Animal Models of Undernutrition and Enteropathy as Tools for Assessment of Nutritional Intervention


Abstract
:1. Introduction
2. Undernutrition Models
2.1. Caloric Restriction
2.2. Protein Undernutrition
2.2.1. Maternal Protein Undernutrition
2.2.2. Protein-Deficient Diet
2.3. Regional Diets
2.4. Zinc-Deficient Diets
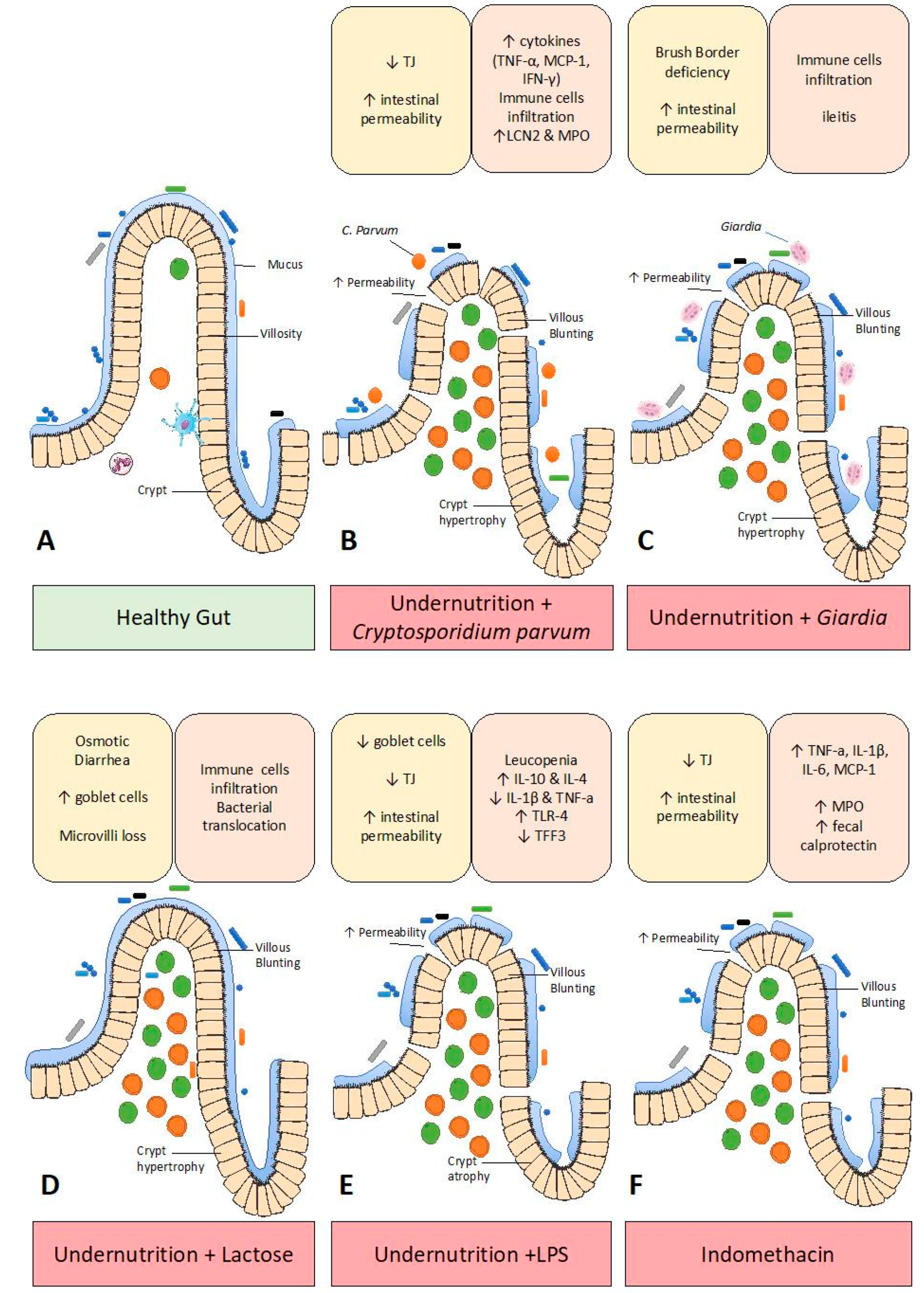
3. Enteropathy Models
3.1. Microbiota Transplantation
3.2. Pathogen-Induced Enteropathy
3.3. Lactose-Induced Enteropathy
3.4. LPS-Induced Enteropathy
3.5. Indomethacin-Induced Enteropathy
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- United Nations Children’s Fund (UNICEF); World Health Organization; International Bank for Reconstruction and Development/The World Bank. Levels and Trends in Child Malnutrition: Key Findings of the 2018 Edition of the Joint Child Malnutrition Estimates; World Health Organization: Geneva, Switzerland, 2018; Licence: CC BY-NC-SA 3.0 IGO. [Google Scholar]
- Black, R.E.; Victora, C.G.; Walker, S.P.; Bhutta, Z.A.; Christian, P.; de Onis, M.; Ezzati, M.; Grantham-McGregor, S.; Katz, J.; Martorell, R.; et al. Maternal and child undernutrition and overweight in low-income and middle-income countries. Lancet 2013, 382, 427–451. [Google Scholar] [CrossRef]
- Victora, C.G.; Adair, L.; Fall, C.; Hallal, P.C.; Martorell, R.; Richter, L.; Sachdev, H.S. Maternal and Child Undernutrition Study Group Maternal and child undernutrition: Consequences for adult health and human capital. Lancet 2008, 371, 340–357. [Google Scholar] [CrossRef]
- Humphrey, J.H. Child undernutrition, tropical enteropathy, toilets, and handwashing. Lancet 2009, 374, 1032–1035. [Google Scholar] [CrossRef]
- Amadi, B.; Besa, E.; Zyambo, K.; Kaonga, P.; Louis-Auguste, J.; Chandwe, K.; Tarr, P.I.; Denno, D.M.; Nataro, J.P.; Faubion, W.; et al. Impaired Barrier Function and Autoantibody Generation in Malnutrition Enteropathy in Zambia. EBioMedicine 2017, 22, 191–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farràs, M.; Chandwe, K.; Mayneris-Perxachs, J.; Amadi, B.; Louis-Auguste, J.; Besa, E.; Zyambo, K.; Guerrant, R.; Kelly, P.; Swann, J.R. Characterizing the metabolic phenotype of intestinal villus blunting in Zambian children with severe acute malnutrition and persistent diarrhea. PLoS ONE 2018, 13, e0192092. [Google Scholar] [CrossRef]
- Attia, S.; Versloot, C.J.; Voskuijl, W.; van Vliet, S.J.; Di Giovanni, V.; Zhang, L.; Richardson, S.; Bourdon, C.; Netea, M.G.; Berkley, J.A.; et al. Mortality in children with complicated severe acute malnutrition is related to intestinal and systemic inflammation: An observational cohort study. Am. J. Clin. Nutr. 2016, 104, 1441–1449. [Google Scholar] [CrossRef] [PubMed]
- Kosek, M.; Guerrant, R.L.; Kang, G.; Bhutta, Z.; Yori, P.P.; Gratz, J.; Gottlieb, M.; Lang, D.; Lee, G.; Haque, R.; et al. Assessment of environmental enteropathy in the MAL-ED cohort study: Theoretical and analytic framework. Clin. Infect. Dis. 2014, 59 (Suppl. 4), S239–S247. [Google Scholar] [CrossRef]
- Demoinet, E.; Roy, R. Analysis of Transgenerational Phenotypes Following Acute Starvation in AMPK-Deficient C. elegans. Methods Mol. Biol. 2018, 1732, 565–579. [Google Scholar]
- Chalvon-Demersay, T.; Even, P.C.; Chaumontet, C.; Piedcoq, J.; Viollet, B.; Gaudichon, C.; Tomé, D.; Foretz, M.; Azzout-Marniche, D. Modifying the Dietary Carbohydrate-to-Protein Ratio Alters the Postprandial Macronutrient Oxidation Pattern in Liver of AMPK-Deficient Mice. J. Nutr. 2017, 147, 1669–1676. [Google Scholar] [CrossRef]
- Ibrahim, M.K.; Zambruni, M.; Melby, C.L.; Melby, P.C. Impact of Childhood Malnutrition on Host Defense and Infection. Clin. Microbiol. Rev. 2017, 30, 919–971. [Google Scholar] [CrossRef] [Green Version]
- Núñez, I.N.; Galdeano, C.M.; Carmuega, E.; Weill, R.; de Moreno de LeBlanc, A.; Perdigón, G. Effect of a probiotic fermented milk on the thymus in Balb/c mice under non-severe protein-energy malnutrition. Br. J. Nutr. 2013, 110, 500–508. [Google Scholar] [CrossRef]
- Pan, F.; Zhang, L.; Li, M.; Hu, Y.; Zeng, B.; Yuan, H.; Zhao, L.; Zhang, C. Predominant gut Lactobacillus murinus strain mediates anti-inflammaging effects in calorie-restricted mice. Microbiome 2018, 6, 54. [Google Scholar] [CrossRef]
- Park, C.Y.; Park, S.; Kim, M.S.; Kim, H.-K.; Han, S.N. Effects of mild calorie restriction on lipid metabolism and inflammation in liver and adipose tissue. Biochem. Biophys. Res. Commun. 2017, 490, 636–642. [Google Scholar] [CrossRef]
- Ma, T.Y.; Hollander, D.; Dadufalza, V.; Krugliak, P. Effect of aging and caloric restriction on intestinal permeability. Exp. Gerontol. 1992, 27, 321–333. [Google Scholar] [CrossRef]
- Mehus, A.A.; Picklo, M.J. Brain and Hepatic Mt mRNA Is Reduced in Response to Mild Energy Restriction and n-3 Polyunsaturated Fatty Acid Deficiency in Juvenile Rats. Nutrients 2017, 9, 1145. [Google Scholar] [CrossRef]
- Jahng, J.W.; Kim, J.G.; Kim, H.J.; Kim, B.-T.; Kang, D.-W.; Lee, J.-H. Chronic food restriction in young rats results in depression- and anxiety-like behaviors with decreased expression of serotonin reuptake transporter. Brain Res. 2007, 1150, 100–107. [Google Scholar] [CrossRef]
- Hallam, M.C.; Reimer, R.A. Postnatal Prebiotic Fiber Intake in Offspring Exposed to Gestational Protein Restriction Has Sex-Specific Effects on Insulin Resistance and Intestinal Permeability in Rats. J. Nutr. 2014, 144, 1556–1563. [Google Scholar] [CrossRef] [Green Version]
- Aubert, P.; Oleynikova, E.; Rizvi, H.; Ndjim, M.; Le Berre-Scoul, C.; Grohard, P.A.; Chevalier, J.; Segain, J.-P.; Le Drean, G.; Neunlist, M.; et al. Maternal protein restriction induces gastrointestinal dysfunction and enteric nervous system remodeling in rat offspring. FASEB J. 2019, 33, 770–781. [Google Scholar] [CrossRef]
- Chen, Y.; Mou, D.; Hu, L.; Zhen, J.; Che, L.; Fang, Z.; Xu, S.; Lin, Y.; Feng, B.; Li, J.; et al. Effects of Maternal Low-Energy Diet during Gestation on Intestinal Morphology, Disaccharidase Activity, and Immune Response to Lipopolysaccharide Challenge in Pig Offspring. Nutrients 2017, 9, 1115. [Google Scholar] [CrossRef]
- D’Inca, R.; Kloareg, M.; Gras-Le Guen, C.; Le Huërou-Luron, I. Intrauterine growth restriction modifies the developmental pattern of intestinal structure, transcriptomic profile, and bacterial colonization in neonatal pigs. J. Nutr. 2010, 140, 925–931. [Google Scholar] [CrossRef]
- Ferenc, K.; Pilżys, T.; Skrzypek, T.; Garbicz, D.; Marcinkowski, M.; Dylewska, M.; Gładysz, P.; Skorobogatov, O.; Gajewski, Z.; Grzesiuk, E.; et al. Structure and Function of Enterocyte in Intrauterine Growth Retarded Pig Neonates. Dis. Markers 2017, 2017, 5238134. [Google Scholar] [CrossRef]
- He, Z.X.; Sun, Z.H.; Yang, W.Z.; Beauchemin, K.A.; Tang, S.X.; Zhou, C.S.; Han, X.F.; Wang, M.; Kang, J.H.; Tan, Z.L. Effects of maternal protein or energy restriction during late gestation on immune status and responses to lipopolysaccharide challenge in postnatal young goats. J. Anim. Sci. 2014, 92, 4856–4864. [Google Scholar] [CrossRef]
- Belmonte, L.; Coëffier, M.; Le Pessot, F.; Miralles-Barrachina, O.; Hiron, M.; Leplingard, A.; Lemeland, J.-F.; Hecketsweiler, B.; Daveau, M.; Ducrotté, P.; et al. Effects of glutamine supplementation on gut barrier, glutathione content and acute phase response in malnourished rats during inflammatory shock. World J. Gastroenterol. 2007, 13, 2833–2840. [Google Scholar] [CrossRef]
- Brown, E.M.; Wlodarska, M.; Willing, B.P.; Vonaesch, P.; Han, J.; Reynolds, L.A.; Arrieta, M.-C.; Uhrig, M.; Scholz, R.; Partida, O.; et al. Diet and specific microbial exposure trigger features of environmental enteropathy in a novel murine model. Nat. Commun. 2015, 6, 7806. [Google Scholar] [CrossRef]
- Eyzaguirre-Velasquez, J.; Olavarria-Ramirez, L.; Gonzalez-Arancibia, C.; Diaz-Merino, C.; Ariz, R.; Lopez, S.; Quiroz, W.; Beltran, C.J.; Bravo, J.A.; Julio-Pieper, M. Protein Malnutrition During Juvenile Age Increases Ileal and Colonic Permeability in the Rat. J. Pediatric Gastroenterol. Nutr. 2017, 64, 707–712. [Google Scholar] [CrossRef]
- Dalvi, P.S.; Yang, S.; Swain, N.; Kim, J.; Saha, S.; Bourdon, C.; Zhang, L.; Chami, R.; Bandsma, R.H.J. Long-term metabolic effects of malnutrition: Liver steatosis and insulin resistance following early-life protein restriction. PLoS ONE 2018, 13, e0199916. [Google Scholar] [CrossRef]
- Fock, R.A.; Vinolo, M.A.R.; Crisma, A.R.; Nakajima, K.; Rogero, M.M.; Borelli, P. Protein-energy malnutrition modifies the production of interleukin-10 in response to lipopolysaccharide (LPS) in a murine model. J. Nutr. Sci. Vitam. 2008, 54, 371–377. [Google Scholar] [CrossRef]
- Mayneris-Perxachs, J.; Bolick, D.T.; Leng, J.; Medlock, G.L.; Kolling, G.L.; Papin, J.A.; Swann, J.R.; Guerrant, R.L. Protein- and zinc-deficient diets modulate the murine microbiome and metabolic phenotype. Am. J. Clin. Nutr. 2016, 104, 1253–1262. [Google Scholar] [CrossRef]
- Santos, A.C.A.; Correia, C.A.; de Oliveira, D.C.; Nogueira-Pedro, A.; Borelli, P.; Fock, R.A. Intravenous Glutamine Administration Modulates TNF-α/IL-10 Ratio and Attenuates NFkB Phosphorylation in a Protein Malnutrition Model. Inflammation 2016, 39, 1883–1891. [Google Scholar] [CrossRef]
- Corware, K.; Yardley, V.; Mack, C.; Schuster, S.; Al-Hassi, H.; Herath, S.; Bergin, P.; Modolell, M.; Munder, M.; Müller, I.; et al. Protein energy malnutrition increases arginase activity in monocytes and macrophages. Nutr. Metab. (Lond.) 2014, 11, 51. [Google Scholar] [CrossRef]
- Ueno, P.M.; Oriá, R.B.; Maier, E.A.; Guedes, M.; Azevedo, O.G.D.; Wu, D.; Willson, T.; Hogan, S.P.; Lima, A.A.M.; Guerrant, R.L.; et al. Alanyl-glutamine promotes intestinal epithelial cell homeostasis in vitro and in a murine model of weanling undernutrition. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 612–622. [Google Scholar] [CrossRef]
- Sampaio, I.C.; Medeiros, P.H.Q.S.; Rodrigues, F.A.P.; Cavalcante, P.A.; Ribeiro, S.A.; Oliveira, J.S.; Prata, M.M.G.; Costa, D.V.S.; Fonseca, S.G.C.; Guedes, M.M.; et al. Impact of acute undernutrition on growth, ileal morphology and nutrient transport in a murine model. Braz. J. Med. Biol. Res. 2016, 49, e5340. [Google Scholar] [CrossRef]
- Charbonneau, M.R.; O’Donnell, D.; Blanton, L.V.; Totten, S.M.; Davis, J.C.C.; Barratt, M.J.; Cheng, J.; Guruge, J.; Talcott, M.; Bain, J.R.; et al. Sialylated Milk Oligosaccharides Promote Microbiota-Dependent Growth in Models of Infant Undernutrition. Cell 2016, 164, 859–871. [Google Scholar] [CrossRef]
- May, T.; Klatt, K.C.; Smith, J.; Castro, E.; Manary, M.; Caudill, M.A.; Jahoor, F.; Fiorotto, M.L. Choline Supplementation Prevents a Hallmark Disturbance of Kwashiorkor in Weanling Mice Fed a Maize Vegetable Diet: Hepatic Steatosis of Undernutrition. Nutrients 2018, 10, 653. [Google Scholar] [CrossRef]
- Lykke, M.; Hother, A.-L.; Hansen, C.F.; Friis, H.; Mølgaard, C.; Michaelsen, K.F.; Briend, A.; Larsen, T.; Sangild, P.T.; Thymann, T. Malnutrition induces gut atrophy and increases hepatic fat infiltration: Studies in a pig model of childhood malnutrition. Am. J. Transl. Res. 2013, 5, 543–554. [Google Scholar]
- Bolick, D.T.; Kolling, G.L.; Moore, J.H.; de Oliveira, L.A.; Tung, K.; Philipson, C.; Viladomiu, M.; Hontecillas, R.; Bassaganya-Riera, J.; Guerrant, R.L. Zinc deficiency alters host response and pathogen virulence in a mouse model of enteroaggregative Escherichia coli-induced diarrhea. Gut Microbes 2014, 5, 618–627. [Google Scholar] [CrossRef]
- Scott, M.E.; Koski, K.G. Zinc deficiency impairs immune responses against parasitic nematode infections at intestinal and systemic sites. J. Nutr. 2000, 130, 1412S–1420S. [Google Scholar] [CrossRef]
- Kau, A.L.; Planer, J.D.; Liu, J.; Rao, S.; Yatsunenko, T.; Trehan, I.; Manary, M.J.; Liu, T.-C.; Stappenbeck, T.S.; Maleta, K.M.; et al. Functional characterization of IgA-targeted bacterial taxa from undernourished Malawian children that produce diet-dependent enteropathy. Sci. Transl. Med. 2015, 7, 276ra24. [Google Scholar] [CrossRef]
- Islam, M.A.; Rahman, M.M.; Mahalanabis, D. Maternal and socioeconomic factors and the risk of severe malnutrition in a child: A case-control study. Eur. J. Clin. Nutr. 1994, 48, 416–424. [Google Scholar]
- Pinheiro, D.F.; Pacheco, P.D.G.; Alvarenga, P.V.; Buratini, J.; Castilho, A.C.S.; Lima, P.F.; Sartori, D.R.S.; Vicentini-Paulino, M.L.M. Maternal protein restriction affects gene expression and enzyme activity of intestinal disaccharidases in adult rat offspring. Braz. J. Med. Biol. Res. 2013, 46, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Batista, T.H.; Veronesi, V.B.; Ribeiro, A.C.A.F.; Giusti-Paiva, A.; Vilela, F.C. Protein malnutrition during pregnancy alters maternal behavior and anxiety-like behavior in offspring. Nutr. Neurosci. 2017, 20, 437–442. [Google Scholar] [CrossRef]
- Crossland, R.F.; Balasa, A.; Ramakrishnan, R.; Mahadevan, S.K.; Fiorotto, M.L.; Van den Veyver, I.B. Chronic Maternal Low-Protein Diet in Mice Affects Anxiety, Night-Time Energy Expenditure and Sleep Patterns, but Not Circadian Rhythm in Male Offspring. PLoS ONE 2017, 12, e0170127. [Google Scholar] [CrossRef]
- Gressens, P.; Muaku, S.M.; Besse, L.; Nsegbe, E.; Gallego, J.; Delpech, B.; Gaultier, C.; Evrard, P.; Ketelslegers, J.M.; Maiter, D. Maternal protein restriction early in rat pregnancy alters brain development in the progeny. Brain Res. Dev. Brain Res. 1997, 103, 21–35. [Google Scholar] [CrossRef]
- Alamy, M.; Bengelloun, W.A. Malnutrition and brain development: An analysis of the effects of inadequate diet during different stages of life in rat. Neurosci. Biobehav. Rev. 2012, 36, 1463–1480. [Google Scholar] [CrossRef]
- Marwarha, G.; Claycombe-Larson, K.; Schommer, J.; Ghribi, O. Maternal low-protein diet decreases brain-derived neurotrophic factor expression in the brains of the neonatal rat offspring. J. Nutr. Biochem. 2017, 45, 54–66. [Google Scholar] [CrossRef]
- Reyes-Castro, L.A.; Padilla-Gómez, E.; Parga-Martínez, N.J.; Castro-Rodríguez, D.C.; Quirarte, G.L.; Díaz-Cintra, S.; Nathanielsz, P.W.; Zambrano, E. Hippocampal mechanisms in impaired spatial learning and memory in male offspring of rats fed a low-protein isocaloric diet in pregnancy and/or lactation. Hippocampus 2018, 28, 18–30. [Google Scholar] [CrossRef]
- Gould, J.M.; Smith, P.J.; Airey, C.J.; Mort, E.J.; Airey, L.E.; Warricker, F.D.M.; Pearson-Farr, J.E.; Weston, E.C.; Gould, P.J.W.; Semmence, O.G.; et al. Mouse maternal protein restriction during preimplantation alone permanently alters brain neuron proportion and adult short-term memory. Proc. Natl. Acad. Sci. USA 2018, 115, E7398–E7407. [Google Scholar] [CrossRef] [Green Version]
- Laeger, T.; Reed, S.D.; Henagan, T.M.; Fernandez, D.H.; Taghavi, M.; Addington, A.; Münzberg, H.; Martin, R.J.; Hutson, S.M.; Morrison, C.D. Leucine acts in the brain to suppress food intake but does not function as a physiological signal of low dietary protein. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R310–R320. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.M.; Laeger, T.; Albarado, D.C.; McDougal, D.H.; Berthoud, H.-R.; Münzberg, H.; Morrison, C.D. Low protein-induced increases in FGF21 drive UCP1-dependent metabolic but not thermoregulatory endpoints. Sci. Rep. 2017, 7, 8209. [Google Scholar] [CrossRef]
- Pezeshki, A.; Zapata, R.C.; Singh, A.; Yee, N.J.; Chelikani, P.K. Low protein diets produce divergent effects on energy balance. Sci. Rep. 2016, 6, 25145. [Google Scholar] [CrossRef]
- Miyazaki, A.; Kandasamy, S.; Michael, H.; Langel, S.N.; Paim, F.C.; Chepngeno, J.; Alhamo, M.A.; Fischer, D.D.; Huang, H.-C.; Srivastava, V.; et al. Protein deficiency reduces efficacy of oral attenuated human rotavirus vaccine in a human infant fecal microbiota transplanted gnotobiotic pig model. Vaccine 2018, 36, 6270–6281. [Google Scholar] [CrossRef]
- Rho, S.; Kim, H.; Shim, S.H.; Lee, S.Y.; Kim, M.J.; Yang, B.-G.; Jang, M.H.; Han, B.W.; Song, M.K.; Czerkinsky, C.; et al. Protein energy malnutrition alters mucosal IgA responses and reduces mucosal vaccine efficacy in mice. Immunol. Lett. 2017, 190, 247–256. [Google Scholar] [CrossRef]
- Fock, R.A.; Rogero, M.M.; Aurélio, M.; Vinolo, R.; Curi, R.; Borges, M.C.; Borelli, P. Effects of Protein-Energy Malnutrition on NF-KappaB Signalling in Murine Peritoneal Macrophages. Inflammation 2010, 33, 101–109. [Google Scholar] [CrossRef]
- Bhutta, Z.A.; Berkley, J.A.; Bandsma, R.H.J.; Kerac, M.; Trehan, I.; Briend, A. Severe childhood malnutrition. Nat. Rev. Dis. Primers 2017, 3, 17067. [Google Scholar] [CrossRef]
- Teodósio, N.R.; Lago, E.S.; Romani, S.A.M.; Guedes, R.C.A. A regional basic diet from Northeast Brazil as a dietary model of experimental malnutrition. Arch. Latinoam. Nutr. 1990, XL, 533–547. [Google Scholar]
- Brigide, P.; Ataide, T.R.; Baptista, A.S.; Canniatti-Brazaca, S.G.; Abdalla, A.L.; Nascimento Filho, V.F.; Piedade, S.M.S.; Sant’ana, A.E.G. Bioavailability of iron in the regional basic diet (RBD) with dietary supplement in Brazil. Biol. Trace Elem. Res. 2011, 140, 53–65. [Google Scholar] [CrossRef]
- De Queiroz, C.A.; Fonseca, S.G.C.; Frota, P.B.; Figueiredo, I.L.; Aragão, K.S.; Magalhães, C.E.C.; de Carvalho, C.B.; Lima, A.Â.M.; Ribeiro, R.A.; Guerrant, R.L.; et al. Zinc treatment ameliorates diarrhea and intestinal inflammation in undernourished rats. BMC Gastroenterol. 2014, 14, 136. [Google Scholar] [CrossRef]
- Bartelt, L.A.; Bolick, D.T.; Kolling, G.L.; Roche, J.K.; Zaenker, E.I.; Lara, A.M.; Noronha, F.J.; Cowardin, C.A.; Moore, J.H.; Turner, J.R.; et al. Cryptosporidium Priming Is More Effective than Vaccine for Protection against Cryptosporidiosis in a Murine Protein Malnutrition Model. PLoS Negl. Trop. Dis. 2016, 10, 1–29. [Google Scholar] [CrossRef]
- Williams, C.D. A nutritional disease of childhood associated with a maize diet. Arch. Dis Child. 1933, 8, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Furuta, C.; Murakami, H. A Novel Concept of Amino Acid Supplementation to Improve the Growth of Young Malnourished Male Rats. Ann. Nutr. Metab. 2018, 72, 231–240. [Google Scholar] [CrossRef]
- Follis, R.H. A kwashiorkor-like syndrome observed in monkeys fed maize. Proc. Soc. Exp. Biol. Med. 1957, 96, 523–528. [Google Scholar] [CrossRef]
- Krebs, N.F.; Miller, L.V.; Hambidge, K.M. Zinc deficiency in infants and children: A review of its complex and synergistic interactions. Paediatr. Int. Child. Health 2014, 34, 279–288. [Google Scholar] [CrossRef]
- Koo, S.I.; Turk, D.E. Effect of zinc deficiency on the ultrastructure of the pancreatic acinar cell and intestinal epithelium in the rat. J. Nutr. 1977, 107, 896–908. [Google Scholar] [CrossRef]
- Wapnir, R.A.; Garcia-Aranda, J.A.; Mevorach, D.E.; Lifshitz, F. Differential absorption of zinc and low-molecular-weight ligands in the rat gut in protein-energy malnutrition. J. Nutr. 1985, 115, 900–908. [Google Scholar] [CrossRef]
- Van Wouwe, J.P.; Veldhuizen, M. Growth characteristics in laboratory animals fed zinc-deficient, copper-deficient, of histidine-supplemented diets. Biol. Trace Elem. Res. 1996, 55, 71–77. [Google Scholar] [CrossRef]
- Zhong, W.; Zhao, Y.; Sun, X.; Song, Z.; McClain, C.J.; Zhou, Z. Dietary zinc deficiency exaggerates ethanol-induced liver injury in mice: Involvement of intrahepatic and extrahepatic factors. PLoS ONE 2013, 8, e76522. [Google Scholar] [CrossRef]
- QS Medeiros, P.H.; Ledwaba, S.E.; Bolick, D.T.; Giallourou, N.; Yum, L.K.; Costa, D.V.S.; Oriá, R.B.; Barry, E.M.; Swann, J.R.; Lima, A.Â.M.; et al. A murine model of diarrhea, growth impairment and metabolic disturbances with Shigella flexneri infection and the role of zinc deficiency. Gut Microbes 2019, 10, 1–16. [Google Scholar] [CrossRef]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, S.; Huq, S.; Yatsunenko, T.; Haque, R.; Mahfuz, M.; Alam, M.A.; Benezra, A.; DeStefano, J.; Meier, M.F.; Muegge, B.D.; et al. Persistent gut microbiota immaturity in malnourished Bangladeshi children. Nature 2014, 510, 417–421. [Google Scholar] [CrossRef]
- Vonaesch, P.; Morien, E.; Andrianonimiadana, L.; Sanke, H.; Mbecko, J.-R.; Huus, K.E.; Naharimanananirina, T.; Gondje, B.P.; Nigatoloum, S.N.; Vondo, S.S.; et al. Stunted childhood growth is associated with decompartmentalization of the gastrointestinal tract and overgrowth of oropharyngeal taxa. Proc. Natl. Acad. Sci. USA 2018, 115, E8489–E8498. [Google Scholar] [CrossRef]
- Bartelt, L.A.; Bolick, D.T.; Mayneris-Perxachs, J.; Kolling, G.L.; Medlock, G.L.; Zaenker, E.I.; Donowitz, J.; Thomas-Beckett, R.V.; Rogala, A.; Carroll, I.M.; et al. Cross-modulation of pathogen-specific pathways enhances malnutrition during enteric co-infection with Giardia lamblia and enteroaggregative Escherichia coli. PLoS Pathog. 2017, 13, e1006471. [Google Scholar] [CrossRef]
- Smith, M.I.; Yatsunenko, T.; Manary, M.J.; Trehan, I.; Mkakosya, R.; Cheng, J.; Kau, A.L.; Rich, S.S.; Concannon, P.; Mychaleckyj, J.C.; et al. Gut microbiomes of Malawian twin pairs discordant for kwashiorkor. Science 2013, 339, 548–554. [Google Scholar] [CrossRef]
- Brown, E.M.; Arrieta, M.-C.; Finlay, B.B. A fresh look at the hygiene hypothesis: How intestinal microbial exposure drives immune effector responses in atopic disease. Semin. Immunol. 2013, 25, 378–387. [Google Scholar] [CrossRef]
- Kau, A.L.; Ahern, P.P.; Griffin, N.W.; Goodman, A.L.; Gordon, J.I. Human nutrition, the gut microbiome and the immune system. Nature 2011, 474, 327–336. [Google Scholar] [CrossRef] [Green Version]
- Tzipori, S.; Ward, H. Cryptosporidiosis: Biology, pathogenesis and disease. Microbes Infect. 2002, 4, 1047–1058. [Google Scholar] [CrossRef]
- Dillingham, R.A.; Lima, A.A.; Guerrant, R.L. Cryptosporidiosis: Epidemiology and impact. Microbes Infect. 2002, 4, 1059–1066. [Google Scholar] [CrossRef]
- Huang, D.B.; Chappell, C.; Okhuysen, P.C. Cryptosporidiosis in children. Semin. Pediatr. Infect. Dis. 2004, 15, 253–259. [Google Scholar] [CrossRef]
- Tanriverdi, S.; Grinberg, A.; Chalmers, R.M.; Hunter, P.R.; Petrovic, Z.; Akiyoshi, D.E.; London, E.; Zhang, L.; Tzipori, S.; Tumwine, J.K.; et al. Inferences about the global population structures of Cryptosporidium parvum and Cryptosporidium hominis. Appl. Environ. Microbiol. 2008, 74, 7227–7234. [Google Scholar] [CrossRef]
- Agnew, D.G.; Lima, A.A.; Newman, R.D.; Wuhib, T.; Moore, R.D.; Guerrant, R.L.; Sears, C.L. Cryptosporidiosis in northeastern Brazilian children: Association with increased diarrhea morbidity. J. Infect. Dis. 1998, 177, 754–760. [Google Scholar] [CrossRef]
- Guerrant, D.I.; Moore, S.R.; Lima, A.A.; Patrick, P.D.; Schorling, J.B.; Guerrant, R.L. Association of early childhood diarrhea and cryptosporidiosis with impaired physical fitness and cognitive function four-seven years later in a poor urban community in northeast Brazil. Am. J. Trop. Med. Hyg. 1999, 61, 707–713. [Google Scholar] [CrossRef]
- Gendrel, D.; Treluyer, J.M.; Richard-Lenoble, D. Parasitic diarrhea in normal and malnourished children. Fundam. Clin. Pharm. 2003, 17, 189–197. [Google Scholar] [CrossRef]
- Katona, P.; Katona-Apte, J. The interaction between nutrition and infection. Clin. Infect. Dis. 2008, 46, 1582–1588. [Google Scholar] [CrossRef]
- Coutinho, B.P.; Oriá, R.B.; Vieira, C.M.G.; Sevilleja, J.E.A.D.; Warren, C.A.; Maciel, J.G.; Thompson, M.R.; Pinkerton, R.C.; Aldo, A.M.; Guerrant, R.L. Cryptospordium Infection Causes Undernutrition and Conversly, Weanling Undernutrition Intensifies Infection. J. Parasitol. 2008, 94, 1225–1232. [Google Scholar] [CrossRef]
- Costa, L.B.; JohnBull, E.A.; Reeves, J.T.; Sevilleja, J.E.; Freire, R.S.; Hoffman, P.S.; Lima, A.A.M.; Oriá, R.B.; Roche, J.K.; Guerrant, R.L.; et al. Cryptosporidium-malnutrition interactions: Mucosal disruption, cytokines, and TLR signaling in a weaned murine model. J. Parasitol. 2011, 97, 1113–1120. [Google Scholar] [CrossRef]
- Bolick, D.T.; Mayneris-Perxachs, J.; Medlock, G.L.; Kolling, G.L.; Papin, J.A.; Swann, J.R.; Guerrant, R.L. Increased Urinary Trimethylamine N-Oxide Following Cryptosporidium Infection and Protein Malnutrition Independent of Microbiome Effects. J. Infect. Dis. 2017, 216, 64–71. [Google Scholar] [CrossRef]
- Tinuade, O.; John, O.; Saheed, O.; Oyeku, O.; Fidelis, N.; Olabisi, D. Parasitic etiology of childhood diarrhea. Indian J. Pediatr 2006, 73, 1081–1084. [Google Scholar] [CrossRef]
- Einarsson, E.; Ma’ayeh, S.; Svärd, S.G. An up-date on Giardia and giardiasis. Curr. Opin. Microbiol. 2016, 34, 47–52. [Google Scholar] [CrossRef]
- Squire, S.A.; Ryan, U. Cryptosporidium and Giardia in Africa: Current and future challenges. Parasites Vectors 2017, 10, 195. [Google Scholar] [CrossRef]
- Caron, Y.; Hong, R.; Gauthier, L.; Laillou, A.; Wieringa, F.T.; Berger, J.; Poirot, E. Stunting, Beyond Acute Diarrhoea: Giardia Duodenalis, in Cambodia. Nutrients 2018, 10, 1420. [Google Scholar] [CrossRef]
- Denno, D.M.; VanBuskirk, K.; Nelson, Z.C.; Musser, C.A.; Hay Burgess, D.C.; Tarr, P.I. Use of the lactulose to mannitol ratio to evaluate childhood environmental enteric dysfunction: A systematic review. Clin. Infect. Dis. 2014, 59 (Suppl. 4), S213–S219. [Google Scholar] [CrossRef]
- Hjelt, K.; Paerregaard, A.; Krasilnikoff, P.A. Giardiasis causing chronic diarrhoea in suburban Copenhagen: Incidence, physical growth, clinical symptoms and small intestinal abnormality. Acta Paediatr. 1992, 81, 881–886. [Google Scholar] [CrossRef]
- Goto, R.; Panter-Brick, C.; Northrop-Clewes, C.A.; Manahdhar, R.; Tuladhar, N.R. Poor intestinal permeability in mildly stunted Nepali children: Associations with weaning practices and Giardia lamblia infection. Br. J. Nutr. 2002, 88, 141–149. [Google Scholar] [CrossRef]
- Upadhyay, P.; Ganguly, N.K.; Mahajan, R.C.; Walia, B.N. Intestinal uptake of nutrients in normal and malnourished animals infected with Giardia lamblia. Digestion 1985, 32, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Buret, A.; Gall, D.G.; Olson, M.E. Effects of murine giardiasis on growth, intestinal morphology, and disaccharidase activity. J. Parasitol. 1990, 76, 403–409. [Google Scholar] [CrossRef]
- Astiazarán-García, H.; Espinosa-Cantellano, M.; Castañón, G.; Chávez-Munguía, B.; Martínez-Palomo, A. Giardia lamblia: Effect of infection with symptomatic and asymptomatic isolates on the growth of gerbils (Meriones unguiculatus). Exp. Parasitol. 2000, 95, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Shukla, G.; Bhatia, R.; Sharma, A. Prebiotic inulin supplementation modulates the immune response and restores gut morphology in Giardia duodenalis-infected malnourished mice. Parasitol. Res. 2016, 115, 4189–4198. [Google Scholar] [CrossRef]
- Shukla, G.; Sidhu, R.K. Lactobacillus casei as a probiotic in malnourished Giardia lamblia-infected mice: A biochemical and histopathological study. Can. J. Microbiol. 2011, 57, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Bartelt, L.A.; Roche, J.; Kolling, G.; Bolick, D.; Noronha, F.; Naylor, C.; Hoffman, P.; Warren, C.; Singer, S.; Guerrant, R.; et al. lamblia impairs growth in a murine malnutrition model. J. Clin. Investig. 2013, 123, 2672–2684. [Google Scholar] [CrossRef]
- Platts-Mills, J.A.; Babji, S.; Bodhidatta, L.; Gratz, J.; Haque, R.; Havt, A.; McCormick, B.J.; McGrath, M.; Olortegui, M.P.; Samie, A.; et al. Pathogen-specific burdens of community diarrhoea in developing countries: A multisite birth cohort study (MAL-ED). Lancet Glob. Health 2015, 3, e564–e575. [Google Scholar] [CrossRef]
- Muhsen, K.; Levine, M.M. A systematic review and meta-analysis of the association between Giardia lamblia and endemic pediatric diarrhea in developing countries. Clin. Infect. Dis. 2012, 55 (Suppl. 4), S271–S293. [Google Scholar] [CrossRef]
- Kosek, M.N. MAL-ED Network Investigators Causal Pathways from Enteropathogens to Environmental Enteropathy: Findings from the MAL-ED Birth Cohort Study. EBioMedicine 2017, 18, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Rogawski, E.T.; Liu, J.; Platts-Mills, J.A.; Kabir, F.; Lertsethtakarn, P.; Siguas, M.; Khan, S.S.; Praharaj, I.; Murei, A.; Nshama, R.; et al. Use of quantitative molecular diagnostic methods to investigate the effect of enteropathogen infections on linear growth in children in low-resource settings: Longitudinal analysis of results from the MAL-ED cohort study. Lancet Glob. Health 2018, 6, e1319–e1328. [Google Scholar] [CrossRef]
- Diarrhoeal Disease. Available online: https://www.who.int/news-room/fact-sheets/detail/diarrhoeal-disease (accessed on 20 April 2019).
- Grenov, B.; Briend, A.; Sangild, P.T.; Thymann, T.; Rytter, M.H.; Hother, A.-L.; Mølgaard, C.; Michaelsen, K.F. Undernourished Children and Milk Lactose. Food Nutr. Bull. 2016, 37, 85–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, S.K.; Sachan, A.; Udupa, V.; Seshadri, S.J.; Jayakumar, K. Histological changes in intestine in semichronic diarrhoea induced by lactose enriched diet in rats: Effect of Diarex-Vet. Indian J. Exp. Biol. 2003, 41, 211–215. [Google Scholar] [PubMed]
- Galvez, J.; Zarzuelo, A.; Crespo, M.E.; Lorente, M.D.; Ocete, M.A.; Jiménez, J. Antidiarrhoeic Activity of Euphorbia hirta Extract and Isolation of an Active Flavonoid Constituent. Planta Med. 1993, 59, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Leforestier, G.; Blais, A.; Blachier, F.; Marsset-Baglieri, A.; Davila-Gay, A.-M.; Perrin, E.; Tomé, D. Effects of galacto-oligosaccharide ingestion on the mucosa-associated mucins and sucrase activity in the small intestine of mice. Eur. J. Nutr. 2009, 48, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, C.; Trent, M.S. Biosynthesis and export of bacterial lipopolysaccharides. Annu. Rev. Biochem. 2014, 83, 99–128. [Google Scholar] [CrossRef]
- Park, B.S.; Lee, J.-O. Recognition of lipopolysaccharide pattern by TLR4 complexes. Exp. Mol. Med. 2013, 45, e66. [Google Scholar] [CrossRef]
- Bandsma, R.H.J.; Ackerley, C.; Koulajian, K.; Zhang, L.; Zutphen, T.V.; Dijk, T.H.V.; Xiao, C.; Giacca, A.; Lewis, G.F. A low-protein diet combined with low-dose endotoxin leads to changes in glucose homeostasis in weanling rats. Am. J. Physiol. Endocrinol. Metab. 2015, 309, 466–473. [Google Scholar] [CrossRef]
- Fock, R.A.; Vinolo, M.A.R.; Blatt, S.L.; Borelli, P. Impairment of the hematological response and interleukin-1?? production in protein-energy malnourished mice after endotoxemia with lipopolysaccharide. Braz. J. Med. Biol. Res. 2012, 45, 1163–1171. [Google Scholar] [CrossRef]
- Iwasa, T.; Matsuzaki, T.; Tungalagsuvd, A.; Munkhzaya, M.; Kuwahara, A.; Yasui, T.; Irahara, M. Prenatal undernutrition increases the febrile response to lipopolysaccharides in adulthood in male rats. Int. J. Dev. Neurosci. 2015, 44, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Lyoumi, S.; Tamion, F.; Petit, J.; Dechelotte, P.; Dauguet, C.; Scotte, M.; Hiron, M.; Leplingard, A.; Salier, J.P.; Daveau, M.; et al. Induction and modulation of acute-phase response by protein malnutrition in rats: Comparative effect of systemic and localized inflammation on interleukin-6 and acute-phase protein synthesis. J. Nutr. 1998, 128, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Wang, Q.; Wang, M.; Jia, J.; Wu, R. Gardenia Decoction Prevent Intestinal Mucosal Injury by Inhibiting Pro-inflammatory Cytokines and NF-κB Signaling. Front. Pharm. 2019, 10, 180. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.-J.; Qian, J.-X.; Wei, Y.; Guo, Q.; Jin, J.; Sun, X.; Liu, S.-L.; Xu, C.-F.; Zhang, G.-X. Tanshinone IIA Sodium Sulfonate Attenuates LPS-Induced Intestinal Injury in Mice. Gastroenterol. Res. Pract. 2018, 2018, 9867150. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Xu, Y.; Yang, D.; Yu, N.; Bai, Z.; Bian, L. Effect of Polysaccharides from Acanthopanax senticosus on Intestinal Mucosal Barrier of Escherichia coli Lipopolysaccharide Challenged Mice. Asian-Australas. J. Anim. Sci. 2016, 29, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Nozu, T.; Miyagishi, S.; Kumei, S.; Nozu, R.; Takakusaki, K.; Okumura, T. Lovastatin inhibits visceral allodynia and increased colonic permeability induced by lipopolysaccharide or repeated water avoidance stress in rats. Eur. J. Pharm. 2018, 818, 228–234. [Google Scholar] [CrossRef] [Green Version]
- Bein, A.; Zilbershtein, A.; Golosovsky, M.; Davidov, D.; Schwartz, B. LPS Induces Hyper-Permeability of Intestinal Epithelial Cells. J. Cell. Physiol. 2017, 232, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Al-sadi, R.; Said, H.M.; Ma, T.Y. Lipopolysaccharide Causes an Increase in Intestinal Tight Junction Permeability in Vitro and in Vivo by Inducing Enterocyte Membrane Expression and Localization of. Am. J. Pathol. 2013, 182, 375–387. [Google Scholar] [CrossRef]
- Guo, S.; Nighot, M.; Al-Sadi, R.; Alhmoud, T.; Nighot, P.; Ma, T.Y. Lipopolysaccharide Regulation of Intestinal Tight Junction Permeability Is Mediated by TLR4 Signal Transduction Pathway Activation of FAK and MyD88. J. Immunol. 2015, 195, 4999–5010. [Google Scholar] [CrossRef]
- Yue, C.; Wang, W.; Tian, W.-L.; Huang, Q.; Zhao, R.-S.; Zhao, Y.-Z.; Li, Q.-R.; Li, J.-S. Lipopolysaccharide-induced failure of the gut barrier is site-specific and inhibitable by growth hormone. Inflamm. Res. 2013, 62, 407–415. [Google Scholar] [CrossRef]
- Chen, S.; Bu, D.; Ma, Y.; Zhu, J.; Sun, L.; Zuo, S.; Ma, J.; Li, T.; Chen, Z.; Zheng, Y.; et al. GYY4137 ameliorates intestinal barrier injury in a mouse model of endotoxemia. Biochem. Pharm. 2016, 118, 59–67. [Google Scholar] [CrossRef]
- Holloway, P.M.; Gillespie, S.; Becker, F.; Vital, S.A.; Nguyen, V.; Alexander, J.S.; Evans, P.C.; Gavins, F.N.E. Sulforaphane induces neurovascular protection against a systemic inflammatory challenge via both Nrf2-dependent and independent pathways. Vasc. Pharm. 2016, 85, 29–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, K.C.; van Mil, S.; Murray, E.; Mallet, J.-F.; Matar, C.; Ismail, N. Age and sex differences in immune response following LPS treatment in mice. Brain Behav. Immun. 2016, 58, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Thun, M.J.; Henley, S.J.; Patrono, C. Nonsteroidal Anti-inflammatory Drugs as Anticancer Agents: Mechanistic, Pharmacologic, and Clinical Issues. J. Natl. Cancer Inst. 2002, 94, 252–266. [Google Scholar] [CrossRef] [Green Version]
- Nash, D.B. The Use of Medicines in the United States: A Detailed Review. Am. Health Drug Benefits 2012, 5, 423. [Google Scholar] [PubMed]
- Syer, S.D.; Wallace, J.L. Environmental and NSAID-enteropathy: Dysbiosis as a common factor. Curr. Gastroenterol. Rep. 2014, 16, 377. [Google Scholar] [CrossRef]
- Graham, D.Y.; Opekun, A.R.; Willingham, F.F.; Qureshi, W.A. Visible small-intestinal mucosal injury in chronic NSAID users. Clin. Gastroenterol. Hepatol. 2005, 3, 55–59. [Google Scholar] [CrossRef]
- Sugimori, S.; Watanabe, T.; Tabuchi, M.; Kameda, N.; Machida, H.; Okazaki, H.; Tanigawa, T.; Yamagami, H.; Shiba, M.; Watanabe, K.; et al. Evaluation of small bowel injury in patients with rheumatoid arthritis by capsule endoscopy: Effects of anti-rheumatoid arthritis drugs. Digestion 2008, 78, 208–213. [Google Scholar] [CrossRef]
- Tanaka, A.; Hase, S.; Miyazawa, T.; Ohno, R.; Takeuchi, K. Role of cyclooxygenase (COX)-1 and COX-2 inhibition in nonsteroidal anti-inflammatory drug-induced intestinal damage in rats: Relation to various pathogenic events. J. Pharm. Exp. 2002, 303, 1248–1254. [Google Scholar] [CrossRef]
- Fortun, P.J.; Hawkey, C.J. Nonsteroidal antiinflammatory drugs and the small intestine. Curr. Opin. Gastroenterol. 2005, 21, 169–175. [Google Scholar] [CrossRef]
- Bjarnason, I.; Hayllar, J.; MacPherson, A.J.; Russell, A.S. Side effects of nonsteroidal anti-inflammatory drugs on the small and large intestine in humans. Gastroenterology 1993, 104, 1832–1847. [Google Scholar] [CrossRef]
- Horibe, S.; Tanahashi, T.; Kawauchi, S.; Mizuno, S.; Rikitake, Y. Preventative effects of sodium alginate on indomethacin-induced small-intestinal injury in mice. Int. J. Med. Sci. 2016, 13, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, K.; Kurihara, C.; Furuhashi, H.; Takajo, T.; Maruta, K.; Yasutake, Y.; Sato, H.; Narimatsu, K.; Okada, Y.; Higashiyama, M.; et al. Psychological stress exacerbates NSAID-induced small bowel injury by inducing changes in intestinal microbiota and permeability via glucocorticoid receptor signaling. J. Gastroenterol. 2017, 52, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.-M.; Park, J.-M.; Kang, J.X.; Cha, J.-Y.; Lee, H.-J.; Jeong, M.; Go, E.-J.; Hahm, K.B. Mitigation of indomethacin-induced gastrointestinal damages in fat-1 transgenic mice via gate-keeper action of $ømega$-3-polyunsaturated fatty acids. Sci. Rep. 2016, 6, 33992. [Google Scholar] [CrossRef] [PubMed]
- Nagai, K.; Ueno, Y.; Tanaka, S.; Hayashi, R.; Shinagawa, K.; Chayama, K. Polysaccharides derived from Ganoderma lucidum fungus mycelia ameliorate indomethacin-induced small intestinal injury via induction of GM-CSF from macrophages. Cell. Immunol. 2017, 320, 20–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwak, D.S.; Lee, O.Y.; Lee, K.N.; Jun, D.W.; Lee, H.L.; Yoon, B.C.; Choi, H.S. The Effect of DA-6034 on Intestinal Permeability in an Indomethacin- Induced Small Intestinal Injury Model. Gut Liver 2016, 10, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Jacob, M.; Foster, R.; Sigthorsson, G.; Simpson, R.; Bjarnason, I. Role of bile in pathogenesis of indomethacin-induced enteropathy. Arch. Toxicol. 2007, 81, 291–298. [Google Scholar] [CrossRef]
- Whitfield-Cargile, C.M.; Cohen, N.D.; Chapkin, R.S.; Weeks, B.R.; Davidson, L.A.; Goldsby, J.S.; Hunt, C.L.; Steinmeyer, S.H.; Menon, R.; Suchodolski, J.S.; et al. The microbiota-derived metabolite indole decreases mucosal inflammation and injury in a murine model of NSAID enteropathy. Gut Microbes 2016, 7, 246–261. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, V.; Guimbaud, R.; Tulliez, M.; Mauprivez, C.; Sogni, P.; Couturier, D.; Giroud, J.P.; Chaussade, S.; Chauvelot-Moachon, L. Increase in tumor necrosis factor-alpha production linked to the toxicity of indomethacin for the rat small intestine. Br. J. Pharm. 1998, 124, 1385–1394. [Google Scholar] [CrossRef]
- Saud, B.; Nandi, J.; Ong, G.; Finocchiaro, S.; Levine, R.A. Inhibition of TNF-alpha improves indomethacin-induced enteropathy in rats by modulating iNOS expression. Dig. Dis. Sci. 2005, 50, 1677–1683. [Google Scholar] [CrossRef]
- Watanabe, T.; Higuchi, K.; Kobata, A.; Nishio, H.; Tanigawa, T.; Shiba, M.; Tominaga, K.; Fujiwara, Y.; Oshitani, N.; Asahara, T.; et al. Non-steroidal anti-inflammatory drug-induced small intestinal damage is Toll-like receptor 4 dependent. Gut 2008, 57, 181–187. [Google Scholar] [CrossRef]
- Konaka, A.; Kato, S.; Tanaka, A.; Kunikata, T.; Korolkiewicz, R.; Takeuchi, K. Roles of enterobacteria, nitric oxide and neutrophil in pathogenesis of indomethacin-induced small intestinal lesions in rats. Pharm. Res. 1999, 40, 517–524. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Preclinical Model | Growth | Gut Hyperpermeability | Gut Inflammation | Ref. |
|---|---|---|---|---|
| Caloric Restriction (CR) | ||||
| −15% CR | Weight loss | n/a | Lower MCP-1 mRNA (liver/adipose tissue) | [14] |
| −25% CR | Weight loss | n/a | Thymus atrophy | [12] |
| −30% CR | Weight loss | No gut hyperpermeability | Decreased systemic TNFα levels | [13] |
| Intrauterine Undernutrition | ||||
| 8% of protein in gestational rats/20% in offspring | Low birth weight | Lower colonic ZO-1 mRNA expression | n/a | [18] |
| No hyperpermeability to LPS | [18] | |||
| Ex vivo colonic hyperpermeability to FSA | [19] | |||
| 20% in gestating sows/low birth weight | Low birth weight | Lower villus length ileum/duodenum | [20,21,22] | |
| 7.3% in gestating sows/20% for piglets | Low birth weight | Lower villus:crypt ratio in piglets | Lower IL-6 and TNF-α mRNA in offspring (ileum) | [20] |
| Maternal protein restriction or energy restriction (60% vs. CT) | Low birth weight | n/a | Lower C3, C4, IgG, and IgM concentration in plasma offspring | [23] |
| Decreased jejunal IL-2 and IL-6 mRNA expression in offspring | ||||
| Protein Energy Undernutrition | ||||
| 0% of protein | Weight loss | Lower jejunal villus length | No difference in plasmatic α-1-Acid Glycoprotein | [24] |
| 7% of protein | Weight loss | No villus atrophy/ | Decreased MCP-1 macrophages release in vitro | [25] |
| In vivo hyperpermeability | ||||
| Lower jejunal ZO-1 and higher claudin-2 mRNA | ||||
| 4% protein | Weight loss | No hyperpermeability to large molecules | n/a | [26] |
| Hyperpermeability to small molecules | n/a | |||
| Lower colonic and ileal occludin | n/a | |||
| n/a | Higher TNF-α, MCP-1 and IL1-β production (liver) | [27] | ||
| Leucopenia with higher systemic IL-10 production | [28] | |||
| Lower CD-4 and TLR-4/MD-2 (macrophages) | ||||
| Lower IL-6, TNF-a and IL1-B production by cultured cell from bone marrow, spleen and peritoneum after in vitro LPS treatment | ||||
| 2% protein | Weight loss | n/a | Higher MPO and LCN-2 production | [29] |
| Decreased leucocyte, peripheral lymphocyte, monocyte and polynuclear cells levels | [30] | |||
| Lower TLR-4 expression | [28] | |||
| Lower mRNA expression of TNF-α by macrophage in vitro | ||||
| Lower NF-κB activation in vitro | ||||
| 0.7% of protein | Weight loss | n/a | Increased monocytes and macrophages number in bone marrow and blood | [31] |
| Higher monocyte arginase expression | ||||
| RBD | Weight loss | Jejunal hyperpermeability | n/a | [32] |
| Decreased jejunal claudin-3 tight junction protein expression | ||||
| Jejunal villous, crypt atrophy | ||||
| Lower ileal basal short circuit current | [33] | |||
| Higher ileal claudin-2 and occludin mRNA expression | ||||
| Ileal crypt atrophy No villous length difference | ||||
| M8 | Weight loss | n/a | n/a | [34] |
| MAIZE | Weight loss | Small mucosal atrophy/shorter villi and crypt | n/a | [35,36] |
| Zinc deficiency | ||||
| Zinc deficiency | Weight loss | No impact on villi/crypt ratio | No inflammation | [37] |
| Decreased villi/crypt (EAEC -infected mice) | Decreased TNF-a, IL1-B and IL-6 | |||
| No ileal hyperpermeability | Decreased ileal neutrophil infiltration | [38] | ||
| No higher plasmatic endotoxin | Altered immune response to parasitic nematodes | |||
| Lower production of IL-4, Decreased level of IgE, IgG1 | ||||
| Lower eosinophils and impaired of antigen-presenting cells function | ||||
| Microbiota Transfer | ||||
| Cocktail of bacteria | Weight loss | Increased global intestinal permeability | Increased IL-6, MCP-1 (macrophages) | [25] |
| Villus and crypt atrophy | ||||
| IgA+ bacteria consortium + M8 in gnotobiotic mice | Higher weight loss | Villus and crypt atrophy | Bacterial translocation | [39] |
| Mucosal immune activation with neutrophils infiltration into lamina propria |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salameh, E.; Morel, F.B.; Zeilani, M.; Déchelotte, P.; Marion-Letellier, R. Animal Models of Undernutrition and Enteropathy as Tools for Assessment of Nutritional Intervention. Nutrients 2019, 11, 2233. https://doi.org/10.3390/nu11092233
Salameh E, Morel FB, Zeilani M, Déchelotte P, Marion-Letellier R. Animal Models of Undernutrition and Enteropathy as Tools for Assessment of Nutritional Intervention. Nutrients. 2019; 11(9):2233. https://doi.org/10.3390/nu11092233
Chicago/Turabian StyleSalameh, Emmeline, Fanny B. Morel, Mamane Zeilani, Pierre Déchelotte, and Rachel Marion-Letellier. 2019. "Animal Models of Undernutrition and Enteropathy as Tools for Assessment of Nutritional Intervention" Nutrients 11, no. 9: 2233. https://doi.org/10.3390/nu11092233
APA StyleSalameh, E., Morel, F. B., Zeilani, M., Déchelotte, P., & Marion-Letellier, R. (2019). Animal Models of Undernutrition and Enteropathy as Tools for Assessment of Nutritional Intervention. Nutrients, 11(9), 2233. https://doi.org/10.3390/nu11092233