; Voicu, P.M.; Fontaine, M.; Dessein, A.F.; Porchet, N.; Mention-Mulliez, K.; Dobbelaere, D.; Soto-Ares, G.; Cheillan, D.; Vamecq, J. Creatine biosynthesis and transport in health and disease. Biochimie 2015, 119, 146–165. [Google Scholar] [CrossRef]
- Bruun, T.U.J.; Sidky, S.; Bandeira, A.O.; Debray, F.G.; Ficicioglu, C.; Goldstein, J.; Joost, K.; Koeberl, D.D.; Luisa, D.; Nassogne, M.C.; et al. Treatment outcome of creatine transporter deficiency: International retrospective cohort study. Metab. Brain Dis. 2018, 33, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Avgerinos, K.I.; Spyrou, N.; Bougioukas, K.I.; Kapogiannis, D. Effects of creatine supplementation on cognitive function of healthy individuals: A systematic review of randomized controlled trials. Exp. Gerontol. 2018, 108, 166–173. [Google Scholar] [CrossRef]
- Beard, E.; Braissant, O. Synthesis and transport of creatine in the CNS: Importance for cerebral functions. J. Neurochem. 2010, 115, 297–313. [Google Scholar] [CrossRef]
- Ek, C.J.; Dziegielewska, K.M.; Habgood, M.D.; Saunders, N.R. Barriers in the developing brain and Neurotoxicology. Neurotoxicology 2012, 33, 586–604. [Google Scholar] [CrossRef]
- Sestili, P.; Ambrogini, P.; Barbieri, E.; Sartini, S.; Fimognari, C.; Calcabrini, C.; Diaz, A.R.; Guescini, M.; Polidori, E.; Luchetti, F.; et al. New insights into the trophic and cytoprotective effects of creatine in in vitro and in vivo models of cell maturation. Amino Acids 2016, 48, 1897–1911. [Google Scholar] [CrossRef]
- Brustovetsky, N.; Brustovetsky, T.; Dubinsky, J.M. On the mechanisms of neuroprotection by creatine and phosphocreatine. J. Neurochem. 2001, 76, 425–434. [Google Scholar] [CrossRef]
- Miller, R.K.; Berndt, W.O. Characterization of neutral amino acid accumulation by human term placental slices. Am. J. Physiol. 1974, 227, 1236–1242. [Google Scholar] [CrossRef] [Green Version]
- Wallimann, T.; Tokarska-Schlattner, M.; Schlattner, U. The creatine kinase system and pleiotropic effects of creatine. Amino Acids 2011, 40, 1271–1296. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, H.; Ellery, S.; Ireland, Z.; LaRosa, D.; Snow, R.; Walker, D.W. Creatine supplementation during pregnancy: Summary of experimental studies suggesting a treatment to improve fetal and neonatal morbidity and reduce mortality in high-risk human pregnancy. BMC Pregnancy Childbirth 2014, 14. [Google Scholar] [CrossRef]
- Ireland, Z.; Castillo-Melendez, M.; Dickinson, H.; Snow, R.; Walker, D.W. A maternal diet supplemented with creatine from mid-pregnancy protects the newborn spiny mouse brain from birth hypoxia. Neuroscience 2011, 194, 372–379. [Google Scholar] [CrossRef]
- Sartini, S.; Lattanzi, D.; Ambrogini, P.; Di Palma, M.; Galati, C.; Savelli, D.; Polidori, E.; Calcabrini, C.; Rocchi, M.B.; Sestili, P.; et al. Maternal creatine supplementation affects the morpho-functional development of hippocampal neurons in rat offspring. Neuroscience 2016, 312, 120–129. [Google Scholar] [CrossRef]
- Gerbatin, R.R.; Silva, L.F.A.; Hoffmann, M.S.; Della-Pace, I.D.; do Nascimento, P.S.; Kegler, A.; de Zorzi, V.N.; Cunha, J.M.; Botelho, P.; Neto, J.B.T.; et al. Delayed creatine supplementation counteracts reduction of GABAergic function and protects against seizures susceptibility after traumatic brain injury in rats. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 92, 328–338. [Google Scholar] [CrossRef]
- Ipsiroglu, O.S.; Stromberger, C.; Ilas, J.; Höger, H.; Mühl, A.; Stöckler-Ipsiroglu, S. Changes of tissue creatine concentrations upon oral supplementation of creatine-monohydrate in various animal species. Life Sci. 2001, 69, 1805–1815. [Google Scholar] [CrossRef]
- Braissant, O.; Henry, H.; Villard, A.M.; Speer, O.; Wallimann, T.; Bachmann, C. Creatine synthesis and transport during rat embryogenesis: Spatiotemporal expression of AGAT, GAMT and CT1. BMC Dev. Biol. 2005, 5. [Google Scholar] [CrossRef]
- Ambrogini, P.; Lattanzi, D.; Ciuffoli, S.; Agostini, D.; Bertini, L.; Stocchi, V.; Santi, S.; Cuppini, R. Morpho-functional characterization of neuronal cells at different stages of maturation in granule cell layer of adult rat dentate gyrus. Brain Res. 2004, 1017, 21–31. [Google Scholar] [CrossRef]
- Betti, M.; Ambrogini, P.; Minelli, A.; Floridi, A.; Lattanzi, D.; Ciuffoli, S.; Bucherelli, C.; Prospero, E.; Frontini, A.; Santarelli, L.; et al. Maternal dietary loads of α-tocopherol depress protein kinase C signaling and synaptic plasticity in rat postnatal developing hippocampus and promote permanent deficits in adult offspring. J. Nutr. Biochem. 2011, 22, 60–70. [Google Scholar] [CrossRef]
- Lattanzi, D.; Savelli, D.; Di Palma, M.; Sartini, S.; Eusebi, S.; Borroto-Escuela, D.O.; Cuppini, R.; Fuxe, K.; Ambrogini, P. Electrophysiological approach to GPCR–RTK interaction study in hippocampus of adult rats. In Neuromethods; Humana Press: Clifton, NJ, USA, 2018; Volume 140, pp. 71–90. [Google Scholar]
- Maravall, M.; Mainen, Z.F.; Sabatini, B.L.; Svoboda, K. Estimating intracellular calcium concentrations and buffering without wavelength ratioing. Biophys. J. 2000, 78, 2655–2667. [Google Scholar] [CrossRef]
- Kumar, A.; Foster, T.C. Enhanced long-term potentiation during aging is masked by processes involving intracellular calcium stores. J. Neurophysiol. 2004, 91, 2437–2444. [Google Scholar] [CrossRef]
- Sartini, S.; Sestili, P.; Colombo, E.; Martinelli, C.; Bartolini, F.; Ciuffoli, S.; Lattanzi, D.; Sisti, D.; Cuppini, R. Creatine affects in vitro electrophysiological maturation of neuroblasts and protects them from oxidative stress. J. Neurosci. Res. 2012, 90, 435–446. [Google Scholar] [CrossRef]
- Altman, J.; Bayer, S.A. Prolonged sojourn of developing pyramidal cells in the intermediate zone of the hippocampus and their settling in the stratum pyramidale. J. Comp. Neurol. 1990, 301, 343–364. [Google Scholar] [CrossRef]
- Bayer, S.A. Development of the hippocampal region in the rat. I. Neurogenesis examined with 3H-thymidine autoradiography. J. Comp. Neurol. 1980, 190, 87–114. [Google Scholar] [CrossRef]
- Fukumitsu, K.; Fujishima, K.; Yoshimura, A.; Wu, Y.K.; Heuser, J.; Kengaku, M. Synergistic action of dendritic mitochondria and creatine kinase maintains ATP homeostasis and actin dynamics in growing neuronal dendrites. J. Neurosci. 2015, 35, 5707–5723. [Google Scholar] [CrossRef]
- Giglio, A.M.; Storm, J.F. Postnatal development of temporal integration, spike timing and spike threshold regulation by a dendrotoxin-sensitive K+ current in rat CA1 hippocampal cells. Eur. J. Neurosci. 2014, 39, 12–23. [Google Scholar] [CrossRef]
- Springer, S.J.; Burkett, B.J.; Schrader, L.A. Modulation of BK channels contributes to activity-dependent increase of excitability through MTORC1 activity in CA1 pyramidal cells of mouse hippocampus. Front. Cell. Neurosci. 2014, 8, 451. [Google Scholar] [CrossRef]
- Borde, M.; Bonansco, C.; Fernandez de Sevilla, D.; Le Ray, D.; Buno, W. Voltage-clamp analysis of the potentiation of the slow Ca2+-activated K+ current in hippocampal pyramidal neurons. Hippocampus 2000, 10, 198–206. [Google Scholar] [CrossRef]
- Kumar, A.; Foster, T.C. 17beta-estradiol benzoate decreases the AHP amplitude in CA1 pyramidal neurons. J. Neurophysiol. 2002, 88, 621–626. [Google Scholar] [CrossRef]
- Landfield, P.W.; Pitler, T.A. Prolonged Ca2+-dependent afterhyperpolarizations in hippocampal neurons of aged rats. Science 1984, 226, 1089–1092. [Google Scholar] [CrossRef]
- Power, J.M.; Wu, W.W.; Sametsky, E.; Oh, M.M.; Disterhoft, J.F. Age-related enhancement of the slow outward calcium-activated potassium current in hippocampal CA1 pyramidal neurons in vitro. J. Neurosci. Off. J. Soc. Neurosci. 2002, 22, 7234–7243. [Google Scholar] [CrossRef]
- Sah, P.; Faber, E.S. Channels underlying neuronal calcium-activated potassium currents. Prog. Neurobiol. 2002, 66, 345–353. [Google Scholar] [CrossRef]
- Sah, P.; Bekkers, J.M. Apical dendritic location of slow afterhyperpolarization current in hippocampal pyramidal neurons: Implications for the integration of long-term potentiation. J. Neurosci. Off. J. Soc. Neurosci. 1996, 16, 4537–4542. [Google Scholar] [CrossRef]
- Norris, C.M.; Halpain, S.; Foster, T.C. Reversal of age-related alterations in synaptic plasticity by blockade of L-type Ca2+ channels. J. Neurosci. Off. J. Soc. Neurosci. 1998, 18, 3171–3179. [Google Scholar] [CrossRef]
- Cohen, A.S.; Coussens, C.M.; Raymond, C.R.; Abraham, W.C. Long-lasting increase in cellular excitability associated with the priming of LTP induction in rat hippocampus. J. Neurophysiol. 1999, 82, 3139–3148. [Google Scholar] [CrossRef]
- Sourdet, V.; Russier, M.; Daoudal, G.; Ankri, N.; Debanne, D. Long-term enhancement of neuronal excitability and temporal fidelity mediated by metabotropic glutamate receptor subtype 5. J. Neurosci. Off. J. Soc. Neurosci. 2003, 23, 10238–10248. [Google Scholar] [CrossRef]
- Foster, T.C.; Norris, C.M. Age-associated changes in Ca2+-dependent processes: Relation to hippocampal synaptic plasticity. Hippocampus 1997, 7, 602–612. [Google Scholar] [CrossRef]
- Foster, T.C. Involvement of hippocampal synaptic plasticity in age-related memory decline. Brain Res. Brain Res. Rev. 1999, 30, 236–249. [Google Scholar] [CrossRef]
- Buchanan, K.A.; Mellor, J.R. The development of synaptic plasticity induction rules and the requirement for postsynaptic spikes in rat hippocampal CA1 pyramidal neurones. J. Physiol. 2007, 585, 429–445. [Google Scholar] [CrossRef]
- Bliss, T.V.; Collingridge, G.L. A synaptic model of memory: Long-term potentiation in the hippocampus. Nature 1993, 361, 31–39. [Google Scholar] [CrossRef]
- Malenka, R.C.; Bear, M.F. LTP and LTD: An embarrassment of riches. Neuron 2004, 44, 5–21. [Google Scholar] [CrossRef]
- Lynch, M.A. Long-term potentiation and memory. Physiol. Rev. 2004, 84, 87–136. [Google Scholar] [CrossRef]
- Udobi, K.C.; Delcimmuto, N.; Kokenge, A.N.; Abdulla, Z.I.; Perna, M.K.; Skelton, M.R. Deletion of the creatine transporter gene in neonatal, but not adult, mice leads to cognitive deficits. J. Inherit. Metab. Dis. 2019. [Google Scholar] [CrossRef]
- Koga, Y.; Takahashi, H.; Oikawa, D.; Tachibana, T.; Denbow, D.M.; Furuse, M. Brain creatine functions to attenuate acute stress responses through GABAnergic system in chicks. Neuroscience 2005, 132, 65–71. [Google Scholar] [CrossRef]
- Cunha, M.P.; Pazini, F.L.; Rosa, J.M.; Ramos-Hryb, A.B.; Oliveira, A.; Kaster, M.P.; Rodrigues, A.L. Creatine, similarly to ketamine, affords antidepressant-like effects in the tail suspension test via adenosine A(1) and A2A receptor activation. Purinergic Signal. 2015, 11, 215–227. [Google Scholar] [CrossRef]
- Allen, P.J.; Debold, J.F.; Rios, M.; Kanarek, R.B. Chronic high-dose creatine has opposing effects on depression-related gene expression and behavior in intact and sex hormone-treated gonadectomized male and female rats. Pharmacol. Biochem. Behav. 2015, 130, 22–33. [Google Scholar] [CrossRef] [Green Version]
- Duan, B.; Liu, D.S.; Huang, Y.; Zeng, W.Z.; Wang, X.; Yu, H.; Zhu, M.X.; Chen, Z.Y.; Xu, T.L. PI3-kinase/Akt pathway-regulated membrane insertion of acid-sensing ion channel 1a underlies BDNF-induced pain hypersensitivity. J. Neurosci. 2012, 32, 6351–6363. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n | RMP (mV) | IR (Mohm) | C (pF) | AP Threshold (mV) | EPSP Peak (mV) | |
|---|---|---|---|---|---|---|
| CTRL | 25 | −67.0 ± 2.9 | 140.9 ± 7.8 | 183.4 ± 10.4 | −54.0 ± 1.5 | 9.3 ± 1.3 |
| TREAT | 28 | −63.1 ± 2.9 | 137.9 ± 8.8 | 171.3 ± 9.0 | −55.6 ± 1.3 | 6.4 ± 0.9 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sartini, S.; Lattanzi, D.; Di Palma, M.; Savelli, D.; Eusebi, S.; Sestili, P.; Cuppini, R.; Ambrogini, P. Maternal Creatine Supplementation Positively Affects Male Rat Hippocampal Synaptic Plasticity in Adult Offspring. Nutrients 2019, 11, 2014. https://doi.org/10.3390/nu11092014
Sartini S, Lattanzi D, Di Palma M, Savelli D, Eusebi S, Sestili P, Cuppini R, Ambrogini P. Maternal Creatine Supplementation Positively Affects Male Rat Hippocampal Synaptic Plasticity in Adult Offspring. Nutrients. 2019; 11(9):2014. https://doi.org/10.3390/nu11092014
Chicago/Turabian StyleSartini, Stefano, Davide Lattanzi, Michael Di Palma, David Savelli, Silvia Eusebi, Piero Sestili, Riccardo Cuppini, and Patrizia Ambrogini. 2019. "Maternal Creatine Supplementation Positively Affects Male Rat Hippocampal Synaptic Plasticity in Adult Offspring" Nutrients 11, no. 9: 2014. https://doi.org/10.3390/nu11092014
APA StyleSartini, S., Lattanzi, D., Di Palma, M., Savelli, D., Eusebi, S., Sestili, P., Cuppini, R., & Ambrogini, P. (2019). Maternal Creatine Supplementation Positively Affects Male Rat Hippocampal Synaptic Plasticity in Adult Offspring. Nutrients, 11(9), 2014. https://doi.org/10.3390/nu11092014