Chemical Profile, Antioxidative, and Gut Microbiota Modulatory Properties of Ganpu Tea: A Derivative of Pu-erh Tea


Abstract
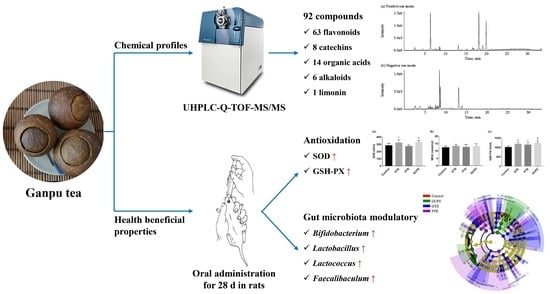
:
1. Introduction
2. Materials and Methods
2.1. Herbal Material and Chemicals
2.2. Sample Preparation
2.3. UHPLC-Q-TOF-MS/MS Analysis
2.4. Animals and Experimental Design
2.5. Assay of Antioxidant Enzyme Activity in Serum
2.6. Gut Microbiota Analysis Using 16S rRNA Gene Sequencing
2.7. Statistical Analysis
3. Results and Discussion
3.1. Identification of Chemical Compounds in GTE by UHPLC-Q-TOF-MS/MS
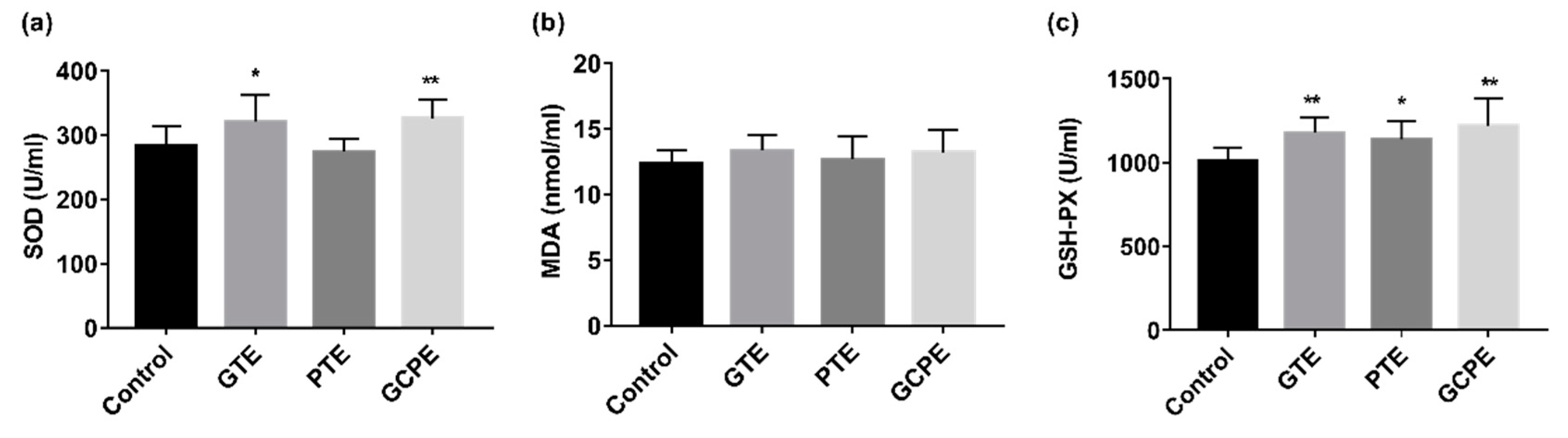
3.2. Antioxidant Activities
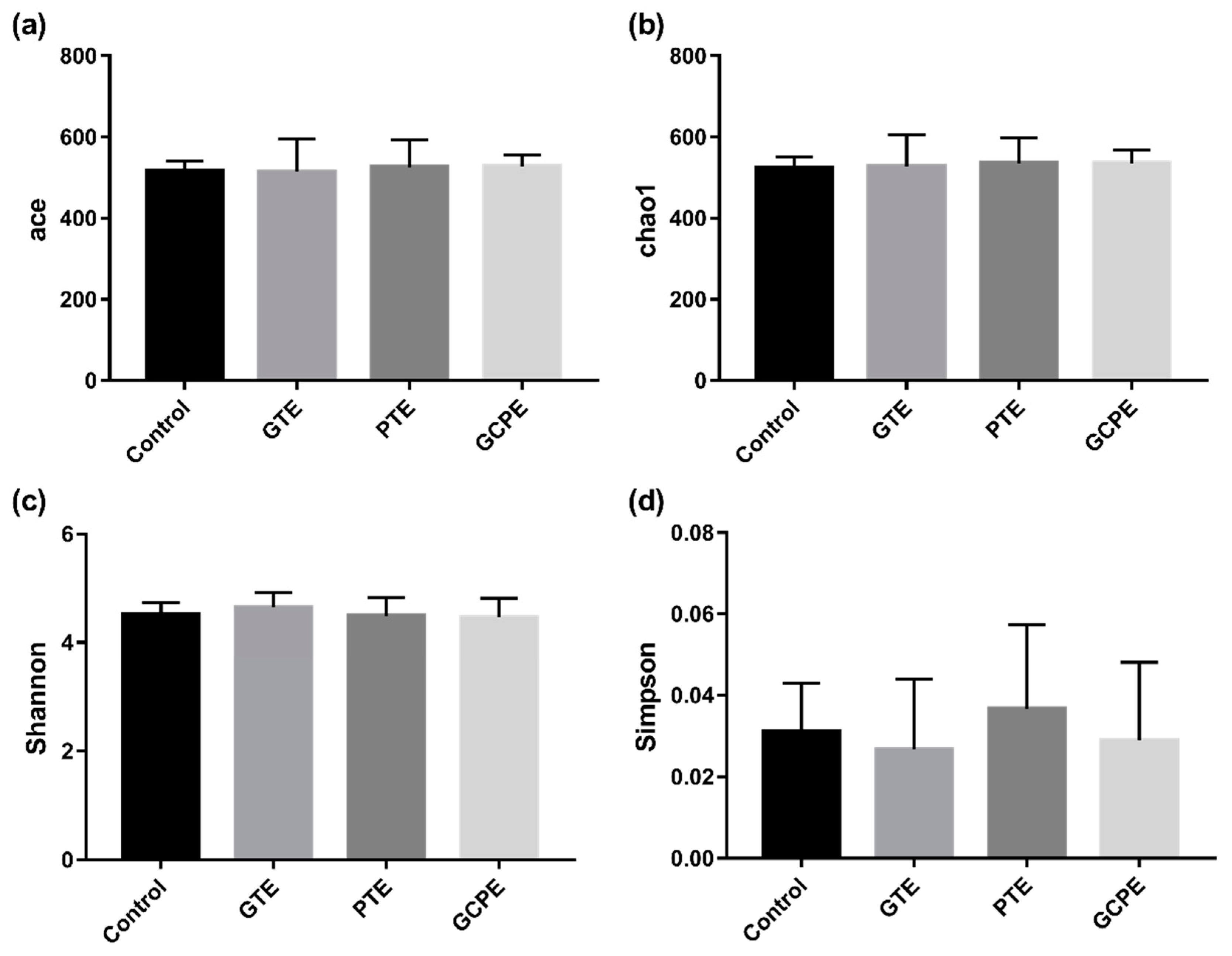
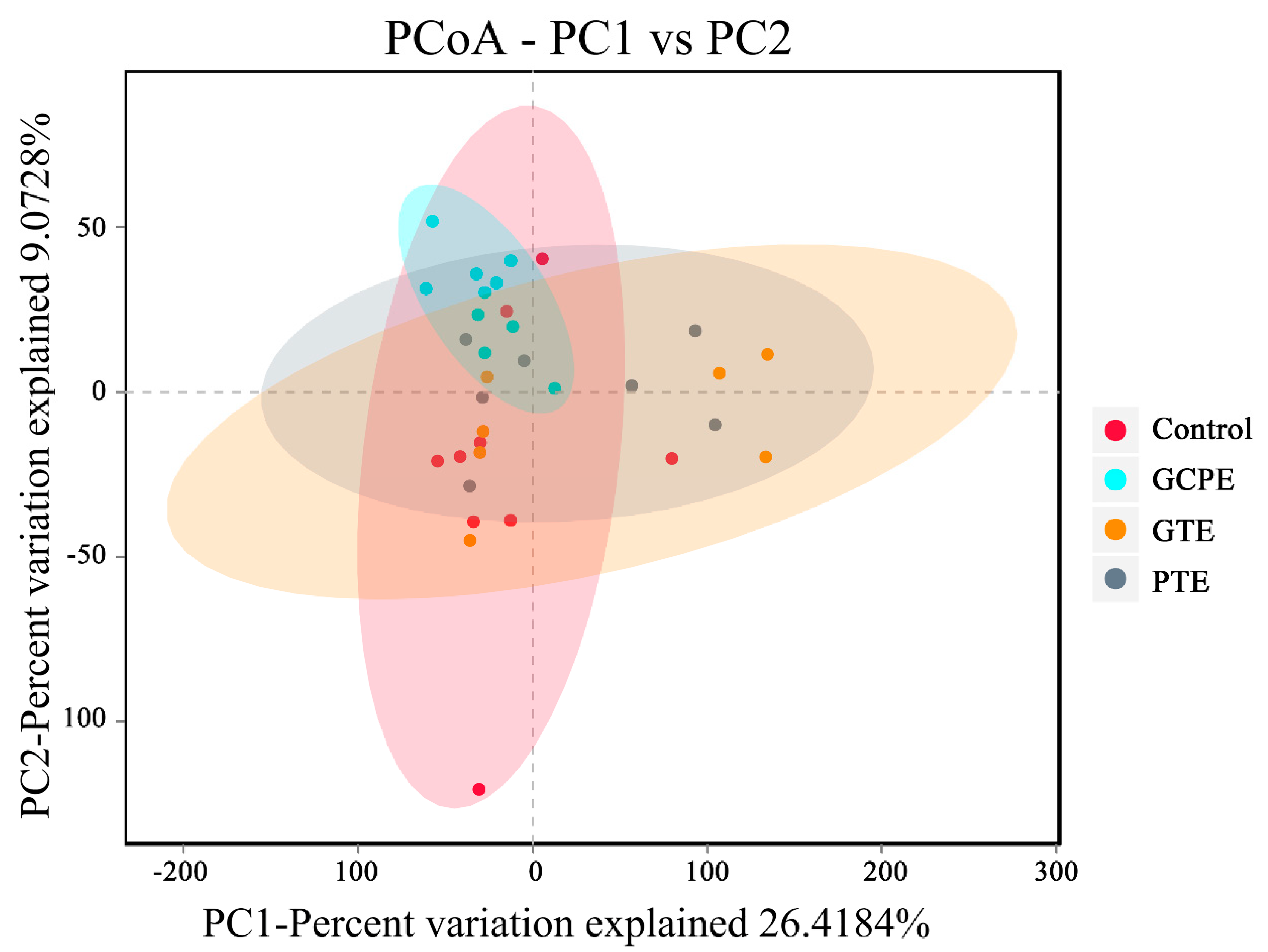
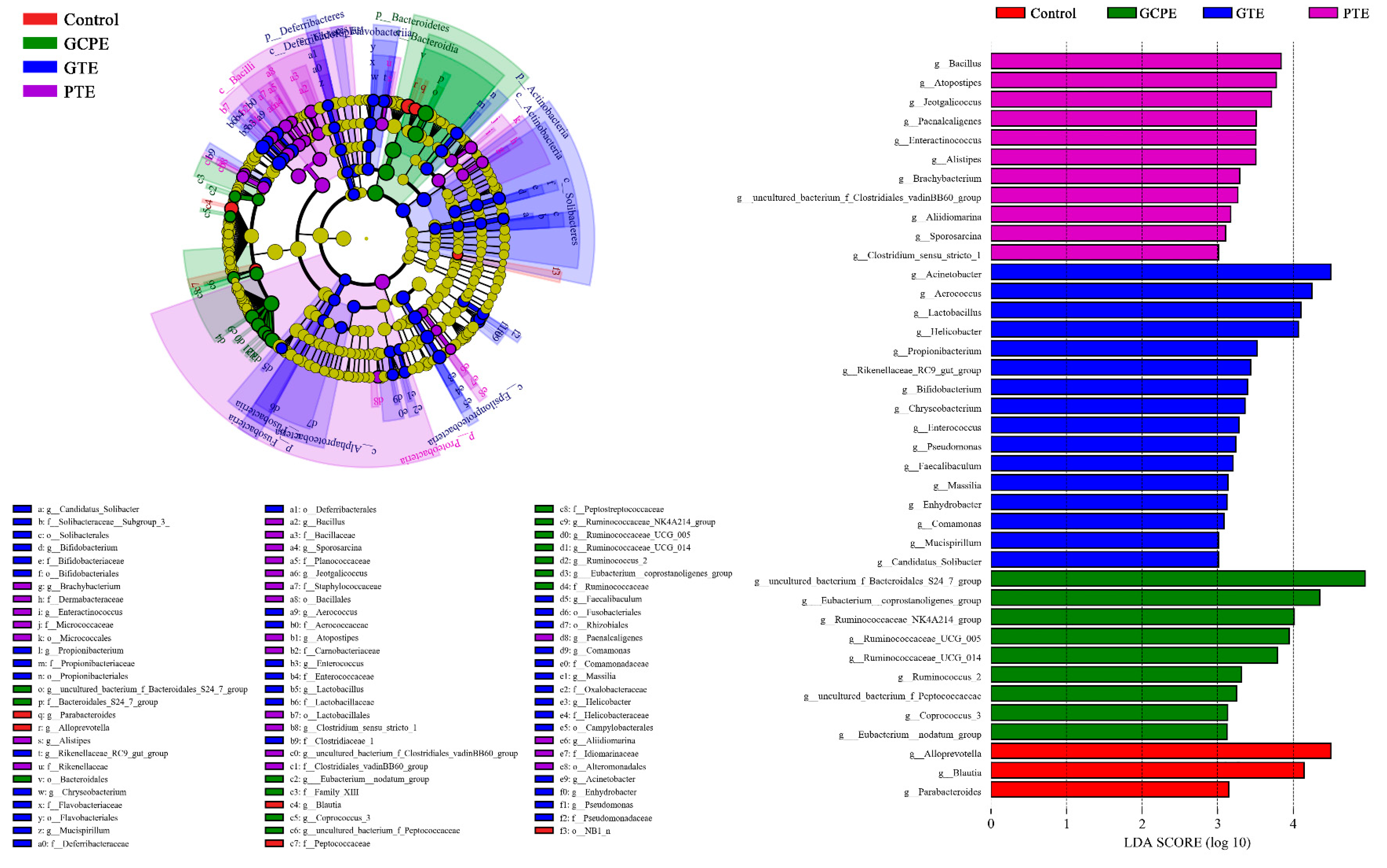
3.3. Effects on Fecal Bacteria Composition
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lv, H.-P.; Zhang, Y.-J.; Lin, Z.; Liang, Y.-R. Processing and chemical constituents of Pu-erh tea: A review. Food Res. Int. 2013, 53, 608–618. [Google Scholar] [CrossRef]
- Duh, P.D.; Yen, G.C.; Yen, W.J.; Wang, B.S.; Chang, L.W. Effects of pu-erh tea on oxidative damage and nitric oxide scavenging. J. Agric. Food Chem. 2004, 52, 8169–8176. [Google Scholar] [CrossRef] [PubMed]
- Kuo, K.L.; Weng, M.S.; Chiang, C.T.; Tsai, Y.J.; Lin-Shiau, S.Y.; Lin, J.K. Comparative studies on the hypolipidemic and growth suppressive effects of oolong, black, pu-erh, and green tea leaves in rats. J. Agric. Food Chem. 2005, 53, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zeng, X.; Peng, W.; Wu, Z.; Su, W. Study on the discrimination between Citri Reticulatae Pericarpium varieties based on HS-SPME-GC-MS combined with multivariate statistical analyses. Molecules 2018, 23, 1235. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Sun, S.; Guo, Y.; Liu, Y.; Yang, D.; Li, G.; Lu, S. Citri Reticulatae Pericarpium (Chenpi): Botany, ethnopharmacology, phytochemistry, and pharmacology of a frequently used traditional Chinese medicine. J. Ethnopharmacol. 2018, 220, 265–282. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zeng, X.; Peng, W.; Wu, Z.; Su, W. Characterisation and classification of Citri Reticulatae Pericarpium varieties based on UHPLC-Q-TOF-MS/MS combined with multivariate statistical analyses. Phytochem. Anal. 2019, 30, 278–291. [Google Scholar] [CrossRef]
- Fu, M.; Xu, Y.; Chen, Y.; Wu, J.; Yu, Y.; Zou, B.; An, K.; Xiao, G. Evaluation of bioactive flavonoids and antioxidant activity in Pericarpium Citri Reticulatae (Citrus reticulata ‘Chachi’) during storage. Food Chem. 2017, 230, 649–656. [Google Scholar] [CrossRef]
- Luo, Y.; Zeng, W.; Huang, K.E.; Li, D.X.; Chen, W.; Yu, X.Q.; Ke, X.H. Discrimination of Citrus reticulata Blanco and Citrus reticulata ‘Chachi’ as well as the Citrus reticulata ‘Chachi’ within different storage years using ultra high performance liquid chromatography quadrupole/time-of-flight mass spectrometry based metabolomics approach. J. Pharm. Biomed. Anal. 2019, 171, 218–231. [Google Scholar]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef] [Green Version]
- Feng, Q.; Chen, W.D.; Wang, Y.D. Gut microbiota: An integral moderator in health and disease. Front. Microbiol. 2018, 9, 151. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambeth, S.M.; Carson, T.; Lowe, J.; Ramaraj, T.; Leff, J.W.; Luo, L.; Bell, C.J.; Shah, V.O. Composition, diversity and abundance of gut microbiome in prediabetes and type 2 diabetes. J. Diabetes Obes. 2015, 2, 1–7. [Google Scholar] [PubMed] [Green Version]
- Guinane, C.M.; Cotter, P.D. Role of the gut microbiota in health and chronic gastrointestinal disease: Understanding a hidden metabolic organ. Ther. Adv. Gastroenterol. 2013, 6, 295–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, W.; Ao, H.; Peng, C.; Yan, D. Gut microbiota, a new frontier to understand traditional Chinese medicines. Pharmacol. Res. 2019, 142, 176–191. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Remesy, C.; Jimenez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Tanja, M.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar]
- Bolger, A.M.; Marc, L.; Bjoern, U. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Han, T.X.; Xu, X.Y.; Zhang, M.J.; Peng, X.; Du, L.L. Global fitness profiling of fission yeast deletion strains by barcode sequencing. Genome Biol. 2010, 11, R60. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.; Su, W.; Bai, Y.; Chen, T.; Yan, Z.; Wang, J.; Su, M.; Zheng, Y.; Peng, W.; Yao, H. Urinary metabolite profiling of flavonoids in Chinese volunteers after consumption of orange juice by UFLC-Q-TOF-MS/MS. J. Chromatogr. B 2017, 1061–1062, 79–88. [Google Scholar] [CrossRef]
- Zeng, X.; Su, W.; Zheng, Y.; Liu, H.; Li, P.; Zhang, W.; Liang, Y.; Bai, Y.; Peng, W.; Yao, H. UFLC-Q-TOF-MS/MS-based screening and identification of flavonoids and derived metabolites in human urine after oral administration of Exocarpium Citri Grandis extract. Molecules 2018, 23, 895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Chen, Z.; Guo, H.; He, D.; Zhao, H.; Wang, Z.; Zhang, W.; Liao, L.; Zhang, C.; Ni, L. The modulatory effect of infusions of green tea, oolong tea, and black tea on gut microbiota in high-fat-induced obese mice. Food Funct. 2016, 7, 4869–4879. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Akhtar, N.; Sambhav, K.; Shete, G.; Bansal, A.K.; Sharma, S.S. Emerging potential of citrus flavanones as an antioxidant in diabetes and its complications. Curr. Top. Med. Chem. 2015, 15, 187–195. [Google Scholar] [CrossRef]
- Scalbert, A.; Manach, C.; Morand, C.; Remesy, C.; Jimenez, L. Dietary polyphenols and the prevention of diseases. Crit. Rev. Food Sci. Nutr. 2005, 45, 287–306. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, J.; Gu, S.; Liu, Z.; Zhang, Y.; Zhang, X. Simultaneous determination of flavonoids in different parts of Citrus reticulata ‘Chachi’ fruit by high performance liquid chromatography-photodiode array detection. Molecules 2010, 15, 5378–5388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Xiao, W.; Ma, Y.; Sun, T.; Yuan, W.; Tang, N.; Zhang, D.; Wang, Y.; Li, Y.; Zhou, H.; et al. Structure and dynamics of the bacterial communities in fermentation of the traditional Chinese post-fermented pu-erh tea revealed by 16S rRNA gene clone library. World J. Microbiol. Biotechnol. 2013, 29, 1877–1884. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zeng, G.; Cai, X.; Deng, S.; Luo, H.; Sun, G. Brachybacterium zhongshanense sp. nov., a cellulose-decomposing bacterium from sediment along the Qijiang River, Zhongshan City, China. Int. J. Syst. Evol. Microbiol. 2007, 57, 2519–2524. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.A.; Grant, L.J.; Gidley, M.J.; Mikkelsen, D. Gut fermentation of dietary fibres: Physico-chemistry of plant cell walls and implications for health. Int. J. Mol. Sci. 2017, 18, 2203. [Google Scholar] [CrossRef] [Green Version]
- Kemperman, R.A.; Gross, G.; Mondot, S.; Possemiers, S.; Marzorati, M.; Van de Wiele, T.; Doré, J.; Vaughan, E.E. Impact of polyphenols from black tea and red wine/grape juice on a gut model microbiome. Food Res. Int. 2013, 53, 659–669. [Google Scholar] [CrossRef]
- Mosele, J.I.; Gosalbes, M.J.; Macia, A.; Rubio, L.; Vazquez-Castellanos, J.F.; Jimenez Hernandez, N.; Moya, A.; Latorre, A.; Motilva, M.J. Effect of daily intake of pomegranate juice on fecal microbiota and feces metabolites from healthy volunteers. Mol. Nutr. Food Res. 2015, 59, 1942–1953. [Google Scholar] [CrossRef] [PubMed]
- Etxeberria, U.; Hijona, E.; Aguirre, L.; Milagro, F.I.; Bujanda, L.; Rimando, A.M.; Martinez, J.A.; Portillo, M.P. Pterostilbene-induced changes in gut microbiota composition in relation to obesity. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Backhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-C.; Walker, W.A. Probiotics and prebiotics: Role in clinical disease states. Adv. Pediatr. 2005, 52, 77–113. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Shan, X.; Cai, C.; Hao, J.; Li, G.; Yu, G. Dietary fucoidan modulates the gut microbiota in mice by increasing the abundance of Lactobacillus and Ruminococcaceae. Food Funct. 2016, 7, 3224–3232. [Google Scholar] [CrossRef]
- Pozuelo, M.; Panda, S.; Santiago, A.; Mendez, S.; Accarino, A.; Santos, J.; Guarner, F.; Azpiroz, F.; Manichanh, C. Reduction of butyrate- and methane-producing microorganisms in patients with Irritable Bowel Syndrome. Sci. Rep. 2015, 5, 12693. [Google Scholar] [CrossRef] [Green Version]
- Menni, C.; Lin, C.H.; Cecelja, M.; Mangino, M.; Matey-Hernandez, M.L.; Keehn, L.; Mohney, R.P.; Steves, C.J.; Spector, T.D.; Kuo, C.F.; et al. Gut microbial diversity is associated with lower arterial stiffness in women. Eur. Heart J. 2018, 39, 2390–2397. [Google Scholar] [CrossRef] [Green Version]
- Kang, C.; Wang, B.; Kaliannan, K.; Wang, X.; Lang, H.; Hui, S.; Huang, L.; Zhang, Y.; Zhou, M.; Chen, M.; et al. Gut microbiota mediates the protective effects of dietary capsaicin against chronic low-grade inflammation and associated obesity induced by high-fat diet. mBio 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Mancabelli, L.; Milani, C.; Lugli, G.A.; Turroni, F.; Mangifesta, M.; Viappiani, A.; Ticinesi, A.; Nouvenne, A.; Meschi, T.; van Sinderen, D.; et al. Unveiling the gut microbiota composition and functionality associated with constipation through metagenomic analyses. Sci. Rep. 2017, 7, 9879. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Batt, S.M.; Wannemuehler, M.; Dispirito, A.; Beitz, D.C. Effect of feeding of a cholesterol-reducing bacterium, Eubacterium coprostanoligenes, to germ-free mice. Lab. Anim. Sci. 1998, 48, 253–255. [Google Scholar]
- Candela, M.; Perna, F.; Carnevali, P.; Vitali, B.; Ciati, R.; Gionchetti, P.; Rizzello, F.; Campieri, M.; Brigidi, P. Interaction of probiotic Lactobacillus and Bifidobacterium strains with human intestinal epithelial cells: Adhesion properties, competition against enteropathogens and modulation of IL-8 production. Int. J. Food Microbiol. 2008, 125, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Resta-Lenert, S.; Barrett, K.E. Live probiotics protect intestinal epithelial cells from the effects of infection with enteroinvasive Escherichia coli (EIEC). Gut 2003, 52, 988–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masood, M.I.; Qadir, M.I.; Shirazi, J.H.; Khan, I.U. Beneficial effects of lactic acid bacteria on human beings. Crit. Rev. Microbiol. 2011, 37, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Llopis, M.; Antolin, M.; Carol, M.; Borruel, N.; Casellas, F.; Martinez, C.; Espin-Basany, E.; Guarner, F.; Malagelada, J.R. Lactobacillus casei downregulates commensals’ inflammatory signals in Crohn’s disease mucosa. Inflamm. Bowel Dis. 2009, 15, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Picard, C.; Fioramonti, J.; Francois, A.; Robinson, T.; Neant, F.; Matuchansky, C. Review article: Bifidobacteria as probiotic agents—Physiological effects and clinical benefits. Aliment. Pharmacol. Ther. 2005, 22, 495–512. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, X.; Hao, Y.; Ding, J.; Shen, J.; Xue, Z.; Qi, W.; Li, Z.; Song, Y.; Zhang, T.; et al. Protective effects of a novel probiotic strain, Lactococcus lactis ML2018, in colitis: In vivo and in vitro evidence. Food Funct. 2019, 10, 1132–1145. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; Chain, F.; Martin, R.; Bermudez-Humaran, L.G.; Courau, S.; Langella, P. Beneficial effects on host energy metabolism of short-chain fatty acids and vitamins produced by commensal and probiotic bacteria. Microb. Cell Fact. 2017, 16, 79. [Google Scholar] [CrossRef] [Green Version]
- Yao, J. Isolation and molecular identification of the bacterial colonization during the pile fermentation process of Pu-erh tea. J. Anhui Agric. Sci. 2013, 41, 2667–2668. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Identification | Molecular Formula | Retention Time (min) | [M+H]+ (Error, ppm) | [M-H]− (Error, ppm) | Fragment Ions in the Positive Ion Mode (m/z) b | Fragment Ions in the Negative Ion Mode (m/z) b | Source |
|---|---|---|---|---|---|---|---|---|
| Organic Acids and Their Esters | ||||||||
| 1 | Quinic acid | C7H12O6 | 2.60 | ND d | 191.0205 (1.5) | ND | 173.0446 [M-H-H2O]−, 127.0388, 85.0309 | Puerh tea |
| 3 | 3-Glucogallic acid/4-Glucogallic acid | C13H16O10 | 3.14 | ND | 331.0673 (0.7) | ND | 169.0114 [M-H-Glc]−, 125.0219 [M-H-Glc-CO2]− | Puerh tea |
| 4 | Theogallin | C14H16O10 | 3.51 | 345.0817 (−0.3) | 343.0673 (0.8) | 153.0184 [M+H-C7H12O6]+ | 191.0545 [M-H-C7H4O4]−, 169.0113 [M-H-C7H10O5]− | Puerh tea |
| 5 | Gallic acid a | C7H6O5 | 3.76 | 171.0286 (−1.1) | 169.0152 (3.4) | 153.0175 [M+H-H2O]+, 135.0063 [M+H-2H2O]+, 125.0230 [M+H-H2O-CO]+, 107.0135 [M+H-2H2O-CO]+, 81.0356 | 125.0239 [M-H-CO2]− | Puerh tea |
| 7 | 3-Caffeoylquinic acid | C16H18O9 | 5.00 | 355.1022 (3.6) | 353.0876 (0.8) | 163.0385 [M+H-C7H12O6]+, 135.0430 [M+H-C7H12O6-CO]+, | 191.0549 [M-H-C9H7O3]−, 179.0338 [M-H-C7H10O5]−, 135.0441 [M-H-C7H10O5-CO2]− | Puerh tea |
| 9 | Vanillic acid c | C8H8O4 | 5.29 | ND | 167.0357 (3.7) | ND | 152.0104 [M-H-CH3]−, 108.0242 [M-H-C2H5]− | GCP |
| 14 | Caffeoyl-glucose | C15H18O9 | 5.75 | ND | 341.0881 (0.9) | ND | 179.0339 [M-H-Glc]−, 161.0568 [M-H-Glc-H2O]−, 135.0435 [M-H-Glc-CO2]− | GCP, Puerh tea |
| 15 | 3-p-Coumaroylquinic acid | C16H18O8 | 5.83 | 339.1074 (−1) | 337.0932 (1) | 147.0437 [M+H-C7H10O5-H2O]+, 119.0495 [M+H-C9H8O2-CO2-CO]+, 91.0564 [M+H-C7H10O5-HCOOH-CO]+ | 191.0544 [M-H-C9H6O2]−, 163.0384 [M-H-C7H10O5]−, 119.0511 [M-H-C7H10O5-CO2]−, | Puerh tea |
| 16 | 4-Caffeoylquinic acid | C16H18O9 | 5.94 | 355.1021 (0) | 353.0874 (0.8) | 163.0387 [M+H-C7H12O6]+, 145.0277, 135.0425 [M+H-C7H12O6-CO]+, | 191.0547 [M-H-C9H6O3]−, 179.0348 [M-H-C7H10O5]−, 173.0442 [M-H-C9H8O4]−, 135.0456 [M-H- C7H10O5-CO2]−, 93.0364 | Puerh tea |
| 24 | 5-p-Coumaroylquinic acid | C16H18O8 | 6.80 | 339.1071 (−0.6) | 337.0937 (2.5) | 147.0444 [M+H-C7H10O5-H2O]+, 119.0500 [M+H-C9H8O2-CO2-CO]+, 91.0555 [M+H-C7H10O5-HCOOH-CO]+ | 173.0444 [M-H-C9H6O2-H2O]− 163.0384 [M-H-C7H10O5]−, 119.0498 [M-H-C7H10O5-CO2]−, | Puerh tea |
| 25 | Caffeic acid c | C9H8O4 | 6.85 | ND | 179.0362 (4.5) | ND | 135.0443 [M-H-CO2]− | GCP, Puerh tea |
| 26 | 4-p-Coumaroylquinic acid | C16H18O8 | 6.93 | 339.1074 (0.1) | 337.0936 (2.5) | 147.0447 [M+H-C7H10O5-H2O]+, 119.0497 [M+H-C9H8O2-CO2-CO]+, 91.0558 [M+H-C7H10O5-HCOOH-CO]+ | 191.0548 [M-H-C9H6O2]−, 173.0446 [M-H-C9H6O2-H2O]− 163.0366 [M-H-C7H10O5]−, 119.0504 [M-H-C7H10O5-CO2]−, | Puerh tea |
| 38 | p-Coumaric acid | C9H8O3 | 8.23 | ND | 163.0411 (2.2) | ND | 119.0503 [M-H-CO2]− | Puerh tea |
| 46 | Ferulic acid a,c | C10H10O4 | 8.69 | ND | 193.0508 (0.7) | ND | 178.0273 [M-H-CH3]−, 134.0360 [M-H-CH3-CO2]−, | GCP |
| Flavan-3-ols | ||||||||
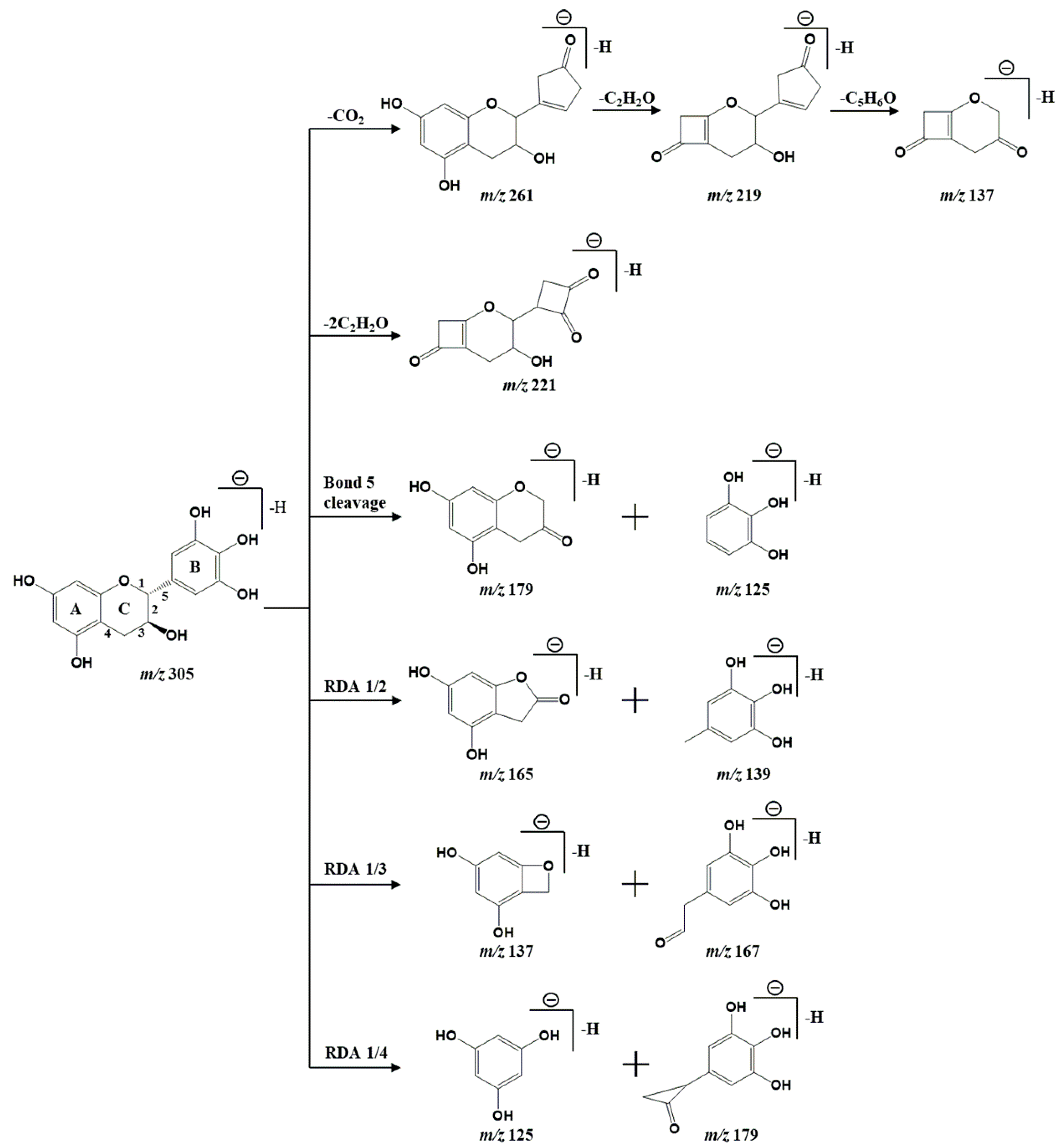
| 6 | Gallocatechin | C15H14O7 | 4.59 | 307.081 (−0.6) | 305.0664 (2.1) | 195.0637 [M+H-CO-2C2H2O]+, 177.0532 [M+H-CO-2C2H2O-H2O]+, 139.0388 [M+H-C8H8O4]+ | 261.0807 [M-H-CO2]−, 221.0413 [M-H-2C2H2O]−, 219.0621 [M-H-CO2-CH2O]−, 179.0447 [M-H-C6H6O3]−, 167.0326 [M-H-C7H6O3]−, 165.0199 [M-H-C7H8O3]−, 139.0398 [M-H-C8H6O4]−, 137.0228 [M-H- CO2-CH2O-C5H6O]−, 125.0236 [M-H-C9H8O4]− | Puerh tea |
| 12 | Epigallocatechin | C15H14O7 | 5.37 | 307.0815 (0.9) | 305.0678 (1.5) | 195.1763 [M+H-CO-2C2H2O]+, 177.0544 [M+H-CO-2C2H2O-H2O]+, 139.0387 [M+H-C8H8O4]+ | 261.0784 [M-H-CO2]−, 221.0439 [M-H-2C2H2O]−, 219.0667 [M-H-CO2-CH2O]−, 179.0350 [M-H-C6H6O3]−, 167.0242 [M-H-C7H6O3]−, 165.0178 [M-H-C7H8O3]−, 137.0234 [M-H- CO2-CH2O-C5H6O]−, 125.0245 [M-H-C9H8O4]− | Puerh tea |
| 17 | Catechin | C15H14O6 | 5.99 | 291.0865 (0.6) | 289.0727 (2.5) | 207.0652 [M+H-C4H4O2]+, 179.0679 [M+H-C4H4O2-CO]+ 139.0396 [M+H-C8H8O3]+, 123.0451 [M+H-C8H8O4]+ | 245.0818 [M-H-CO2]−, 205.0504 [M-H-2C2H2O]−, 203.0703 [M-H-CO2-C2H2O]−, 137.0244 [M-H-C8H8O3]− | Puerh tea |
| 22 | Epicatechin | C15H14O6 | 6.61 | 291.0866 (0.9) | 289.0727 (3.4) | 207.0649 [M+H-C4H4O2]+, 147.0441 [M+H-C4H4O2-C2H2O-H2O]+, 139.0397 [M+H-C8H8O3]+, 123.0452 [M+H-C8H8O4]+ | 245.0839 [M-H-CO2]−, 205.0505 [M-H-2C2H2O]−, 203.0714 [M-H-CO2-C2H2O]−, 161.0595 [M-H-CO2-2C2H2O]−, 151.0397 [M-H-C7H6O3]− | Puerh tea |
| 23 | Epigallocatechin-3-O-gallate | C22H18O11 | 6.76 | 459.0918 (0.2) | 457.0787 (2.2) | 289.0365 [M+H-C7H6O5]+, 139.0388 | 305.0658 [M-H-C7H4O4]−, 169.0123 [C7H5O5]−, 125.0237 [M-H-C7H4O4-C9H8O4]− | Puerh tea |
| 28 | Gallocatechin-3-O-gallate | C22H18O11 | 7.00 | 459.0920 (0) | 457.0777 (1.6) | 289.2269[M+H-C7H6O5]+, 139.1220 | 305.0636 [M-H-C7H4O4]−, 169.0137 [C7H5O5]−, 125.0244 [M-H-C7H4O4-C9H8O4]− | Puerh tea |
| 33 | Epicatechin-3-O-gallate | C22H18O10 | 7.86 | 443.097 (−0.2) | 441.0830 (1.2) | 291.0484 [M+H-C7H4O4]+, 191.0347, 123.0459 | 289.0711 [M-H-C7H4O4]−, 169.0131 [C7H5O5]−, 125.0243 [M-H-C7H4O4-C9H8O3]− | Puerh tea |
| 35 | Catechin-3-O-gallate | C22H18O10 | 8.01 | 443.0972 (0.3) | 441.0824 (0.7) | 291.0884 [M+H-C7H4O4]+, 273.0787, 139.0393, 123.0455 | 289.0710 [M-H-C7H4O4]−, 245.0819 [M-H-C7H4O4-CO2]−, 169.0129 [C7H5O5]−, 125.0244 [M-H-C7H4O4-C9H8O3]− | Puerh tea |
| Alkaloids | ||||||||
| 2 | Synephrine a,c C9H13NO2 | 2.72 | 168.1014 (−4.7) | ND | 150.0913 [M+H-H2O]+, 135.0681 [M+H-H2O-CH3]+, 107.0500 [M+H- | ND | GCP | |
| H2O-CH3-CO]+, 91.0561, 77.0409, 65.0417 | ||||||||
| 8 | Theobromine/Theophylline | C7H8N4O2 | 5.21 | 181.0721 (0.4) | 179.0585 (5.7) | 124.0500 [M+H-C2H3NO] +,96.0567 [M+H-C2H3NO-CO]+ | 164.0332 [M-H-CH3]−, 122.0354 [M-H-C2H3NO]− | Puerh tea |
| 11 | 8-Oxocaffeine | C8H10N4O3 | 5.36 | 211.0824 (−0.9) | 209.0689 (2.9) | 196.0589 [M+H-CH3]+, 154.0581 [M+H-C2H3NO]+, | 194.0432 [M-H-CH3]−, 137.0218 [M-H-C2H3NO-CH3]− | Puerh tea |
| 19 | Caffeine a | C8H10N4O2 | 6.32 | 195.0877 (0.2) | ND | 138.0658 [M+H-C2H3NO]+, 123.0425 [M+H-C2H3NO-CH3]+, 110.0714 [M+H-C2H3NO-CO]+ | ND | Puerh tea |
| 53 | Citrusin III | C36H53N7O9 | 11.84 | 728.3982 (0.6) | 726.3872 (4.4) | 700.4052 [M+H-CO]+, 587.3161, 474.2319 | 696.3815, 590.3348 | GCP |
| 60 | Citrusin I | C34H53N7O9 | 13.74 | 704.3975 (−0.4) | ND | 686.3833 [M+H-H2O]+ | ND | GCP |
| Limonoids | ||||||||
| 75 | Limonin c | C26H30O8 | 16.65 | 471.2013 (0) | ND | 425.1982 [M+H-CH2O2]+, 161.0631 | ND | GCP |
| Flavonoids | ||||||||
| 10 | Ampelopsin | C15H12O8 | 5.33 | 321.0605 (−0.1) | 319.046 (0.3) | 183.0294 [M+H-C7H6O3]+, 139.0374 [M+H-C8H6O4]+ | 137.0239 [M-H-C8H6O4]− | Puerh tea |
| 13 | Luteolin-6,8-di-C-glucoside c | C27H30O16 | 5.73 | 611.1599 (−1) | 609.1477 (2.6) | 593.1499 [M+H-H2O]+, 575.1404 [M+H-2H2O]+, 557.1410 [M+H−3H2O]+, 473.1021 [M+H-C4H8O4-H2O]+, 353.0650 [M+H-2C4H8O4-H2O]+ | 489.1057 [M-H-C4H8O4]−, 399.0745 [M-H-C4H8O4-C3H6O3]−, 369.0630 [M-H-2C4H8O4]− | GCP |
| 18 | Vicenin-2 c | C27H30O15 | 6.21 | 595.1659 (0.2) | 593.1531 (3.3) | 577.1542 [M+H-H2O]+, 559.1450 [M+H-2H2O]+ 541.1356 [[M+H-3H2O]+ | 503.1220 [M-H-C3H6O3]−, 473.1112 [M-H-C4H8O4]−, 383.0790 [M-H-C4H8O4-C3H6O3]−, 353.0681 [M-H-2C4H8O4]− | GCP |
| 20 | Chrysoeriol-6,8-di-C-glucoside | C28H32O16 | 6.41 | 625.1762 (−0.3) | 623.1641 (3.8) | 607.1664 [M+H-H2O]+, 589.1550 [M+H-2H2O]+, 571.1434 [M+H-3H2O]+, 487.1228 [M+H-C4H8O4-H2O]+, 367.0839 [M+H-2C4H8O4-H2O]+ | 503.1236 [M-H-C4H8O4]−, 413.0898 [M-H-C4H8O4-C3H6O3]−, 383.0794 [M-H-2C4H8O4]− | GCP |
| 21 | Narirutin c | C27H32O14 | 6.43 | 581.1855 (−1.8) | 579.1376 (−1.1) | 273.0742 [M+H-Glc-Rha]+, | ND | GCP |
| 27 | Isoorientin c | C21H20O11 | 6.95 | 449.1079 (0.2) | 447.0937 (1) | 431.0971 [M+H-H2O]+, 413.0877 [M+H-2H2O]+, 395.0755 [M+H-3H2O]+, 329.0657 [M+H-C4H8O4]+, 299.0556 [M+H-C8H6O3]+ | 357.0623 [M-H-C3H6O3]−, 327.0505 [M-H-C4H8O4]− | GCP |
| 29 | Orientin c | C21H20O11 | 7.18 | 449.1075 (−0.7) | 447.0938 (2.1) | 431.0972 [M+H-H2O]+, 413.0860 [M+H-2H2O]+, 395.0802 [M+H-3H2O]+, 329.0644 [M+H-C4H8O4]+, 299.0561 [M+H-C8H6O3]+ | 357.0623 [M-H-C3H6O3]−, 327.0505 [M-H-C4H8O4]− | GCP |
| 30 | Rutin a,c | C27H30O16 | 7.54 | 611.1607 (0) | 609.1479 (2.9) | 465.1023 [M+H-Rha]+, 303.0498 [M+H-Rha-Glc]+ | 301.0352 [M-H-Rha-Glc]− | GCP, Puerh tea |
| 31 | Lonicerin c | C27H30O15 | 7.60 | 595.1656 (−0.3) | 593.1534 (3) | 449.1063 [M+H-Rha]+, 287.0548 [M+H-Glc-Rha]+ | 285.0413 [M-H-Glc-Rha]− | GCP, Puerh tea |
| 32 | Apigenin-8-C-glucoside | C21H20O10 | 7.74 | 433.1128 (−0.2) | 431.0992 (1.8) | 415.1031 [M+H-H2O]+, 397.0910 [M+H-2H2O]+, 379.0809 [M+H-3H2O]+, 313.0705 [M+H-C4H8O4]+, 283.0601 [M+H-C8H6O3]+ | 341.0674 [M-H-C3H6O3]−, 311.0567 [M-H-C4H8O4]−, 283.0617 [M-H-C4H8O4-CO]− | GCP |
| 34 | Quercetin-3-O-glucoside | C21H20O12 | 7.95 | 465.1031 (0.7) | 463.0887 (1) | 303.0497 [M+H-Glc]+ | 301.0356 [M-H-Glc]−, 271.0249, 151.0024 [M-H-Glc-C8H6O3]− | Puerh tea |
| 36 | Diosmetin-6-C-glucoside | C22H22O11 | 8.08 | 463.1237 (−0.2) | 461.1093 (0.8) | 445.1231 [M+H-H2O]+, 427.1036 [M+H-2H2O]+, 409.0924 [M+H-3H2O]+, 343.0801 [M+H-C4H8O4]+, 313.0700 [M+H-C8H6O3]+ | 371.0829 [M-H-C3H6O3]−, 341.0686 [M-H-C4H8O4]−, 298.0489 [M-H-C4H8O4-CO-CH3]− | GCP |
| 37 | Kaempferol-3-O-rutinoside | C27H30O15 | 8.15 | 595.1652 (−0.1) | 593.1529 (2.9) | 449.1063 [M+H-Rha]+, 287.0548 [M+H-Rha-Glc]+ | 285.0406 [M-H-Glc-Rha]− | Puerh tea |
| 39 | Naringin a,c | C27H32O14 | 8.25 | 581.1868 (0.6) | 597.1743 (4.1) | 419.1312 [M+H-Glc]+, 273.0762 [M+H-Rha-Glc]+, 153.0179 [M+H-Rha-Glc-C8H8O]+ | 271.0615 [M-H-Glc-Rha]−, 151.0027 [M-H-Rha-Glc-C8H8O]− | GCP |
| 40 | Rhoifolin a,c | C27H30O14 | 8.30 | 579.1713 (0.8) | 577.158 (2.9) | 433.1121 [M+H-Rha]+, 271.0599 [M+H-Rha-Glc]+ | 269.0455 [M-H-Rha-Glc]− | GCP |
| 41 | Diosmin c | C28H32O15 | 8.47 | 609.1868 (−0.4) | 607.1687 (3.4) | 299.0566 [M-H-Glc-Rha]−, 284.0333 [M-H-Glc-Rha-CH3]− | GCP | |
| 42 | Kaempferol-3-O-glucoside | C21H20O11 | 8.59 | 449.108 (0.4) | 447.0944 (1) | 287.0561 [M+H-Glc]+ | 284.0329 [M-H-Glc]−, 255.0300 [M-H-Glc-CHO]−, 227.0344 [M-H-Glc-CHO-CO]−, | Puerh tea |
| 43 | Neodiosmin | C28H32O15 | 8.60 | 609.1811 (−0.4) | 607.1684 (2.6) | 463.1253 [M+H-Rha]+, 301.0717 [M+H-Rha-Glc]+, 286.0465 [M+H-Rha-Glc-CH3]+ | 299.0569 [M-H-Glc-Rha]−, 284.0325 [M-H-Glc-Rha-CH3]− | GCP |
| 44 | Hesperidin a,c | C28H34O15 | 8.64 | 611.1968 (−0.3) | 609.1849 (3.7) | 449.1425 [M+H-Glc]+, 303.0864 [M+H-Rha-Glc]+, 153.0181 [M+H-Rha-Glc-C9H10O2]+ | 301.0729 [M-H-Rha-Glc]−, 286.0496 [M-H-Rha-Glc-CH3]− | GCP |
| 45 | Homoeriodictyol c | C16H14O6 | 8.68 | 303.0863 (0.1) | 301.0722 (0.6) | 153.0177 [M+H-C9H10O2]+, 117.0337 [M+H-C9H10O2-2H2O]+ | 286.0511 [M-H-CH3]−, 151.0032 [M-H-C9H10O2]− | GCP |
| 47 | 5,3′-Dihydroxy-7,4′-dimethoxyflavone c | C17H14O6 | 9.54 | 315.0856 (−2) | ND | 300.0617 [M+H-CH3]+, 285.0427 [M+H-2CH3]+, | ND | GCP |
| 48 | Myricetin | C15H10O8 | 9.57 | 319.0447 (−0.4) | 317.0312 (2.3) | 273.0387 [M+H-H2O-CO]+, 245.0457 [M+H-H2O-2CO]+, 153.0185 [M+H-C8H6O4]+ | 271.0213 [M-H-H2O-CO]−, 178.9975 [M-H-C7H6O3]−, 151.0024 [M-H-C8H6O4]− | Puerh tea |
| 49 | Poncirin a | C28H34O14 | 10.63 | 595.2021 (0) | 593.1903 (3.8) | 449.1415 [M+H-Rha]+, 287.0914 [M+H-Glc-Rha]+, 153.0170 [M+H-Glc-Rha-C9H10O]+ | 285.0768 [M-H-Glc-Rha]− | GCP |
| 50 | Isosakuranetin c | C16H14O5 | 10.69 | 287.0913 (−0.5) | 285.0775 (2.8) | 153.0173 [M+H-C9H10O]+, 133.0633 [M+H-C7H4O4]+ | 243.0632 [M-H-C2H3O]−, | GCP |
| 51 | Luteolin | C15H10O6 | 11.40 | 287.0549 (−0.4) | 285.0409 (1.1) | 153.0203 [M+H-C8H6O2]+ | 133.0290 [M-H-C7H4O4]− | Puerh tea |
| 52 | Quercetin c | C15H10O7 | 11.51 | 303.0501 (0.4) | 301.0365 (3.8) | 285.0372 [M+H-H2O]+, 257.0441 [M+H-H2O-CO]+, 229.0498 [M+H-H2O-2CO]+, 153.0177 [M+H-C8H6O3]+ | 178.9976 [M-H-C7H6O2]−, 151.0027 [M-H-C8H6O3]− | Puerh tea |
| 54 | Monohydroxy-trimethoxyflavone | C18H16O6 | 12.25 | 329.1019 (−0.1) | ND | 314.0820 [M+H-CH3]+, 299.0540 [M+H-2CH3]+, 271,0580 [M+H-2CH3-CO]+, 181.0096, 153.0138 | ND | GCP |
| 55 | 7-Hydroxy-3,5,6,8-tetramethoxyflavone | C19H18O7 | 12.69 | 359.113 (1.3) | ND | 344.0912 [M+H-CH3]+, 329.0663 [M+H-2CH3]+ | ND | GCP |
| 56 | Naringenin a,c | C15H12O5 | 13.16 | 273.0758 (0.1) | 271.0626 (2.5) | 153.0185 [M+H-C8H8O]+, 147.0442, 119.0500 | 151.0032 [M-H-C8H8O]−, 119.0505 [M-H-C7H4O4]−, 107.0143 [M-H-C8H8O-CO2]− | GCP |
| 57 | Apigenin a, c | C15H10O5 | 13.30 | 271.0601 (0.1) | 269.0466 (3.9) | 153.0175 [M+H-C8H6O]+ | 151.003 [M-H-C8H6O]+ | Puerh tea |
| 58 | 7-Hydroxy-5,6,8,4′-tetramethoxyflavone c | C19H18O7 | 13.65 | 359.1125 (−0.1) | ND | 344.0851 [M+H-CH3]+, 329.0633 [M+H-2CH3]+, 326.0779 [M+H-CH3-H2O]+, 298.0814 [M+H-CH3-CO-H2O]+, | ND | GCP |
| 59 | Kaempferol a,c | C15H10O6 | 13.67 | 287.055 (0.1) | 285.0413 (4.5) | 153.0170 [M+H-C8H6O2]+ | Puerh tea | |
| 61 | Hesperetin a,c | C16H14O6 | 13.76 | 303.0865 (0.7) | 301.0724 (3.4) | 177.0546, 153.0177 [M+H-C9H10O2] + | 286.0484 [M-H-CH3]−, 151.0022 [M-H-C9H10O2]− | GCP |
| 62 | Chrysoeriol c | C16H12O6 | 13.76 | 301.0709 (0.9) | 299.0571 (3.2) | 286.0466 [M+H-CH3]+, 258.0526 [M+H-CH3-CO]+, 229.0485 [M+H-CO2-CO]+, 153.0152 [M+H-C9H8O2]+ | 284.0328 [M-H-CH3]−, 256.0393 [M-H-CH3-CO]−, 227.0352 [M-H-CO2-CO]−, 151.0010 [M-H-C9H8O2]− | GCP |
| 63 | 5-Hydroxy-3,6,7,8-tetramethoxyflavone | C19H18O7 | 14.12 | 359.1126 (0.3) | ND | 344.0904 [M+H-CH3]+, 329.0614 [M+H-2CH3]+ 298.0815 [M+H-CH3-CO-H2O]+ | ND | GCP |
| 64 | 5,6,7,3′,4′-Pentamethoxyflavanone | C20H22O7 | 14.36 | 375.1441 (0.8) | ND | 211.0594 [M+H-C10H10O2]+, 196.0361 [M+H-C10H10O2-CH3]+, 150.0311 [M+H-C10H10O2-CO-CH3-H2O]+ | ND | GCP |
| 65 | 7-Hydroxy-5,6,8,3′,4′-pentamethoxyflavone | C20H20O8 | 14.52 | 389.1233 (0.4) | ND | 374.0990 [M+H-CH3]+, 359.0759 [M+H-2CH3]+, 341.0635 [M+H-2CH3-H2O]+, 197.0073 [M+H-C10H10O2-2CH3]+ | ND | GCP |
| 66 | 3′-Hydroxy-5,6,7,8,4′-pentamethoxyflavone/4′-Hydroxy-5,6,7, 8,3′-pentamethoxyflavone | C20H20O8 | 14.53 | 389.1233 (0.4) | ND | 374.0990 [M+H-CH3]+, 359.0759 [M+H-2CH3]+, 344.0635 [M+H-3CH3]+ | ND | GCP |
| 67 | 6-Hydroxy-5,7,8,4′-tetramethoxyflavone | C19H18O7 | 14.80 | 359.1124 (−0.3) | ND | 344.0866 [M+H-CH3]+, 329.0637 [M+H-2CH3]+, 314.0393 [M+H-3CH3]+, 183.0314 | ND | GCP |
| 68 | Isosinensetin c (3′,4′,5,7,8-pentamethoxyflavone) | C20H20O7 | 15.16 | 373.1285 (0.9) | ND | 358.1048 [M+H-CH3]+, 343.0811 [M+H-2CH3]+, 315.0865 [M+H-2CH3-CO]+ | ND | GCP |
| 69 | Monohydroxy-hexamethoxyflavone | C21H22O9 | 15.35 | 419.1335 (−0.4) | ND | 404.1055 [M+H-CH3]+, 389.0878 [M+H-2CH3]+ | ND | GCP |
| 70 | Monohydroxy-pentamethoxyflavanone | C20H22O8 | 15.49 | 391.1385 (−0.6) | ND | 241.0709 [M+H-C9H10O2]+, 226.0452 [M+H-C9H10O2-CH3]+, 211.0249 [M+H-C9H10O2-2CH3]+, 183.0300 [M+H-C9H10O2-2CH3-CO]+ | ND | GCP |
| 71 | 5-Hydroxy-6,7,8,4′-tetramethoxyflavone | C19H18O7 | 15.51 | 359.1133 (2.2) | ND | 329.0656 [M+H-2CH3]+ | ND | GCP |
| 72 | 5-Hydroxy-7,8,3′,4′-tetramethoxyflavone | C19H18O7 | 16.21 | 359.1129 (1) | ND | 344.0908 [M+H-CH3]+, 329.0672 [M+H-2CH3]+, 311.0543 [M+H-2CH3-H2O]+, 197.0043 [M+H-C10H10O2]+,169.0114 [M+H-C10H10O2-CO]+ | ND | GCP |
| 73 | 5,7,3′,4′-Tetramethoxyflavone | C19H18O6 | 16.22 | 343.1178 (0.5) | ND | 328.0944 [M+H-CH3]+, 327.0881 [M+H-CH4]+, 312.0610 [M+H-CH4-CH3]+, 299.0894 [M+H-CH4-CO]+, 283.0562 [M+H-2CH4-CO]+ | ND | GCP |
| 74 | Sinensetin a,c | C20H20O7 | 16.60 | 373.1287 (1.3) | ND | 358.1072 [M+H-CH3]+, 357.0983 [M+H-CH4]+, 343.0829 [M+H-2CH3]+, 315.0868 [M+H-2CH3-CO]+ | ND | GCP |
| 76 | 5,6,7,4′-Tetramethoxyflavone | C19H18O6 | 16.83 | 343.1178 (0.5) | ND | 328.0946 [M+H-CH3]+, 327.0846 [M+H-CH4]+, 313.0710 [M+H-2CH3]+, 299.0918 [M+H-CH4-CO]+, 285.0763 [M+H-2CH3-CO]+, 153.0185 | ND | GCP |
| 77 | 5,7,8,3′,4′-Pentamethoxyflavanone | C20H22O7 | 16.95 | 375.1438 (0) | ND | 211.0594 [M+H-C10H10O2]+, 196.0362 [M+H-C10H10O2-CH3]+, 168.0406 [M+H-C11H10O3-CH3]+ | ND | GCP |
| 78 | Dihydroxy-trimethoxyflavone | C18H16O7 | 17.57 | 345.0972 (1) | ND | 330.0747 [M+H-CH3]+, 315.0490 [M+H-2CH3]+, 301.0706 [M+H-CO2]+ | ND | GCP |
| 79 | 5,6,7,8,3′,4′-Hexamethoxyflavanone | C21H24O8 | 17.76 | 405.1547 (0.8) | ND | 241.0705 [M+H-C10H10O2]+, 226.0464 [M+H-C10H10O2-CH3]+, 211.0233 [M+H-C10H10O2-2CH3]+, 183.0287 | ND | GCP |
| 80 | 5,7,4′-Trimethoxyflavone | C18H16O5 | 18.07 | 313.1084 (4.4) | ND | 298.0896 [M+H-CH3]+, 270.0929 [M+H-CO-CH3]+, 269.0823 [M+H-CO2]+ | ND | GCP |
| 81 | Nobiletin a,c | C21H22O8 | 18.09 | 403.1391 (0.8) | ND | 388.1145 [M+H-CH3]+, 373.0905 [M+H-2CH3]+, 358.0677 [M+H-3CH3]+, 327.0853 [M+H-3CH3-OCH3]+ | ND | GCP |
| 82 | Dihydroxy-tetramethoxyflavone | C19H18O8 | 18.16 | 375.1072 (0.9) | ND | 360.0817 [M+H-CH3]+, 345.0568 [M+H-2CH3]+, 330.0371 [M+H-3CH3]+, 327.0484 [M+H-2CH3-H2O]+, 197.0088 | ND | GCP |
| 83 | 5,7,8,4′-Tetramethoxyflavone | C19H18O6 | 18.39 | 343.118 (1.2) | ND | 327.0862 [M+H-CH4]+, 313.0710 [M+H-2CH3]+, 285.0751 [M+H-2CH3-CO]+, 282.0890 [M+H-2CH3-OCH3]+, 153.0179 | ND | GCP |
| 84 | Monohydroxy-tetramethoxyflavanone | C19H20O7 | 18.88 | 361.1284 (0.5) | ND | 197.0425 [M+H-C10H12O2]+, 182.0205 [M+H-C10H12O2-CH3]+, 136.0151 | ND | GCP |
| 85 | 3,5,6,7,8,3′,4′-Heptemethoxyflavone | C22H24O9 | 19.08 | 433.1496 (0.8) | ND | 418.1268 [M+H-CH3]+, 403.1024 [M+H-2CH3]+, 385.0925 [M+H-2CH3-H2O]+, 345.0610 [M+H-4CH3-CO]+ | ND | GCP |
| 86 | 5-Hydroxy-6,7,8,3′,4′-pentamethoxyflavanone | C20H22O8 | 19.56 | 391.139 (0.7) | ND | 227.0535 [M+H-C10H12O2]+, 212.0306 [M+H-C10H12O2-CH3] +,149.0224 | ND | GCP |
| 87 | Monohydroxy-tetramethoxyflavone | C19H18O7 | 19.65 | 359.1124 (−0.3) | ND | 344.0900 [M+H-CH3]+, 326.0762 [M+H-CH3-H2O]+, 298.0828 [M+H-CH3-CO-H2O]+, 162.0676 | ND | GCP |
| 88 | Tangeretin a,c | C20H20O7 | 19.88 | 373.1286 (1.1) | ND | 358.1049 [M+H-CH3]+, 343.0810 [M+H-2CH3]+, 328.0584 [M+H-3CH3]+, 325.0715 [M+H-2CH3-H2O]+, 315.0868 [M+H-2CH3-CO]+ | ND | GCP |
| 89 | Monohydroxy-tetramethoxyflavone | C19H18O7 | 20.50 | 359.1125 (0) | ND | 344.0867 [M+H-CH3]+, 343.0808 [M+H-CH4]+, 315.0845 [M+H-CO2]+, 164.0841 | ND | GCP |
| 90 | 5-Hydroxy-6,7,8,3′,4′-pentamethoxyflavone | C20H20O8 | 21.20 | 389.1234 (0.9) | ND | 374.0984 [M+H-CH3]+, 359.0756 [M+H-2CH3]+, 341.0658 [M+H-2CH3-H2O]+, 197.0088 [M+H-C10H12O2-2CH3]+ | ND | GCP |
| 91 | Natsudaidain | C21H22O9 | 22.26 | 419.1337 (0.1) | ND | 404.1168 [M+H-CH3]+, 389.0870 [M+H-2CH3]+, 371.0800 [M+H-2CH3-H2O]+ | ND | GCP |
| 92 | Monohydroxy-tetramethoxyflavone | C19H18O7 | 23.02 | 359.1127 (−0.3) | ND | 344.0886 [M+H-CH3]+, 329.0654 [M+H-2CH3]+, 311.0552 [M+H-2CH3-H2O]+, 197.0069 | ND | GCP |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Y.; Zeng, X.; Chen, T.; Peng, W.; Su, W. Chemical Profile, Antioxidative, and Gut Microbiota Modulatory Properties of Ganpu Tea: A Derivative of Pu-erh Tea. Nutrients 2020, 12, 224. https://doi.org/10.3390/nu12010224
Zheng Y, Zeng X, Chen T, Peng W, Su W. Chemical Profile, Antioxidative, and Gut Microbiota Modulatory Properties of Ganpu Tea: A Derivative of Pu-erh Tea. Nutrients. 2020; 12(1):224. https://doi.org/10.3390/nu12010224
Chicago/Turabian StyleZheng, Yuying, Xuan Zeng, Tingting Chen, Wei Peng, and Weiwei Su. 2020. "Chemical Profile, Antioxidative, and Gut Microbiota Modulatory Properties of Ganpu Tea: A Derivative of Pu-erh Tea" Nutrients 12, no. 1: 224. https://doi.org/10.3390/nu12010224
APA StyleZheng, Y., Zeng, X., Chen, T., Peng, W., & Su, W. (2020). Chemical Profile, Antioxidative, and Gut Microbiota Modulatory Properties of Ganpu Tea: A Derivative of Pu-erh Tea. Nutrients, 12(1), 224. https://doi.org/10.3390/nu12010224