Gut Microbiota Modulation by Dietary Barley Malt Melanoidins †


 and
and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Animals
2.2. Experimental Design
2.3. Fecal Short Chain Fatty Acids (SCFAs) Quantification
2.4. DNA Extraction and PCR Amplification
2.5. Library Preparation and Sequencing
2.6. Bioinformatics and Statistical Analyses
3. Results
3.1. Impact of Diet on Feed Intake and Average Daily Gain
3.2. SCFAs Abundance and Dynamics
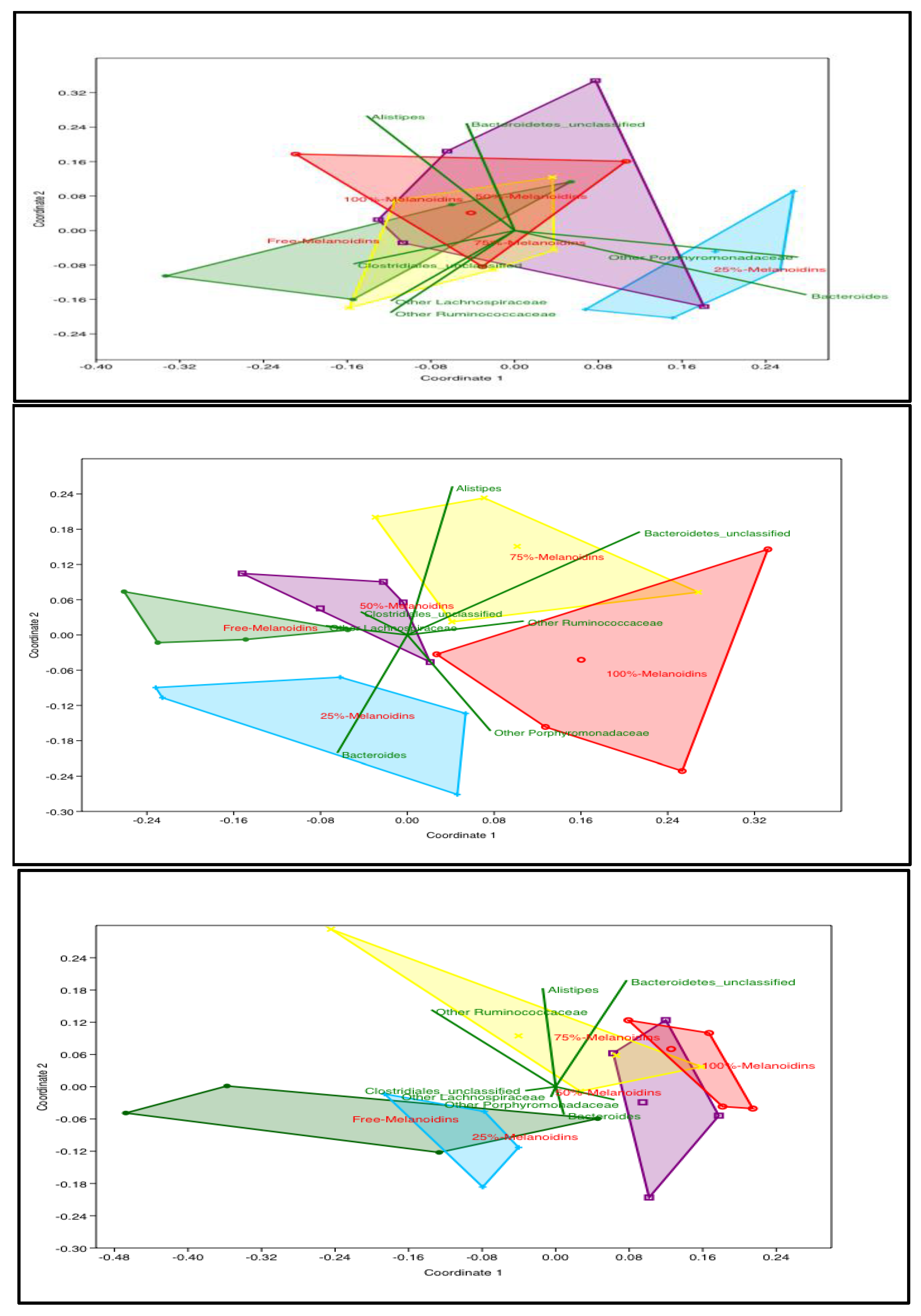
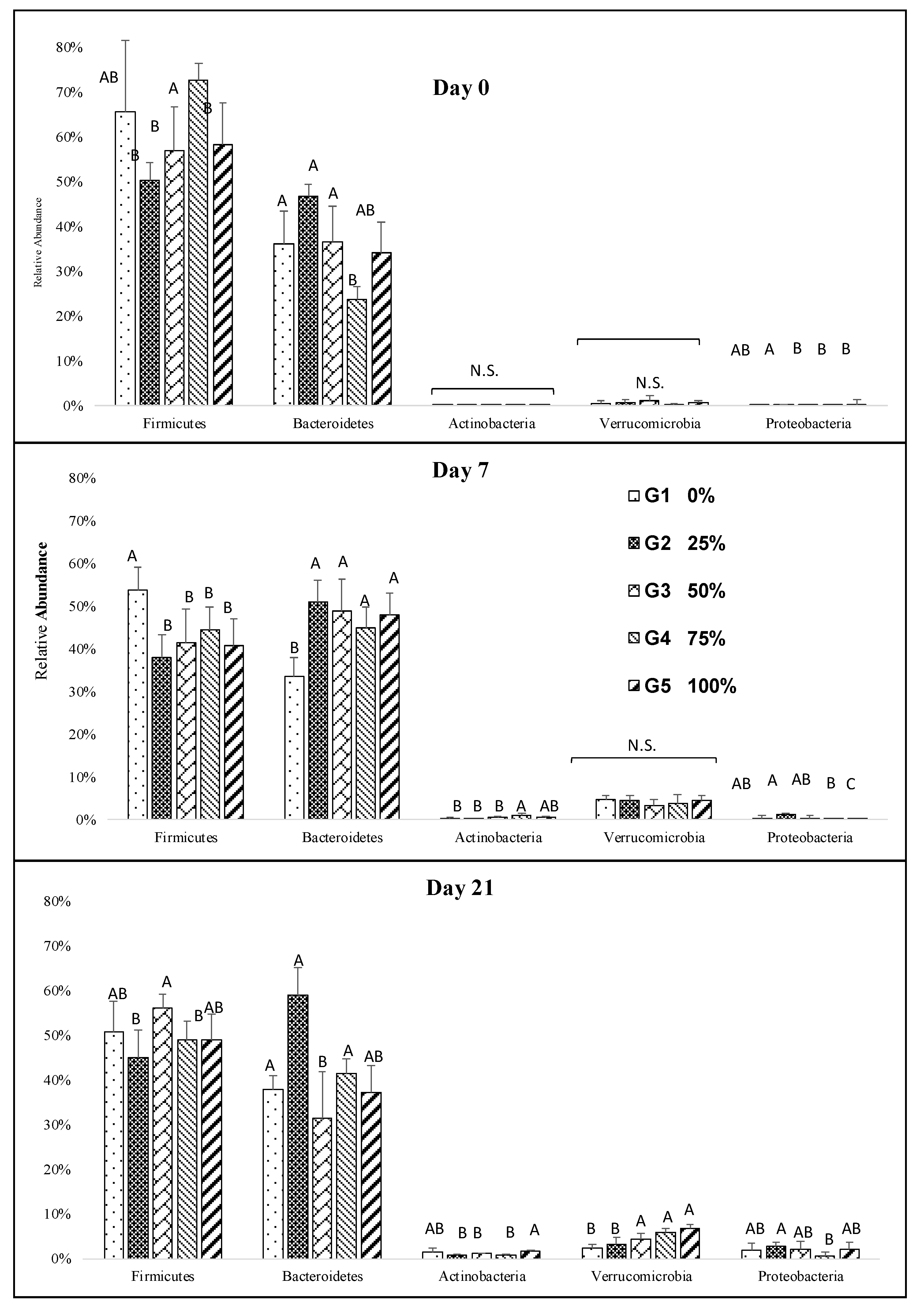
3.3. Impact of Diet on Gut Microbiota Profiles and Dynamics
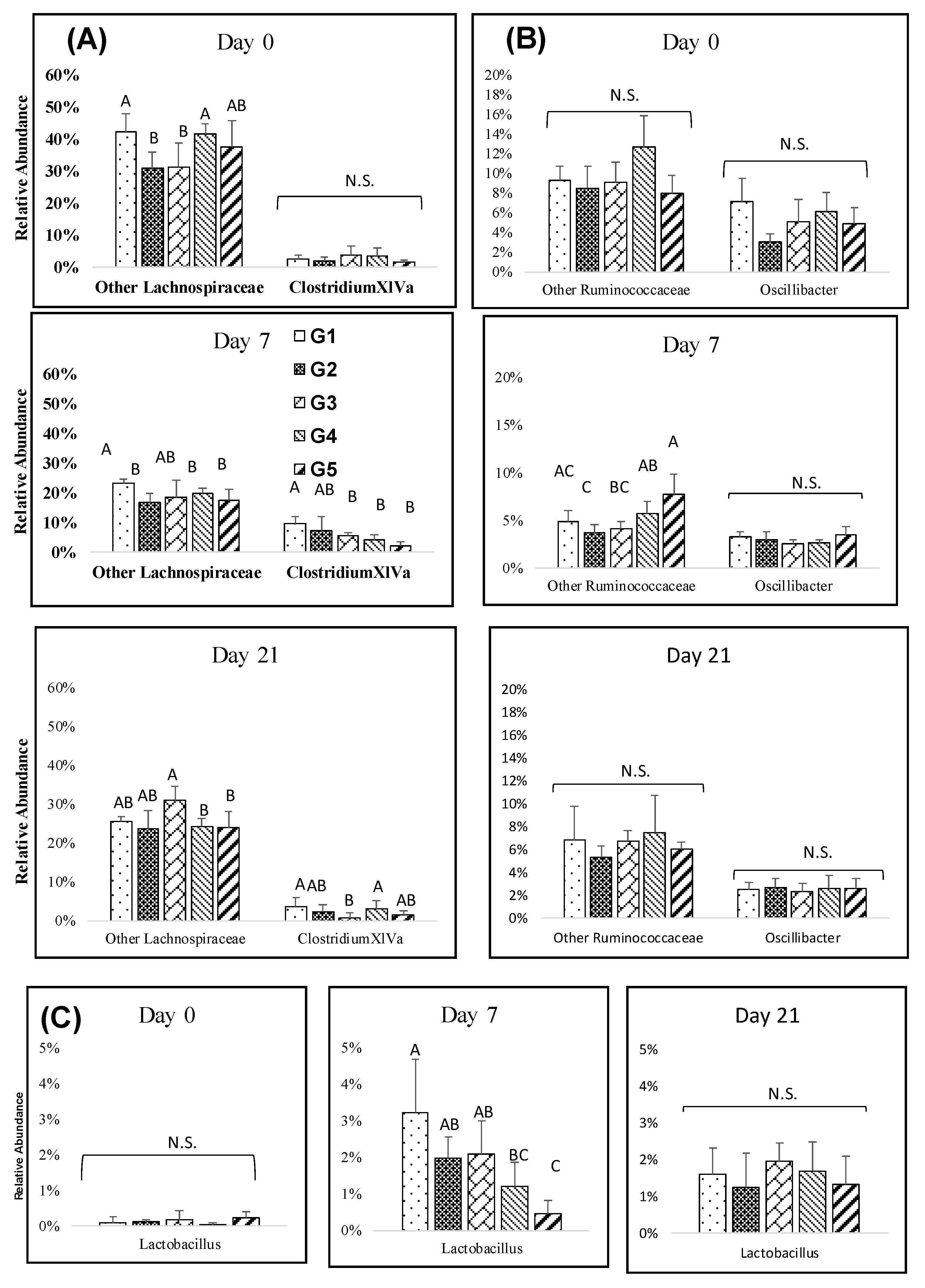
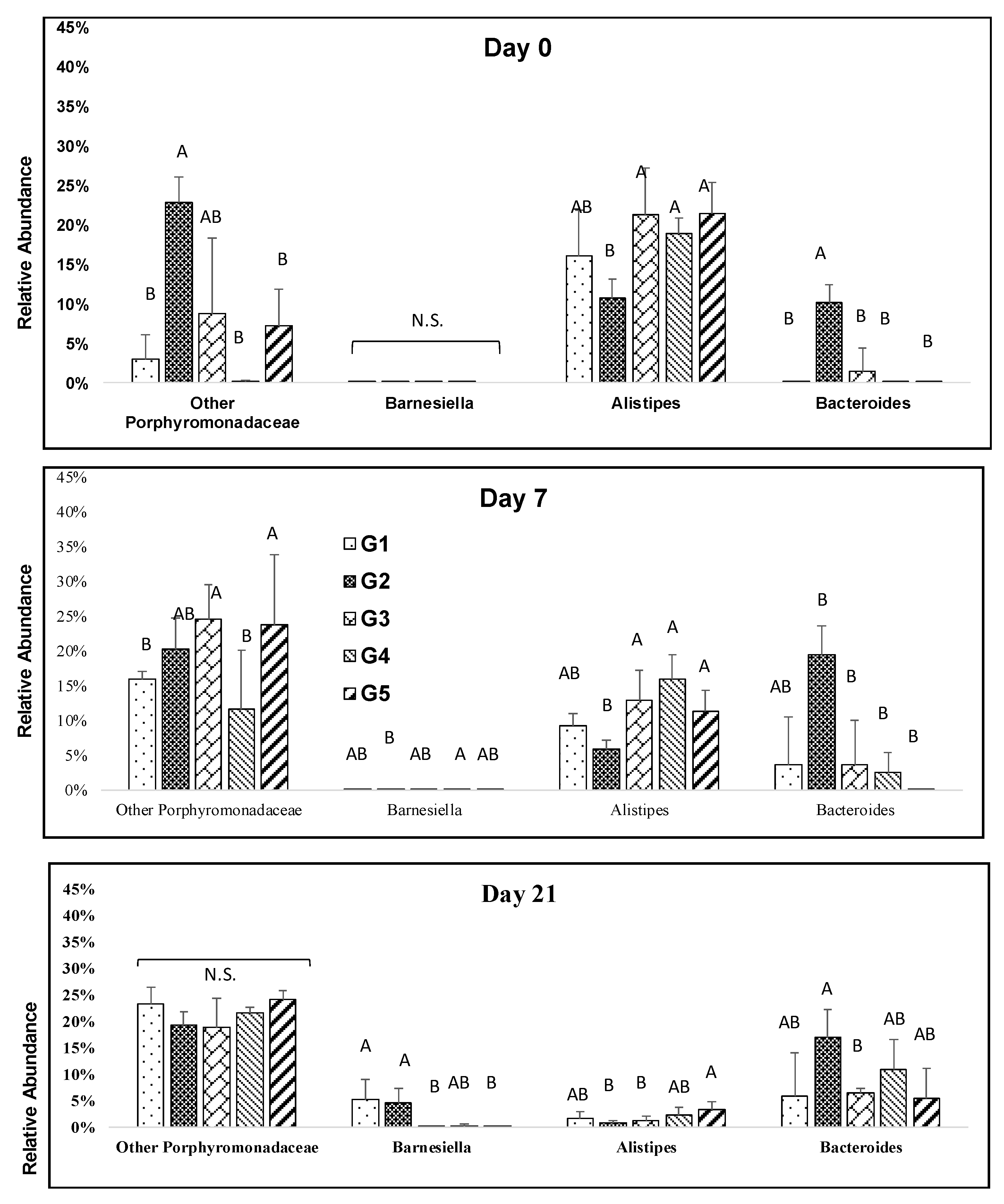
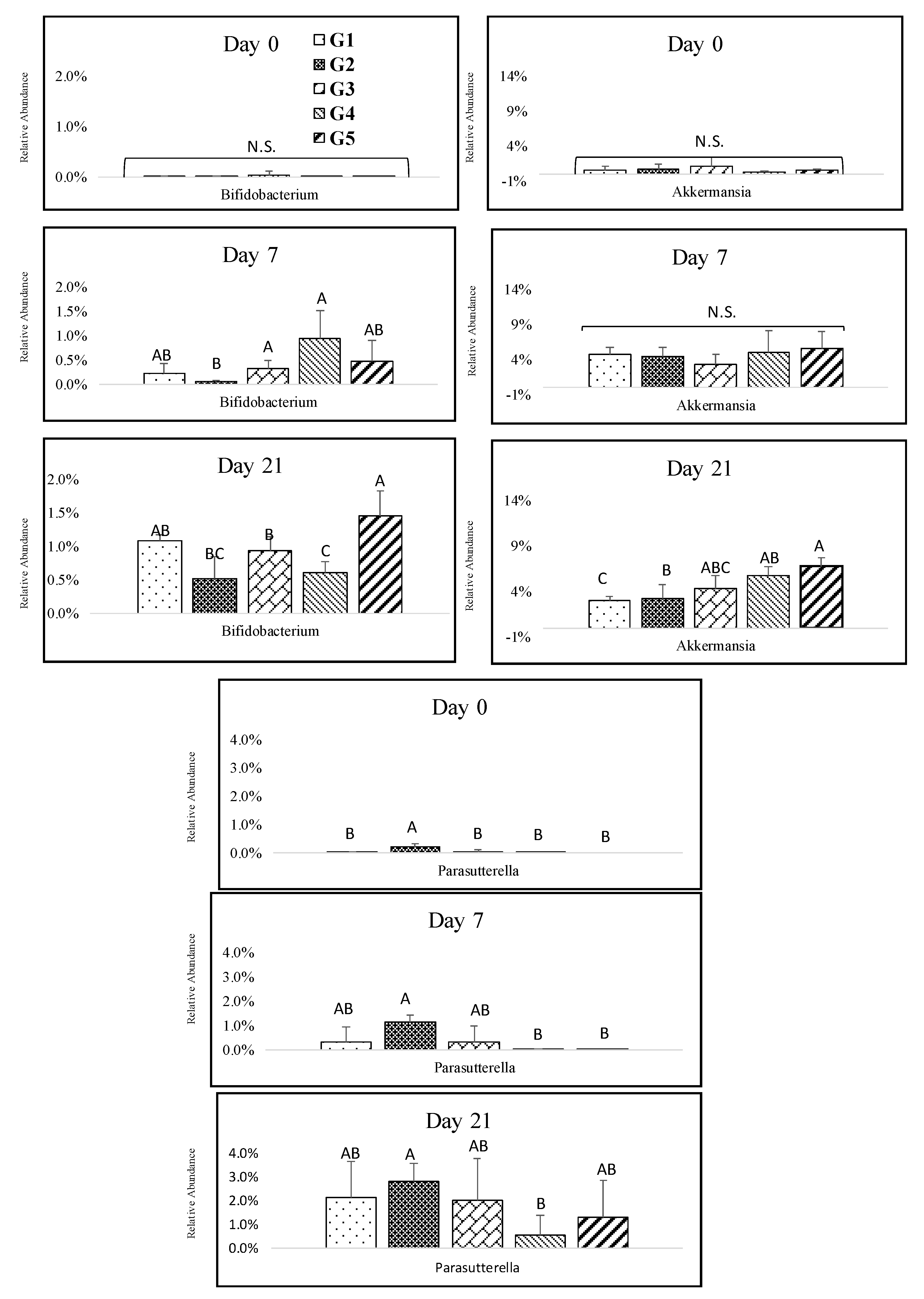
3.4. Impact of Melanoidin Malts on the Composition of the Gut Microbiota
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bekedam, E.K.; Roos, E.; Schols, H.A.; Van Boekel, M.A.J.S.; Smit, G. Low molecular weight melanoidins in coffee brew. J. Agric. Food Chem. 2008, 56, 4060–4067. [Google Scholar] [CrossRef]
- Hodge, J.E. Dehydrated foods, chemistry of browning reactions in model systems. J. Agric. Food Chem. 1953, 1, 928–943. [Google Scholar] [CrossRef]
- Hofmann, T. Characterization of the most intense coloured compounds from Maillard reactions of pentoses by application of colour dilution analysis. Carbohydr. Res. 1998, 313, 203–213. [Google Scholar] [CrossRef]
- Ćosović, B.; Vojvodić, V.; Bošković, N.; Plavšić, M.; Lee, C. Characterization of natural and synthetic humic substances (melanoidins) by chemical composition and adsorption measurements. Org. Geochem. 2010, 41, 200–205. [Google Scholar] [CrossRef]
- Wang, H.-Y.; Qian, H.; Yao, W.-R. Melanoidins produced by the Maillard reaction: Structure and biological activity. Food Chem. 2011, 128, 573–584. [Google Scholar] [CrossRef]
- Coghe, S.; Adriaenssens, B.; Leonard, S.; Delvaux, F.R. Fractionation of colored Maillard reaction products from dark specialty malts. J. Am. Soc. Brew. Chem. 2004, 62, 79–86. [Google Scholar] [CrossRef]
- Milic, B.L.; Grujic-Injac, B.; Piletic, M.V.; Lajsic, S.; Kolarov, L.A. Melanoidins and carbohydrates in roasted barley. J. Agric. Food Chem. 1975, 23, 960–963. [Google Scholar] [CrossRef] [PubMed]
- ALjahdali, N.; Carbonero, F. Impact of Maillard reaction products on nutrition and health: Current knowledge and need to understand their fate in the human digestive system. Crit. Rev. Food Sci. Nutr. 2019, 59, 474–487. [Google Scholar] [CrossRef]
- Glösl, S.; Wagner, K.-H.; Draxler, A.; Kaniak, M.; Lichtenecker, S.; Sonnleitner, A.; Somoza, V.; Erbersdobler, H.; Elmadfa, I. Genotoxicity and mutagenicity of melanoidins isolated from a roasted glucose–glycine model in human lymphocyte cultures, intestinal Caco-2 cells and in the Salmonella typhimurium strains TA98 and TA102 applying the AMES test. Food Chem. Toxicol. 2004, 42, 1487–1495. [Google Scholar] [CrossRef]
- Taylor, J.L.S.; Demyttenaere, J.C.R.; Abbaspour Tehrani, K.; Olave, C.A.; Regniers, L.; Verschaeve, L.; Maes, A.; Elgorashi, E.E.; van Staden, J.; De Kimpe, N. Genotoxicity of melanoidin fractions derived from a standard glucose/glycine model. J. Agric. Food Chem. 2004, 52, 318–323. [Google Scholar] [CrossRef]
- Delgado-Andrade, C. Maillard reaction products: Some considerations on their health effects. Clin. Chem. Lab. Med. 2014, 52, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Moreira, A.S.P.; Nunes, F.M.; Domingues, M.R.; Coimbra, M.A. Coffee melanoidins: Structures, mechanisms of formation and potential health impacts. Food Funct. 2012, 3, 903–915. [Google Scholar] [CrossRef] [PubMed]
- Faist, V.; Erbersdobler, H.F. Metabolic transit and in vivo effects of melanoidins and precursor compounds deriving from the Maillard reaction. Ann. Nutr. Metab. 2001, 45, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Fogliano, V.; Morales, F.J. Estimation of dietary intake of melanoidins from coffee and bread. Food Funct. 2011, 2, 117–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helou, C.; Denis, S.; Spatz, M.; Marier, D.; Rame, V.; Alric, M.; Tessier, F.J.; Gadonna-Widehem, P. Insights into bread melanoidins: Fate in the upper digestive tract and impact on the gut microbiota using in vitro systems. Food Funct. 2015, 6, 3737–3745. [Google Scholar] [CrossRef]
- Helou, C.; Jacolot, P.; Niquet-Léridon, C.; Gadonna-Widehem, P.; Tessier, F.J. Maillard reaction products in bread: A novel semi-quantitative method for evaluating melanoidins in bread. Food Chem. 2016, 190, 904–911. [Google Scholar] [CrossRef]
- Helou, C.; Anton, P.M.; Niquet-Léridon, C.; Spatz, M.; Tessier, F.J.; Gadonna-Widehem, P. Fecal excretion of Maillard reaction products and the gut microbiota composition of rats fed with bread crust or bread crumb. Food Funct. 2017, 8, 2722–2730. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Bellesia, A. The gastro-intestinal tract as the major site of biological action of dietary melanoidins. Amino Acids 2015, 47, 1077–1089. [Google Scholar] [CrossRef] [Green Version]
- Rufián-Henares, J.A.; Morales, F.J. A new application of a commercial microtiter plate-based assay for assessing the antimicrobial activity of Maillard reaction products. Food Res. Int. 2006, 39, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Rufian-Henares, J.A.; Morales, F.J. Antimicrobial activity of melanoidins. J. Food Qual. 2007, 30, 160–168. [Google Scholar] [CrossRef] [Green Version]
- Einarsson, H.; Snygg, B.G.; Eriksson, C. Inhibition of bacterial growth by Maillard reaction products. J. Agric. Food Chem. 1983, 31, 1043–1047. [Google Scholar] [CrossRef]
- Hiramoto, S.; Itoh, K.; Shizuuchi, S.; Kawachi, Y.; Morishita, Y.; Nagase, M.; Suzuki, Y.; Nobuta, Y.; Sudou, Y.; Nakamura, O. Melanoidin, a food protein-derived advanced Maillard reaction product, suppresses Helicobacter pylori in vitro and in vivo. Helicobacter 2004, 9, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Rufián-Henares, J.A.; Morales, F.J. Antimicrobial activity of melanoidins against Escherichia coli is mediated by a membrane-damage mechanism. J. Agric. Food Chem. 2008, 56, 2357–2362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casterline, J.L.; Oles, C.J.; Ku, Y. In vitro fermentation of various food fiber fractions. J. Agric. Food Chem. 1997, 45, 2463–2467. [Google Scholar] [CrossRef]
- Zhong, Y.; Nyman, M.; Fåk, F. Modulation of gut microbiota in rats fed high-fat diets by processing whole-grain barley to barley malt. Mol. Nutr. Food Res. 2015, 59, 2066–2076. [Google Scholar] [CrossRef] [PubMed]
- Tuohy, K.M.; Hinton, D.J.S.; Davies, S.J.; Crabbe, M.J.C.; Gibson, G.R.; Ames, J.M. Metabolism of Maillard reaction products by the human gut microbiota–implications for health. Mol. Nutr. Food Res. 2006, 50, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, R.C.; Fogliano, V. Bread crust melanoidins as potential prebiotic ingredients. Mol. Nutr. Food Res. 2005, 49, 673–678. [Google Scholar] [CrossRef]
- Ames, J.M.; Wynne, A.; Hofmann, A.; Plos, S.; Gibson, G.R. The effect of a model melanoidin mixture on faecal bacterial populations in vitro. Br. J. Nutr. 1999, 82, 489–495. [Google Scholar] [CrossRef] [Green Version]
- Reichardt, N.; Gniechwitz, D.; Steinhart, H.; Bunzel, M.; Blaut, M. Characterization of high molecular weight coffee fractions and their fermentation by human intestinal microbiota. Mol. Nutr. Food Res. 2009, 53, 287–299. [Google Scholar] [CrossRef]
- Rodhouse, L.; Carbonero, F. Overview of craft brewing specificities and potentially associated microbiota. Crit. Rev. Food Sci. Nutr. 2019, 59, 462–473. [Google Scholar] [CrossRef]
- Carvalho, D.O.; Correia, E.; Lopes, L.; Guido, L.F. Further insights into the role of melanoidins on the antioxidant potential of barley malt. Food Chem. 2014, 160, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Tangerman, A.; Nagengast, F.M. A gas chromatographic analysis of fecal short-chain fatty acids, using the direct injection method. Anal. Biochem. 1996, 236, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoetendal, E.G.; Heilig, H.G.H.J.; Klaassens, E.S.; Booijink, C.C.G.M.; Kleerebezem, M.; Smidt, H.; De Vos, W.M. Isolation of DNA from bacterial samples of the human gastrointestinal tract. Nat. Protoc. 2006, 1, 870. [Google Scholar] [CrossRef] [PubMed]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.; Harper, D.; Ryan, R. Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Morales, F.J.; Somoza, V.; Fogliano, V. Physiological relevance of dietary melanoidins. Amino Acids 2012, 42, 1097–1109. [Google Scholar] [CrossRef]
- Finot, P.A.; Magnenat, E. Metabolic transit of early and advanced Maillard products. Prog. Food Nutr. Sci. 1981, 5, 193–207. [Google Scholar]
- Vitaglione, P.; Morisco, F.; Mazzone, G.; Amoruso, D.C.; Ribecco, M.T.; Romano, A.; Fogliano, V.; Caporaso, N.; D’Argenio, G. Coffee reduces liver damage in a rat model of steatohepatitis: The underlying mechanisms and the role of polyphenols and melanoidins. Hepatology 2010, 52, 1652–1661. [Google Scholar] [CrossRef]
- Kanauchi, O.; Oshima, T.; Andoh, A.; Shioya, M.; Mitsuyama, K. Germinated barley foodstuff ameliorates inflammation in mice with colitis through modulation of mucosal immune system. Scand. J. Gastroenterol. 2008, 43, 1346–1352. [Google Scholar] [CrossRef]
- Delgado-Andrade, C.; de la Cueva, S.P.; Peinado, M.J.; Rufián-Henares, J.Á.; Navarro, M.P.; Rubio, L.A. Modifications in bacterial groups and short chain fatty acid production in the gut of healthy adult rats after long-term consumption of dietary Maillard reaction products. Food Res. Int. 2017, 100, 134–142. [Google Scholar] [CrossRef] [PubMed]
- De Souza, A.Z.Z.; Zambom, A.Z.; Abboud, K.Y.; Reis, S.K.; Tannihão, F.; Guadagnini, D.; Saad, M.J.A.; Prada, P.O. Oral supplementation with L-glutamine alters gut microbiota of obese and overweight adults: A pilot study. Nutrition 2015, 31, 884–889. [Google Scholar] [CrossRef] [PubMed]
- Martínez, I.; Lattimer, J.M.; Hubach, K.L.; Case, J.A.; Yang, J.; Weber, C.G.; Louk, J.A.; Rose, D.J.; Kyureghian, G.; Peterson, D.A. Gut microbiome composition is linked to whole grain-induced immunological improvements. ISME J. 2013, 7, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Mu, C.; He, X.; Su, Y.; Mao, S.; Zhang, J.; Smidt, H.; Zhu, W. Effects of dietary fibre source on microbiota composition in the large intestine of suckling piglets. FEMS Microbiol. Lett. 2016, 363. [Google Scholar] [CrossRef]
- Salyers, A.A.; Vercellotti, J.R.; West, S.E.; Wilkins, T.D. Fermentation of mucin and plant polysaccharides by strains of Bacteroides from the human colon. Appl. Environ. Microbiol. 1977, 33, 319–322. [Google Scholar] [CrossRef] [Green Version]
- Ubeda, C.; Bucci, V.; Caballero, S.; Djukovic, A.; Toussaint, N.C.; Equinda, M.; Lipuma, L.; Ling, L.; Gobourne, A.; No, D. Intestinal microbiota containing Barnesiella species cures vancomycin-resistant Enterococcus faecium colonization. Infect. Immun. 2013, 81, 965–973. [Google Scholar] [CrossRef] [Green Version]
- Wylie, K.M.; Truty, R.M.; Sharpton, T.J.; Mihindukulasuriya, K.A.; Zhou, Y.; Gao, H.; Sodergren, E.; Weinstock, G.M.; Pollard, K.S. Novel bacterial taxa in the human microbiome. PLoS ONE 2012, 7, e35294. [Google Scholar] [CrossRef]
- Al, K.; Sarr, O.; Dunlop, K.; Gloor, G.B.; Reid, G.; Burton, J.; Regnault, T.R.H. Impact of birth weight and postnatal diet on the gut microbiota of young adult guinea pigs. PeerJ 2017, 5, e2840. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Le Leu, R.K.; Christophersen, C.T.; Somashekar, R.; Conlon, M.A.; Meng, X.Q.; Winter, J.M.; Woodman, R.J.; McKinnon, R.; Young, G.P. Manipulation of the gut microbiota using resistant starch is associated with protection against colitis-associated colorectal cancer in rats. Carcinogenesis 2016, 37, 366–375. [Google Scholar] [CrossRef]
- Wang, L.; Hu, L.; Yan, S.; Jiang, T.; Fang, S.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Effects of different oligosaccharides at various dosages on the composition of gut microbiota and short-chain fatty acids in mice with constipation. Food Funct. 2017, 8, 1966–1978. [Google Scholar] [CrossRef]
- Tailford, L.E.; Crost, E.H.; Kavanaugh, D.; Juge, N. Mucin glycan foraging in the human gut microbiome. Front. Genet. 2015, 6, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, Q.; Sun, W.; Shan, X.; Jiang, H.; Cai, C.; Hao, J.; Li, G.; Yu, G. Carrageenan-induced colitis is associated with decreased population of anti-inflammatory bacterium, Akkermansia muciniphila, in the gut microbiota of C57BL/6J mice. Toxicol. Lett. 2017, 279, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, R.C.; Esposito, F.; Napolitano, A.; Ritieni, A.; Fogliano, V. Characterization of a new potential functional ingredient: Coffee silverskin. J. Agric. Food Chem. 2004, 52, 1338–1343. [Google Scholar] [CrossRef] [PubMed]
- Jaquet, M.; Rochat, I.; Moulin, J.; Cavin, C.; Bibiloni, R. Impact of coffee consumption on the gut microbiota: A human volunteer study. Int. J. Food Microbiol. 2009, 130, 117–121. [Google Scholar] [CrossRef]
- Noble, E.E.; Hsu, T.M.; Jones, R.B.; Fodor, A.A.; Goran, M.I.; Kanoski, S.E. Early-life sugar consumption affects the rat microbiome independently of obesity. J. Nutr. 2016, 147, 20–28. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feed Intake by Days (g/day) | Estimated Mean |
|---|---|
| 4 days | 0.53 ± 0.01 b |
| 7 days | 0.74 ± 0.01 a |
| 11 days | 0.46 ± 0.01 cd |
| 14 days | 0.45 ± 0.01 d |
| 18 days | 0.49 ± 0.01 c |
| 21 days | 0.33 ± 0.01 f |
| 25 days | 0.38 ± 0.01 e |
| Average Daily Gain (g/day) | 7 Days | 18 Days | 25 Days |
|---|---|---|---|
| Melanoidin-free | 0.17 ± 0.07 abcA | 0.19 ± 0.07 aA | 0.31 ± 0.07 abA |
| 25% Melanoidins | 0.048 ± 0.07 cB | 0.34 ± 0.07 aA | 0.47 ± 0.07 aA |
| 50% Melanoidins | 0.28 ± 0.07 abA | 0.18 ± 0.07 aA | 0.18 ± 0.07 bA |
| 75% Melanoidins | 0.34 ± 0.07 aA | 0.20 ± 0.07 aAB | 0.14 ± 0.07 bB |
| 100% Melanoidins | 0.13 ± 0.07 bcA | 0.19 ± 0.07 aA | 0.23 ± 0.07 bA |
| Melanoidins | Day 0 | Day 3 | Day 7 | Day 14 | Day 21 | |
|---|---|---|---|---|---|---|
| Acetate (mmol/mL) | 0% | 4.01 ± 0.68 a | 3.1 ± 0.94 aA | 4.1 ± 1.33 aA | 2.35 ± 0.97 aA | 2.13 ± 0.63 aB |
| 100% | 4.01 ± 0.68 a | 3.7 ± 1.33 aA | 3.6 ± 1.41 aA | 4.14 ± 1.05 aA | 4.51 ± 0.43 aA | |
| Propionate (mmol/mL) | 0% | 0.25 ± 0.03 a | 0.21 ± 0.03 aA | 0.16 ± 0.04 aA | 0.07 ± 0.03 bA | 0.06 ± 0.01 bB |
| 100% | 0.25 ± 0.03 a | 0.14 ± 0.05 aA | 0.11 ± 0.04 bA | 0.09 ± 0.02 bA | 0.09 ± 0.03 bA | |
| Butyrate (mmol/mL) | 0% | 0.42 ± 0.17 a | 0.27 ± 0.04 aA | 0.18 ± 0.07 aA | 0.06 ± 0.03 bB | 0.10 ± 0.01 bA |
| 100% | 0.42 ± 0.17 a | 0.13 ± 0.04 bA | 0.09 ± 0.02 bA | 0.14 ± 0.01 bA | 0.08 ± 0.02 bA | |
| Total (mmol/mL) | 0% | 4.7 ± 0.88 | 3.6 ± 1.01 | 4.44 ± 1.44 | 2.5 ± 1.03 | 2.29 ± 0.62 |
| 100% | 4.7 ± 0.88 | 3.8 ± 1.42 | 3.8 ± 1.5 | 4.4 ± 1.08 | 4.68 ± 0.48 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aljahdali, N.; Gadonna-Widehem, P.; Anton, P.M.; Carbonero, F. Gut Microbiota Modulation by Dietary Barley Malt Melanoidins. Nutrients 2020, 12, 241. https://doi.org/10.3390/nu12010241
Aljahdali N, Gadonna-Widehem P, Anton PM, Carbonero F. Gut Microbiota Modulation by Dietary Barley Malt Melanoidins. Nutrients. 2020; 12(1):241. https://doi.org/10.3390/nu12010241
Chicago/Turabian StyleAljahdali, Nesreen, Pascale Gadonna-Widehem, Pauline M. Anton, and Franck Carbonero. 2020. "Gut Microbiota Modulation by Dietary Barley Malt Melanoidins" Nutrients 12, no. 1: 241. https://doi.org/10.3390/nu12010241
APA StyleAljahdali, N., Gadonna-Widehem, P., Anton, P. M., & Carbonero, F. (2020). Gut Microbiota Modulation by Dietary Barley Malt Melanoidins. Nutrients, 12(1), 241. https://doi.org/10.3390/nu12010241