Oral Tolerance Induction to Newly Introduced Allergen is Favored by a Transforming Growth Factor-β-Enriched Formula

 ,
,  and
and 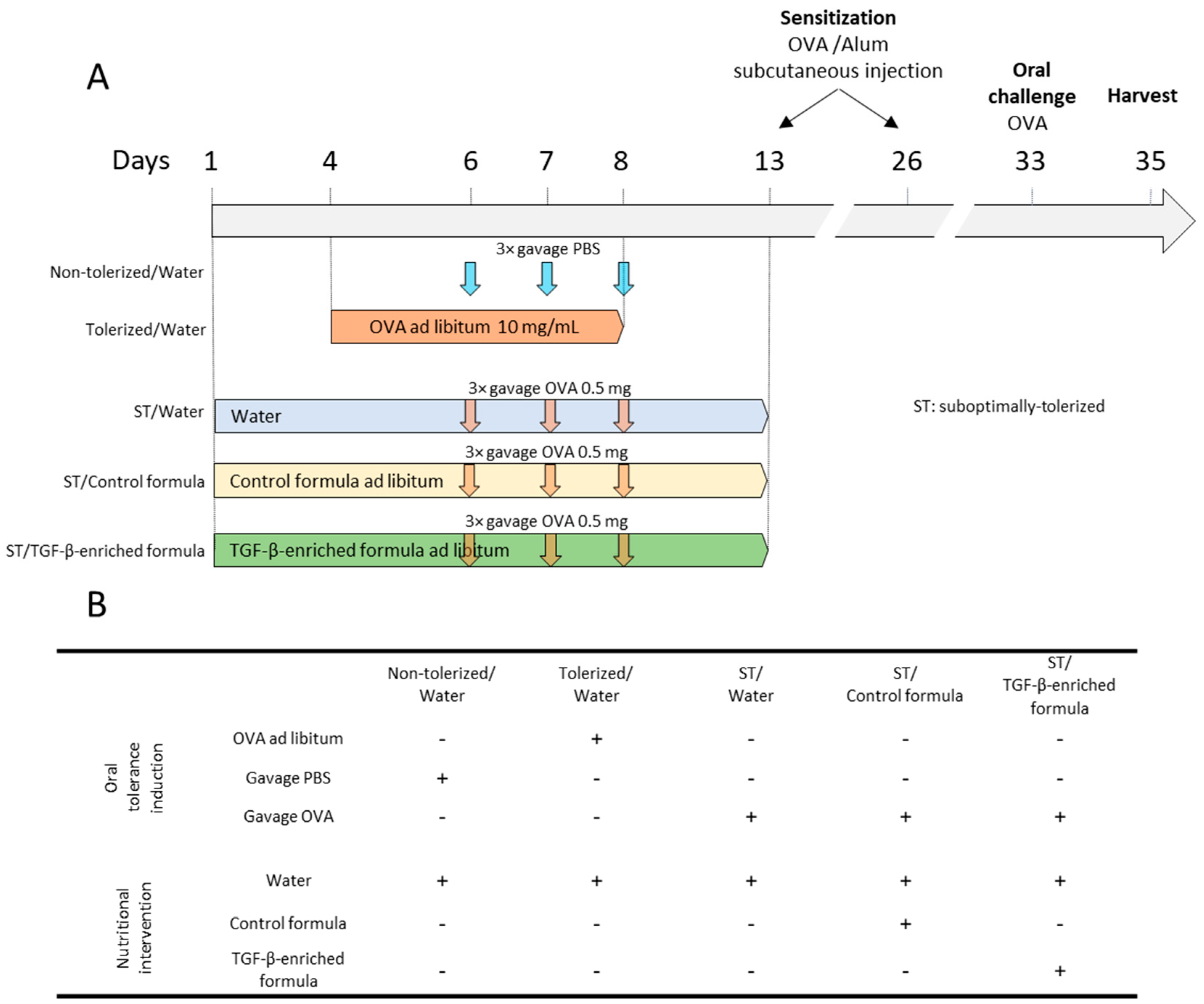{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. TGF-β-Enriched Formula
2.2. Quantification of TGF-β1, TGF-β2, and TGF-β3
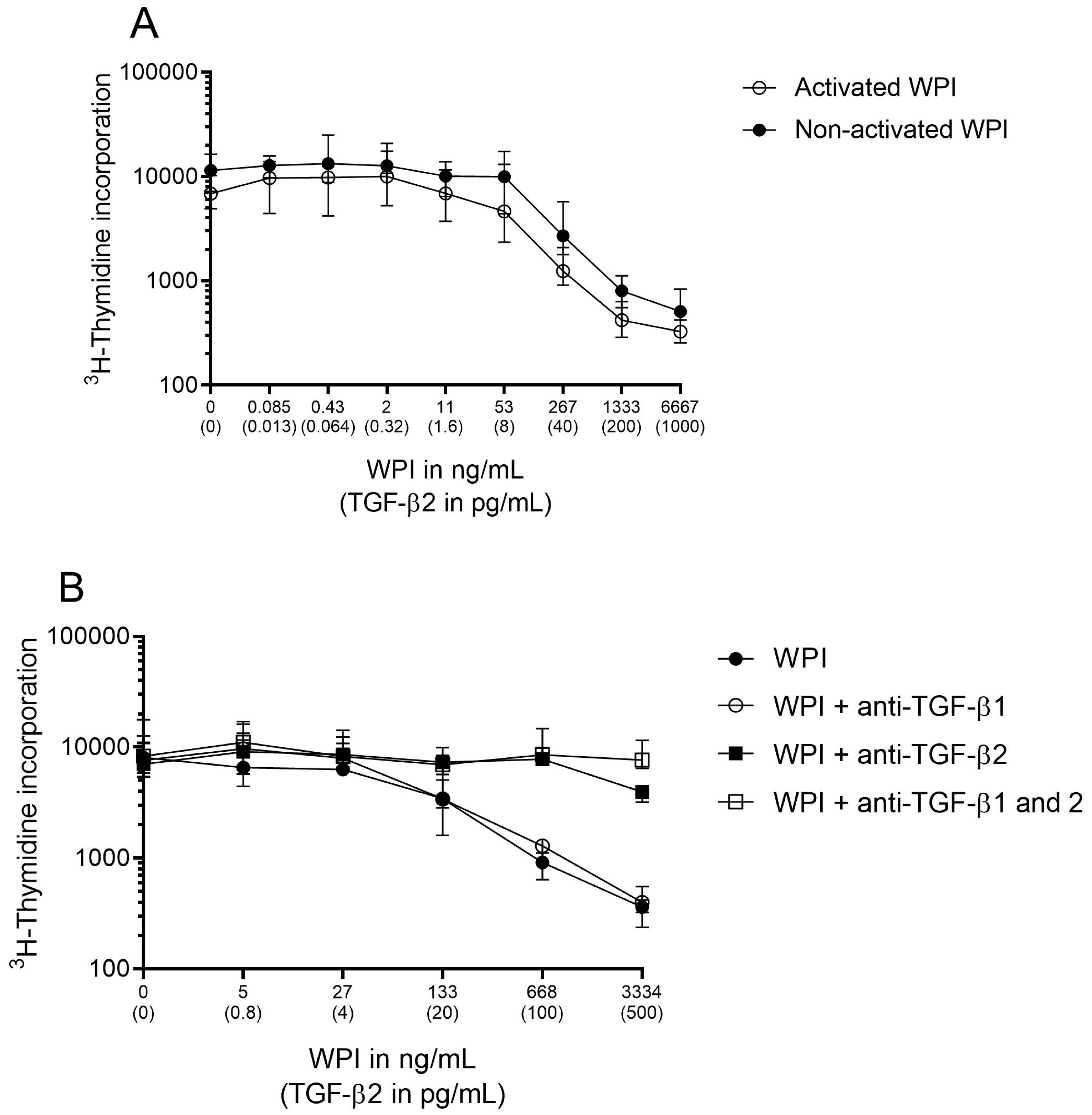
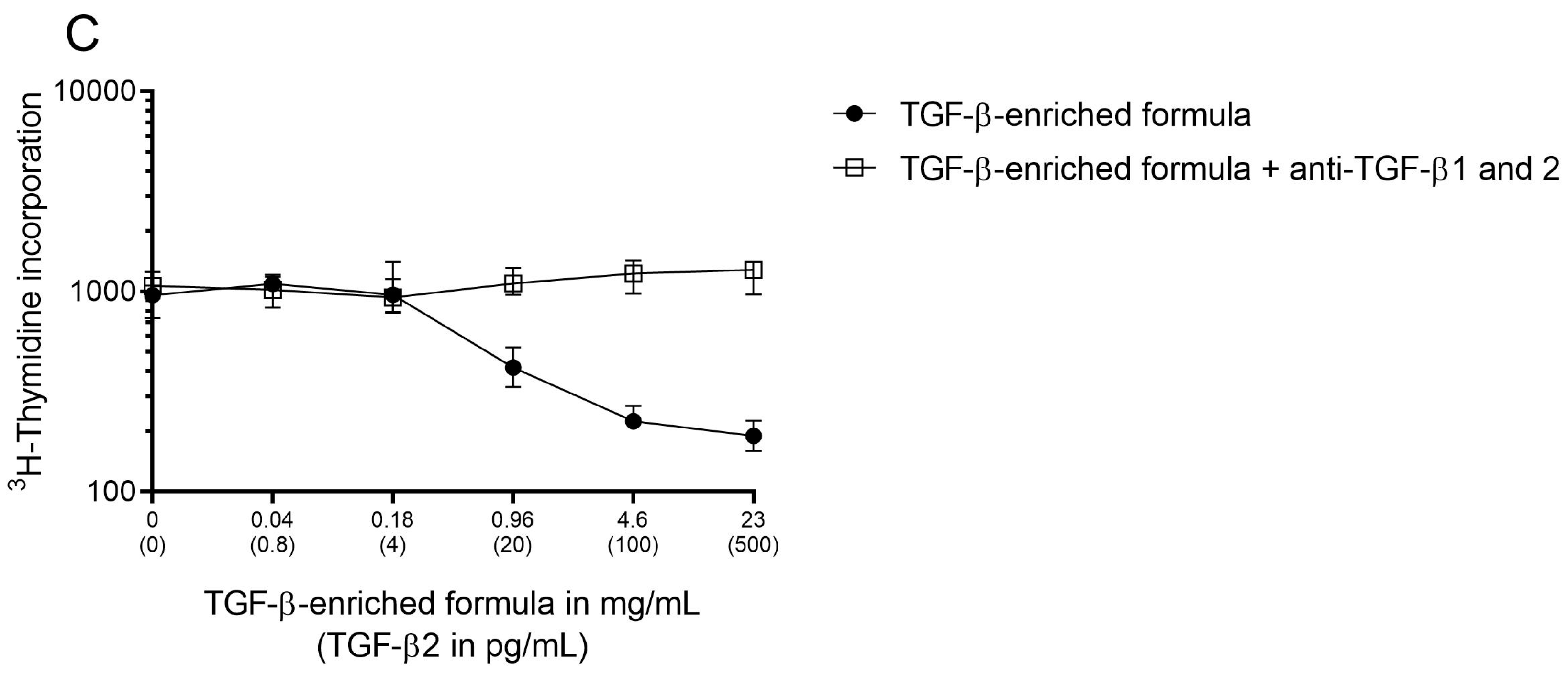
2.3. Assessment of TGF-β Bioactivity
2.4. Induction of Allergic Immune Response to OVA in Mice
2.5. Oral Tolerance and Oral Supplementation
2.6. Quantification of Total IgE, and OVA-Specific IgG1 and MCP-1 in Mouse Plasma
2.7. Isolation and Culture of Axillary and Brachial Lymph Node Cells
2.8. Cytokine Quantification in Culture Supernatants
2.9. Statistical Analyses
3. Results
3.1. Partially Hydrolyzed Whey Formula Supplemented with TGF-β Containing WPI Contained Bioactive TGF-β
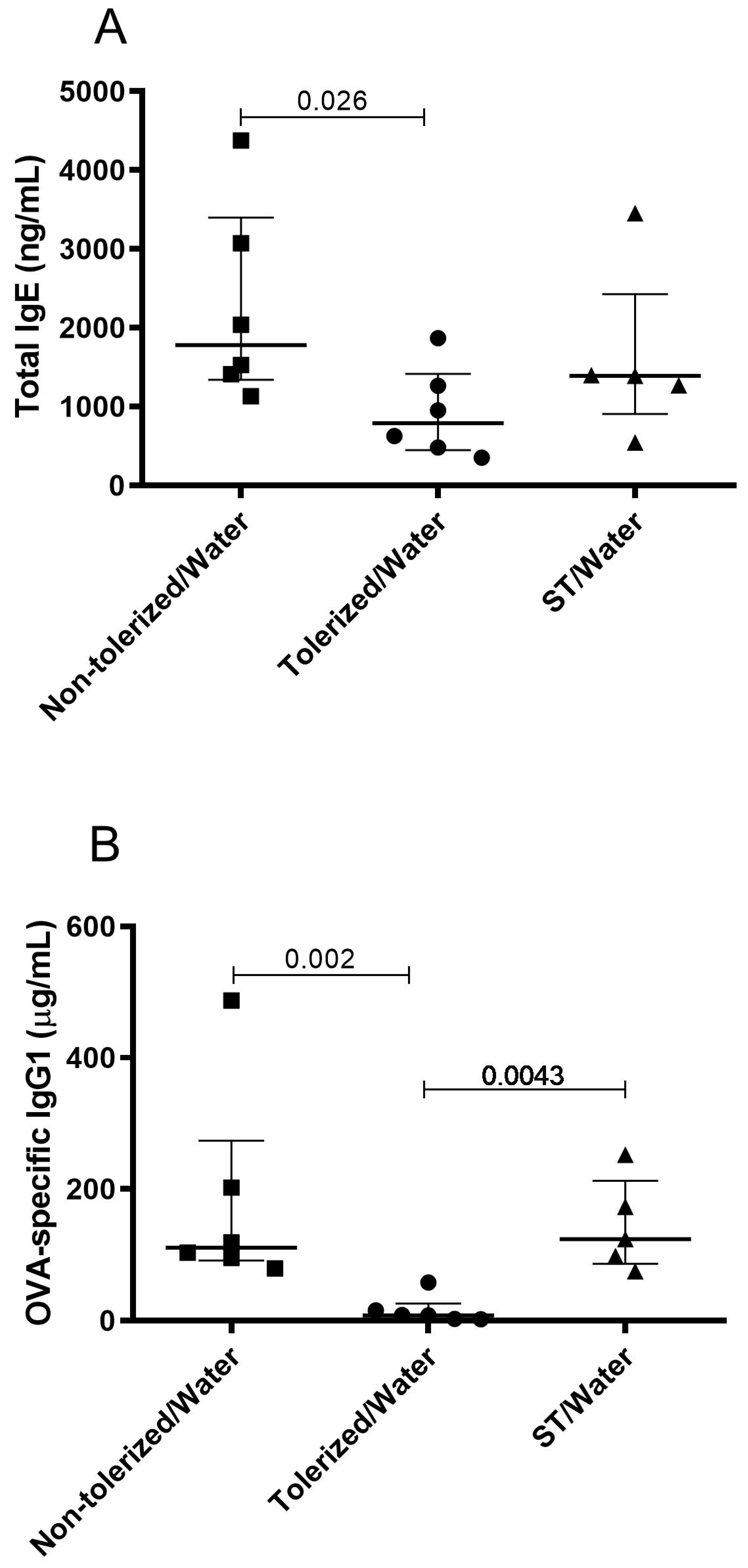
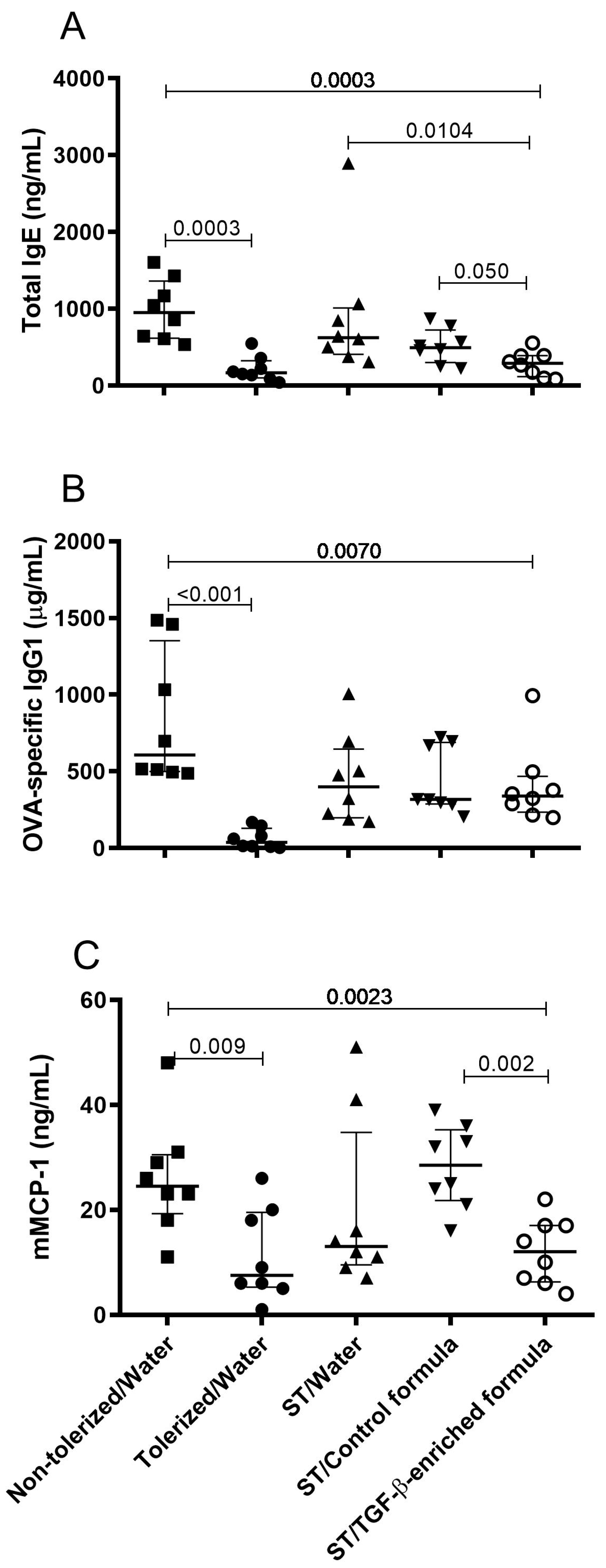
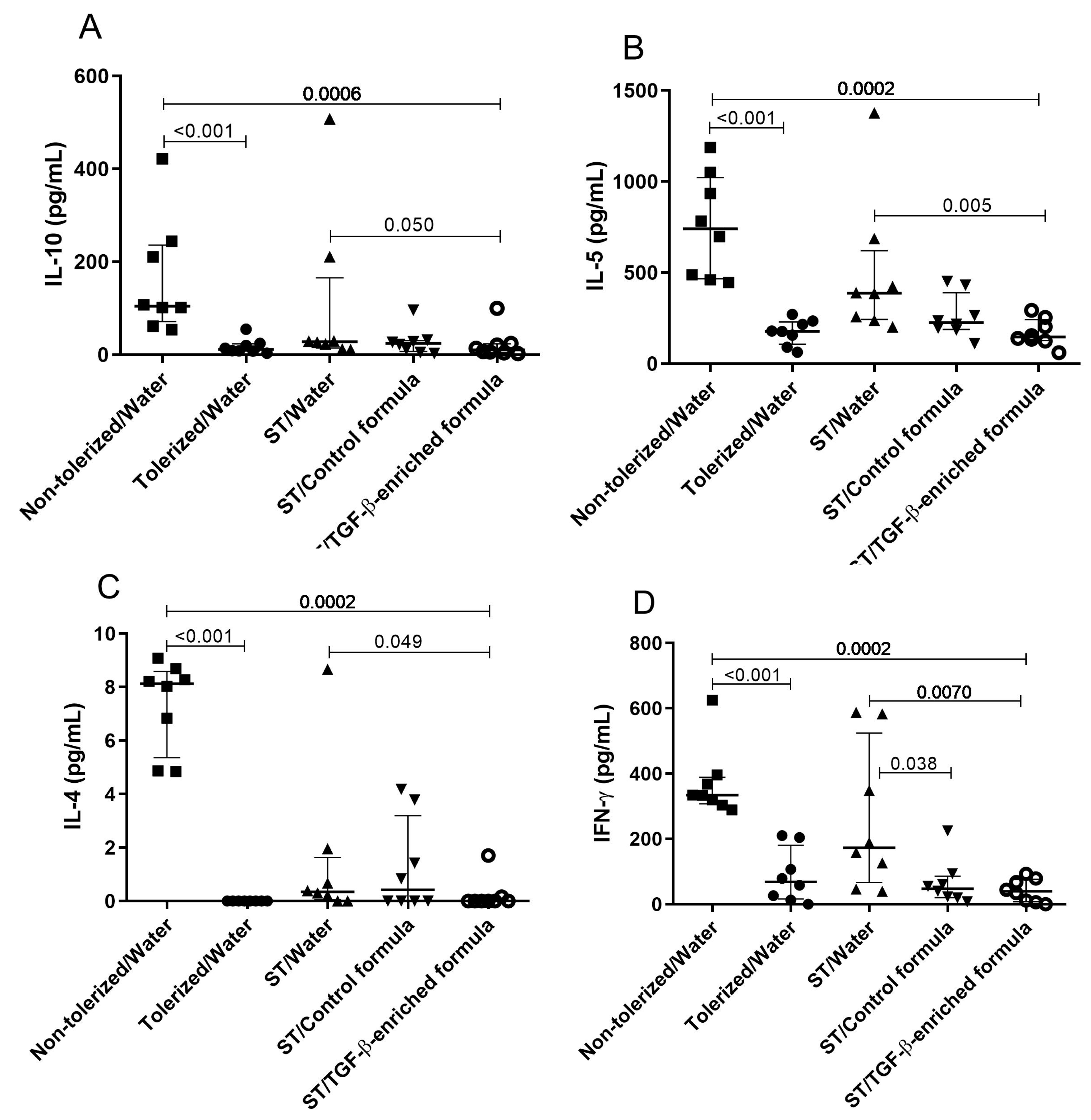
3.2. TGF-β-Enriched Formula Enhanced the Protection Against Sensitization and Response to an Ovalbumin Challenge
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Prescott, S.; Allen, K.J. Food allergy: Riding the second wave of the allergy epidemic. Pediatr. Allergy Immunol. 2011, 22, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Prescott, S.L.; Pawankar, R.; Allen, K.J.; Campbell, D.E.; Dinn, J.K.H.; Fiocchi, A.; Ebisawa, M.; Sampson, U.A.; Beyer, K.; Lee, B.-W. A global survey of changing patterns of food allergy burden in children. World Allergy Organ. J. 2013, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Osborne, N.J.; Koplin, J.J.; Martin, P.E.; Gurrin, L.C.; Lowe, A.J.; Matheson, M.C.; Ponsonby, A.L.; Wake, M.; Tang, M.L.K.; Dharmage, S.C.; et al. Prevalence of challenge-Proven IgE-Mediated food allergy using population-Based sampling and predetermined challenge criteria in infants. J. Allergy Clin. Immunol. 2011, 127, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Pabst, O.; Mowat, A.M. Oral tolerance to food protein. Mucosal Immunol. 2012, 5, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Oddy, W.H. Breastfeeding, childhood asthma, and allergic disease. Ann. Nutr. Metab. 2017, 70, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Jeurink, P.V.; Knipping, K.; Wiens, F.; Barańska, K.; Stahl, B.; Garssen, J.; Krolak-Olejnik, B. Importance of maternal diet in the training of the infant’s immune system during gestation and lactation. Crit. Rev. Food Sci. Nutr. 2018, 2, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sampson, H.U.; O’Mahony, L.; Burks, A.W.; Plaut, M.; Lack, G.; Akdis, C.A. Mechanisms of food allergy. J. Allergy Clin. Immunol. 2018, 141, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Elsen, L.W.J.; Rekima, A.; Verhasselt, V. Early-Life nutrition and gut immune development. Nestle Nutr. Inst. Workshop Ser. 2019, 90, 137–149. [Google Scholar]
- Turfkruyer, M.; Verhasselt, V. Breast milk and its impact on maturation of the neonatal immune system. Curr. Opin. Infect. Dis. 2015, 28, 199–206. [Google Scholar] [CrossRef]
- Munblit, D.; Peroni, D.G.; Boix-Amorós, A.; Hsu, P.T.; Van’t Land, B.; Gay, M.C.L.; Kolotilina, A.; Skevaki, C.; Boyle, R.J.; Collado, M.C. Human milk and allergic diseases: An unsolved puzzle. Nutrients 2017, 9, 894. [Google Scholar] [CrossRef]
- Friedman, N.J.; Zeiger, R.S. The role of breast-Feeding in the development of allergies and asthma. J. Allergy Clin. Immunol. 2005, 115, 1238–1248. [Google Scholar] [CrossRef] [PubMed]
- Oddy, W.H.; Rosales, F. A systematic review of the importance of milk TGF-β on immunological outcomes in the infant and young child. Pediatr. Allergy Immunol. 2010, 21, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Nakao, A. The role and potential use of oral transforming growth factor-β in the prevention of infant allergy. Clin. Exp. Allergy 2010, 40, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.W. Review of the activation of TGF-β in immunity. J. Leukoc. Biol. 2009, 85, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, S.F.; Pouliot, Y.; Maubois, J.L. Growth factors from bovine milk and colostrum: Composition, extraction and biological activities. Lait 2006, 86, 99–125. [Google Scholar] [CrossRef]
- Hawkes, J.S.; Bryan, D.L.; James, M.J.; Gibson, R.A. Cytokines (IL-1beta, IL-6, TNF-alpha, TGF-beta1, and TGF-beta2) and prostaglandin E2 in human milk during the first three months postpartum. Pediatr. Res. 1999, 46, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Sitarik, A.R.; Bobbitt, K.R.; Havstad, S.L.; Fujimura, K.E.; Levin, A.M.; Zoratti, E.M.; Kim, H.; Woodcroft, K.J.; Wegienka, G.; Ownby, D.R.; et al. Breast milk TGFβ is associated with neonatal gut microbial composition. J. Pediatr. Gastroenterol. Nutr. 2017, 65, 60–67. [Google Scholar] [CrossRef]
- Annes, J.P.; Munger, J.S.; Rifkin, D.B. Making sense of latent TGFβ activation. J. Cell Sci. 2003, 116, 217–224. [Google Scholar] [CrossRef]
- Munger, J.S.; Harpel, J.G.; Gleizes, P.E.; Mazzieri, R.; Nunes, I.; Rifkin, D.B. Latent transforming growth factor-β: Structural features and mechanisms of activation. Kidney Int. 1997, 51, 1376–1382. [Google Scholar] [CrossRef]
- Rogers, M.L.; Goddard, C.; Ballard, F.J.; Belford, D.A. Transforming growth factor beta in bovine milk: Concentration, stability and molecular mass forms. J. Endocrinol. 1996, 151, 77–86. [Google Scholar] [CrossRef]
- Nakamura, Y.; Miyata, M.; Ando, T.; Shimokawa, N.; Ohnuma, Y.; Katoh, R.; Ogawa, H.; Okumura, K.; Nakao, A. The latent form of transforming growth factor-β administered orally is activated by gastric acid in mice. J. Nutr. 2009, 139, 1463–1468. [Google Scholar] [CrossRef] [PubMed]
- Faria, A.M.; Weiner, H.L. Oral tolerance and TGF-Beta-Producing cells. Inflamm. Allergy Drug Targets 2006, 5, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Noval Rivas, M.; Chatila, T.A. Regulatory T cells in allergic diseases. J. Allergy Clin. Immunol. 2016, 138, 639–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, Y.; Lee, S.H.; Kim, D.H.; Kang, C.Y. Complementary role of CD4 + CD25 + regulatory T cells and TGF-beta in oral tolerance. J. Leukoc. Biol. 2005, 77, 906–913. [Google Scholar] [CrossRef] [PubMed]
- Szajewska, H.; Horvath, A. A partially hydrolyzed 100% whey formula and the risk of eczema and any allergy: An updated meta-analysis. World Allergy Organ. J. 2017, 10, 27. [Google Scholar] [CrossRef]
- Sauser, J.; Nutten, S.; de Groot, N.; Pecquet, S.; Simon, D.; Simon, H.U.; Spergel, J.M.; Koletzko, S.; Blanchard, C. Partially hydrolyzed whey infant formula: Literature review on effects on growth and the risk of developing atopic dermatitis in infants from the general population. Int. Arch. Allergy Immunol. 2018, 177, 123–124. [Google Scholar] [CrossRef]
- Muraro, A.; Halken, S.; Arshad, S.H.; Beyer, K.; Dubois, A.E.J.; Du Toit, D.; Eigenmann, P.A.; Grimshaw, K.E.; Hoest, A.; Lack, G.; et al. AEECI food allergy and anaphylaxis guidelines. Primary prevention of food allergy. Allergy 2014, 69, 590–601. [Google Scholar] [CrossRef]
- Peroni, D.G.; Piacentini, G.L.; Bodini, A.; Pigozzi, R.; Boner, A.L. Transforming growth factor-β1 is elevated in unpasteurized cow’s milk. Pediatr. Allergy Immunol. 2009, 20, 42–44. [Google Scholar] [CrossRef]
- Forster, R.; Bourtourault, M.; Chung, Y.J.; Silvano, J.; Sire, G.; Spezia, F.; Puel, C.; Descotes, J.; Mikogami, T. Safety evaluation of a whey protein fraction containing a concentrated amount of naturally occurring TGF-β2. Regul. Toxicol. Pharmacol. 2014, 69, 398–407. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (EFSA NDA Panel); Turck, D.; Bresson, J.L.; Burlingame, B.; Dean, T.; Fairweather-Tait, S.; Heinonen, M.; Hirsch-Ernst, K.I.; Mangelsdorf, I.; McArdle, H.J.; et al. Safety of whey basic protein isolates as a novel food pursuant to regulation (EU) 2015/2283. EFSA J. 2018, 16, 5360. [Google Scholar]
- Lyons, R.M.; Keski-Oja, J.; Moses, H.L. Proteolytic activation of latent transforming growth factor-Beta from fibroblast-Conditioned medium. J. Cell Biol. 1988, 106, 1659–1665. [Google Scholar] [CrossRef] [PubMed]
- Castro-Junior, A.B.; Horta, B.C.; Gomes-Santos, A.C.; Cunha, A.P.; Silva Steinberg, R.; Nascimento, D.S.; Faria, A.M.; Vaz, N.M. Oral tolerance correlates with high levels of lymphocyte activity. Cell Immunol. 2012, 280, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Ehirchiou, D.; Xiong, Y.; Xu, G.; Chen, W.; Shi, Y.; Zhang, L. CD11b facilitates the development of peripheral tolerance by suppressing Th17 differentiation. J. Exp. Med. 2007, 204, 1519–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tunis, M.C.; Dawicki, W.; Carson, K.R.; Wang, J.; Marshall, J.S. Mast cells and IgE activation do not alter the development of oral tolerance in a murine model. J. Allergy Clin. Immunol. 2012, 130, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Cascio, J.A.; Haymaker, C.L.; Divekar, R.D.; Zaghouani, S.; Khairallah, M.T.; Wan, X.; Rowland, L.M.; Dhakal, M.; Chen, W.; Zaghouani, H. Antigen-Specific effector CD4 T lymphocytes school lamina propria dendritic cells to transfer innate tolerance. J. Immunol. 2013, 190, 6004–6014. [Google Scholar] [CrossRef] [PubMed]
- Hacini-Rachinel, F.; Vissers, Y.M.; Doucet-Ladevéze, R.; Blanchard, C.; Demont, A.; Perrot, M.; Panchaud, A.; Prioult, G.; Mercenier, A.; Nutten, S. Low-Allergenic hydrolyzed egg induces oral tolerance in mice. Int. Arch. Allergy Immunol. 2014, 164, 64–73. [Google Scholar] [CrossRef]
- Stevens, T.L.; Bossie, A.; Sanders, V.M.; Fernandez-Botran, R.; Coffman, R.L.; Mosmann, T.R.; Vitetta, E.S. Regulation of antibody isotype secretion by subsets of antigen-Specific helper T cells. Nature 1988, 334, 255–258. [Google Scholar] [CrossRef] [PubMed]
- Weiner, H.L.; Friedman, A.; Miller, A.; Khoury, S.J.; Al-Sabbagh, A.; Santos, L.; Sayegh, M.; Nussenblatt, R.B.; Trentham, D.E.; Hafler, D.A. Oral tolerance: Immunologic mechanisms and treatment of animal and human organ-Specific autoimmune diseases by oral administration of autoantigens. Ann. Rev. Immunol. 1994, 12, 809–837. [Google Scholar] [CrossRef]
- Oddy, W.H.; Halonen, M.; Martinez, F.D.; Lohman, I.C.; Stern, D.A.; Kurzius-Spencer, M.; Guerra, S.; Wright, A.L. TGF-β in human milk is associated with wheeze in infancy. J. Allergy Clin. Immunol. 2003, 112, 723–728. [Google Scholar] [CrossRef]
- Laiho, K.; Lampi, A.M.; Hamalainen, M.; Moilanen, E.; Piironen, V.; Arvola, T.; Syrjänen, S.; Isolauri, E. Breast milk fatty acids, eicosanoids and cytokines in mothers with and withour allergic disease. Paediatr. Res. 2003, 53, 642–647. [Google Scholar] [CrossRef]
- Penttila, I.A.; van Spriel, A.B.; Zhang, M.F.; Xian, C.J.; Steeb, C.B.; Cummins, A.G.; Zola, H.; Read, L.C. Transforming growth factor-Beta levels in maternal milk and expression in postnatal rat duodenum and ileum. Pediatr. Res. 1998, 44, 524–531. [Google Scholar] [CrossRef]
- Prokesová, L.; Lodinová-Zádníková, R.; Zizka, J.; Novotná, O.; Petrásková, P.; Sterzl, I. Cytokine levels in healthy and allergic mothers and their children during the first year of life. Pediatr. Allergy Immunol. 2006, 17, 175–183. [Google Scholar] [CrossRef]
- Verhasselt, V.; Milcent, V.; Cazareth, J.; Kanda, A.; Fleury, S.; Dombrowicz, D.; Glaichenhaus, N.; Julia, V. Breast milk-Mediated transfer of an antigen induces tolerance and protection from allergic asthma. Nat. Med. 2008, 14, 170–175. [Google Scholar] [CrossRef]
- Ando, T.; Hatsushika, K.; Wako, M.; Ohba, T.; Koyama, K.; Ohnuma, Y.; Katoh, R.; Ogawa, H.; Okumura, K.; Luo, J.; et al. Orally administered TGF-β is biologically active in the intestinal mucosa and enhances oral tolerance. J. Allergy Clin. Immunol. 2007, 120, 916–923. [Google Scholar] [CrossRef]
- Okamoto, A.; Kawamura, T.; Kanbe, K.; Kanamaru, Y.; Ogawa, H.; Okumara, K.; Nakao, A. Suppression of serum IgE response and systemic anaphylaxis in a food allergy model by orally administered high-Dose TGF-β. Int. Immunol. 2005, 17, 705–712. [Google Scholar] [CrossRef]
- Penttila, I. Effects of transforming growth factor-Beta and formula feeding on systemic immune responses to dietary β-Lactoglobulin in allergy-Prone rats. Pediatr. Res. 2006, 59, 650–655. [Google Scholar] [CrossRef]
- Khaleva, E.; Gridneva, Z.; Geddes, D.T.; Oddy, W.H.; Colicino, S.; Blyuss, O.; Boyle, R.J.; Warner, J.O.; Munblit, D. Transforming growth factor beta in human milk and allergic outcomes in children: A systematic review. Clin. Exp. Allergy 2019. [Google Scholar] [CrossRef]
- Penttila, I.A.; Flesch, I.E.A.; McCue, A.L.; Powell, B.C.; Zhou, F.H.; Read, L.C.; Zola, H. Maternal milk regulation of cell infiltration and interleukin 18 in the intestine of suckling rat pups. Gut 2003, 52, 1579–1586. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.H.; Huang, P.H.; Lee, C.C.; Chen, P.Y.; Chen, H.C. A bovine whey protein extract can induce the generation of regulatory T cells and shows potential to alleviate asthma symptoms in a murine asthma model. Br. J. Nutr. 2013, 109, 1813–1820. [Google Scholar] [CrossRef]
- Rekima, A.; Macchiaverni, P.; Turfkruyer, M.; Holvoet, S.; Dupuis, L.; Baiz, N.; Annesi-Maesano, I.; Mercenier, A.; Nutten, S.; Verhasselt, V. Long-Term reduction in food allergy susceptibility in mice by combining breast-Feeding-Induced tolerance and TGF-β-Enriched formula after weaning. Clin. Exp. Allergy 2017, 47, 565–576. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holvoet, S.; Perrot, M.; de Groot, N.; Prioult, G.; Mikogami, T.; Verhasselt, V.; Nutten, S. Oral Tolerance Induction to Newly Introduced Allergen is Favored by a Transforming Growth Factor-β-Enriched Formula. Nutrients 2019, 11, 2210. https://doi.org/10.3390/nu11092210
Holvoet S, Perrot M, de Groot N, Prioult G, Mikogami T, Verhasselt V, Nutten S. Oral Tolerance Induction to Newly Introduced Allergen is Favored by a Transforming Growth Factor-β-Enriched Formula. Nutrients. 2019; 11(9):2210. https://doi.org/10.3390/nu11092210
Chicago/Turabian StyleHolvoet, Sébastien, Marie Perrot, Nanda de Groot, Guénolée Prioult, Takashi Mikogami, Valérie Verhasselt, and Sophie Nutten. 2019. "Oral Tolerance Induction to Newly Introduced Allergen is Favored by a Transforming Growth Factor-β-Enriched Formula" Nutrients 11, no. 9: 2210. https://doi.org/10.3390/nu11092210
APA StyleHolvoet, S., Perrot, M., de Groot, N., Prioult, G., Mikogami, T., Verhasselt, V., & Nutten, S. (2019). Oral Tolerance Induction to Newly Introduced Allergen is Favored by a Transforming Growth Factor-β-Enriched Formula. Nutrients, 11(9), 2210. https://doi.org/10.3390/nu11092210