Urine Untargeted Metabolomic Profiling Is Associated with the Dietary Pattern of Successful Aging among Malaysian Elderly

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Study Design
2.2. Criteria of Successful Aging
2.3. Identification of Dietary Pattern for Successful Aging
2.4. Sample Preparation
2.5. NMR Acquisition
2.6. Urinary Metabolites Identification
2.7. Statistical Analysis
2.8. Pathway Analysis
3. Results
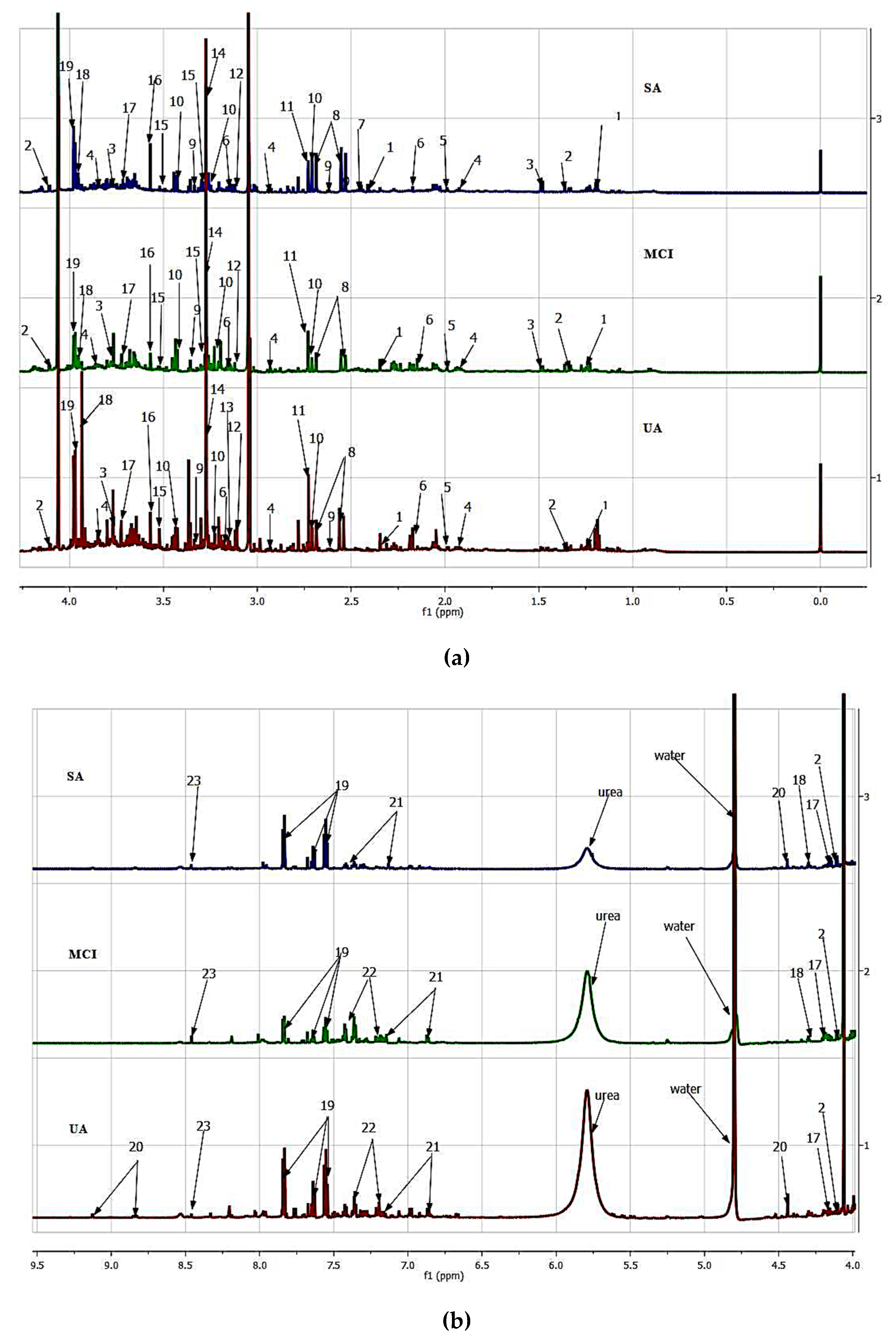
3.1. Urinary Metabolites Identification
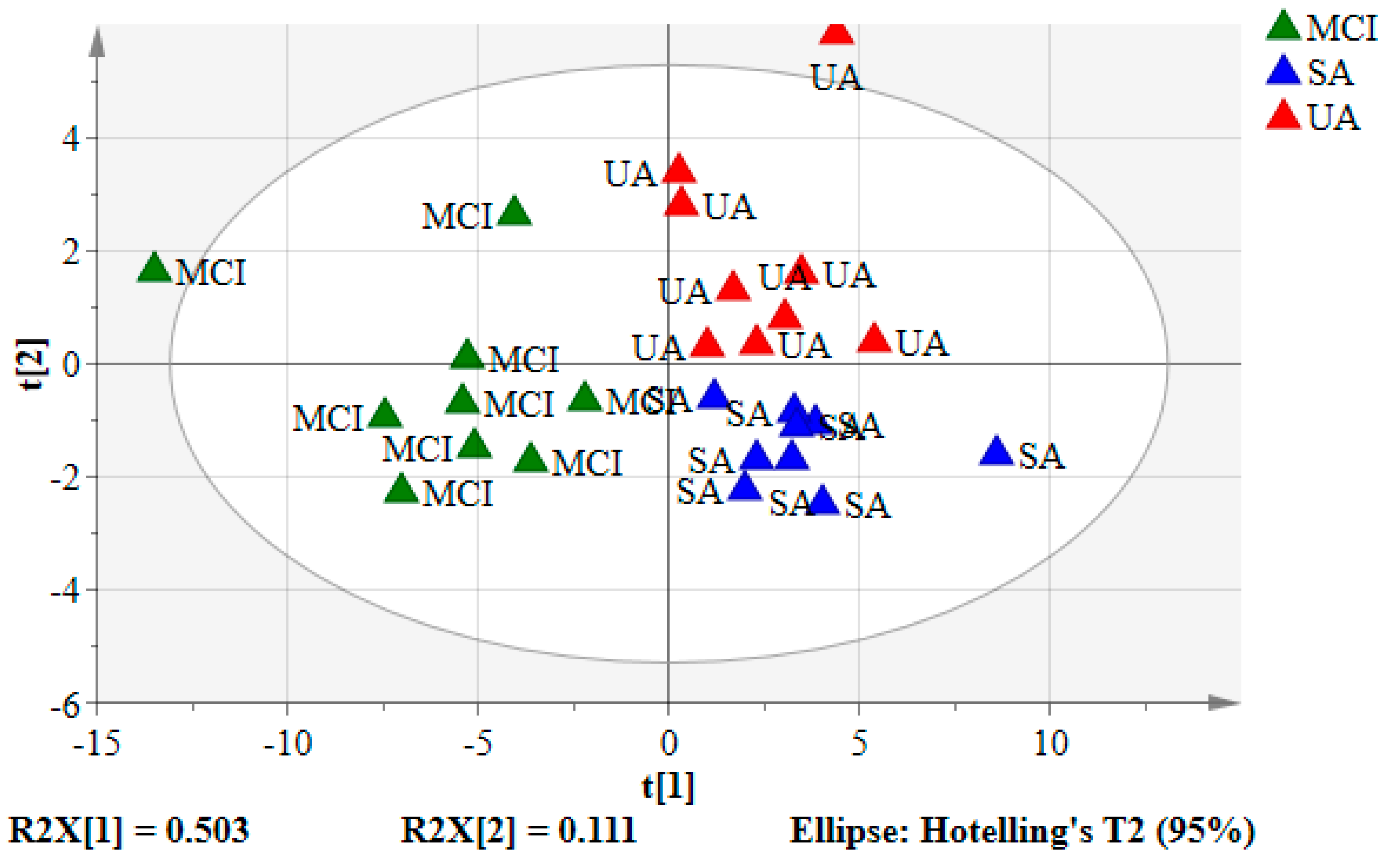
3.2. Principal Component Analysis (PCA) of the Urine Samples
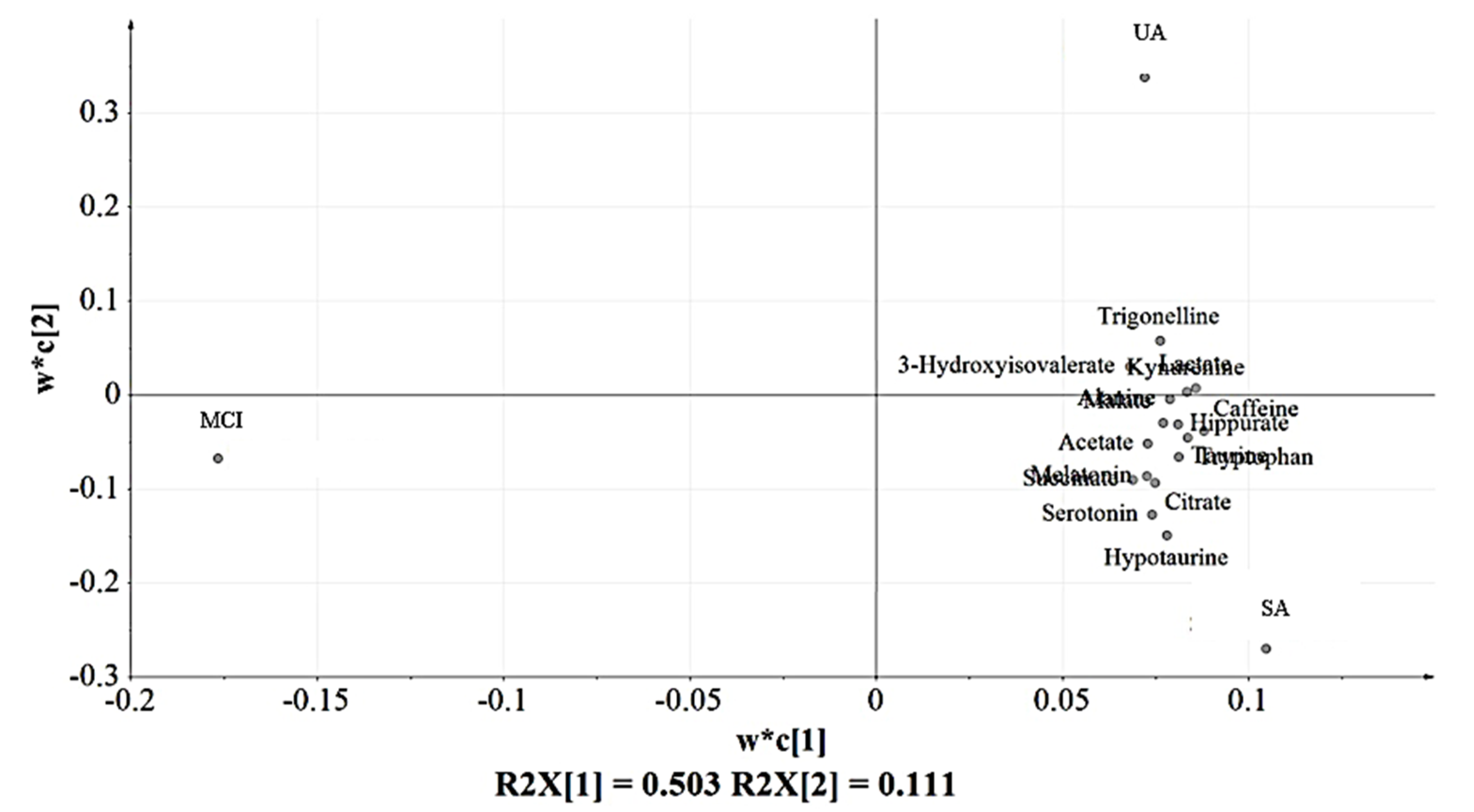
3.3. Partial Least Squares-Discriminant Analysis (PLS-DA) of the Urine Samples
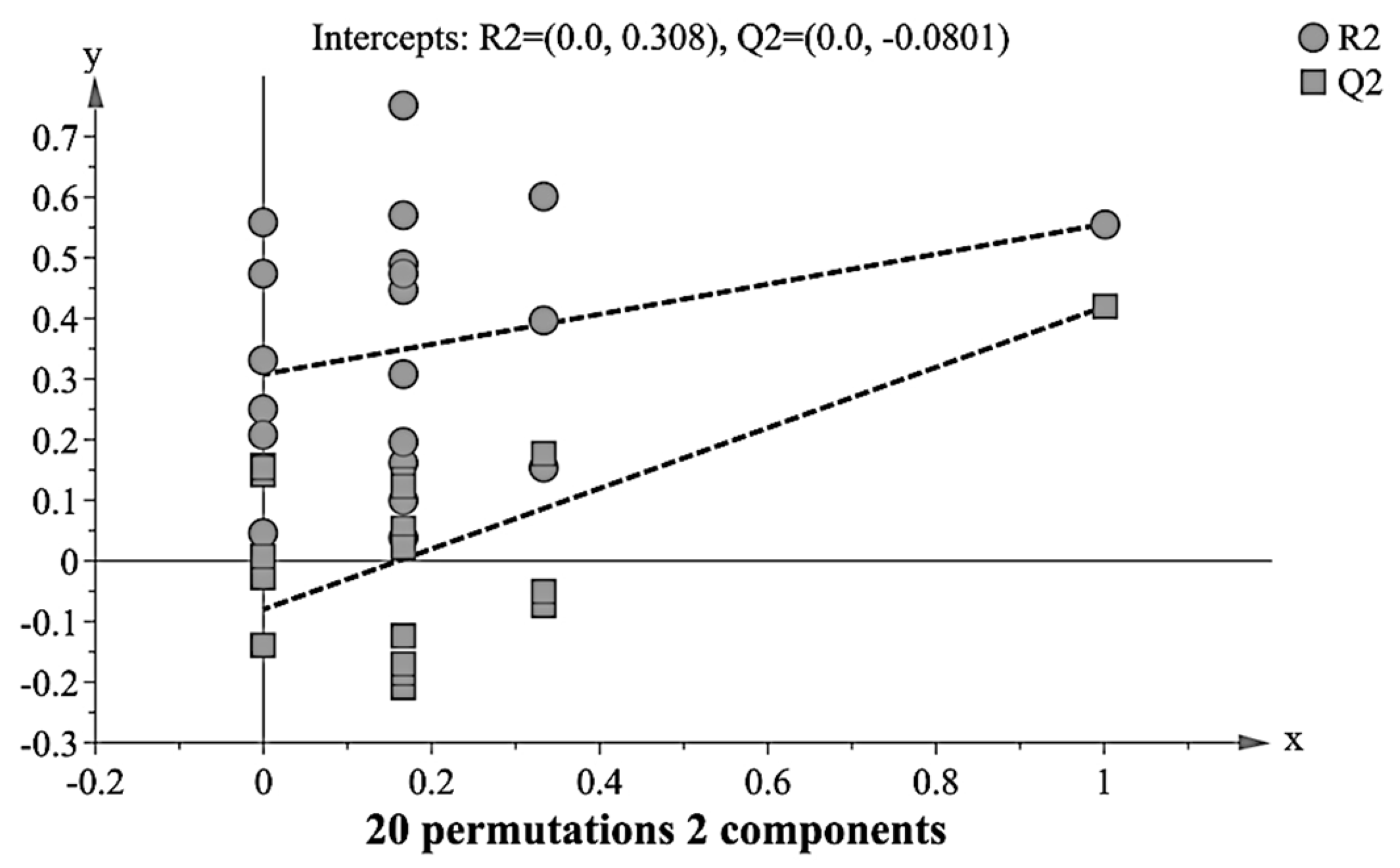
3.4. Validation of PLS-DA Model
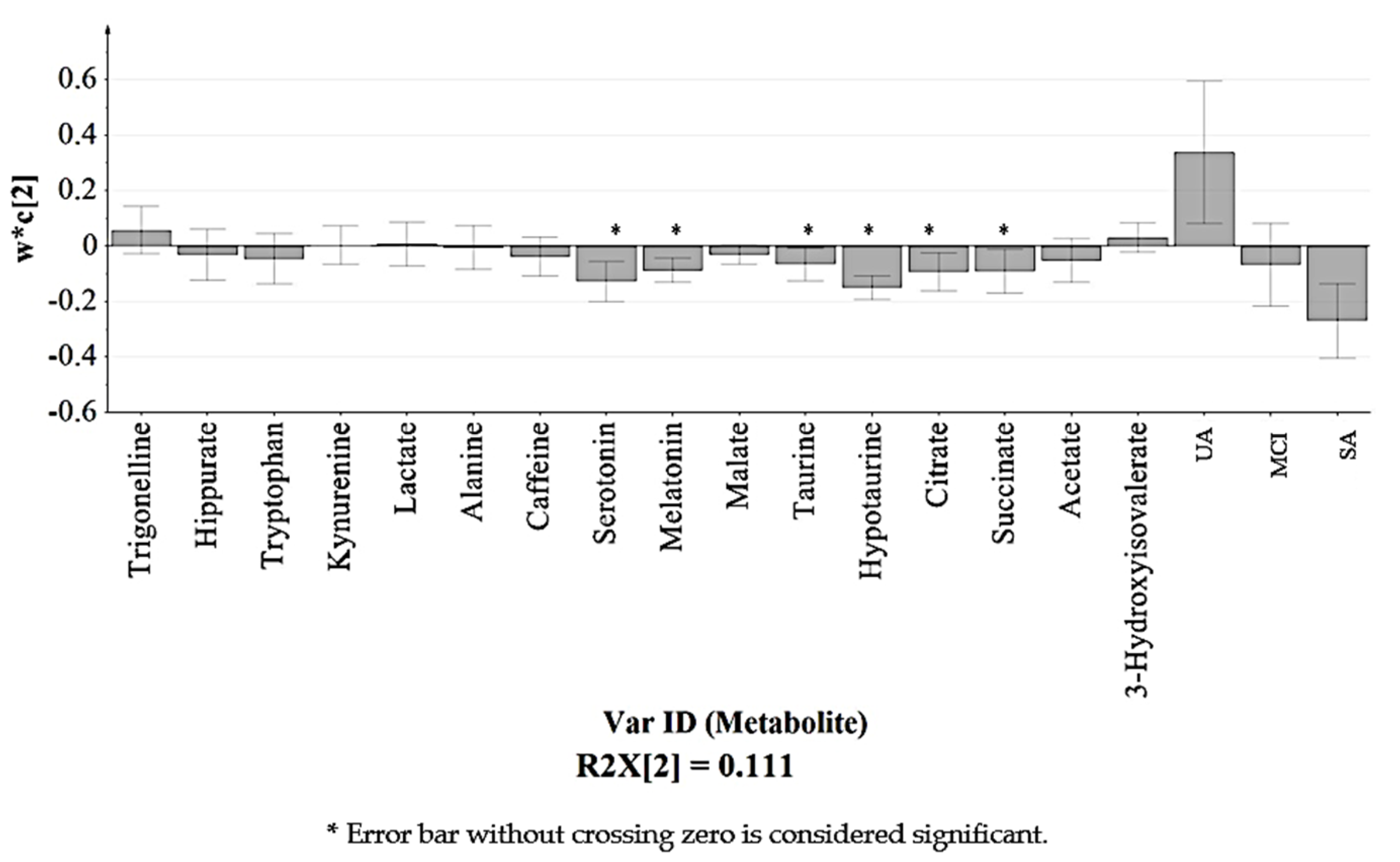
3.5. Relative Quantification of Identified Urinary Metabolites
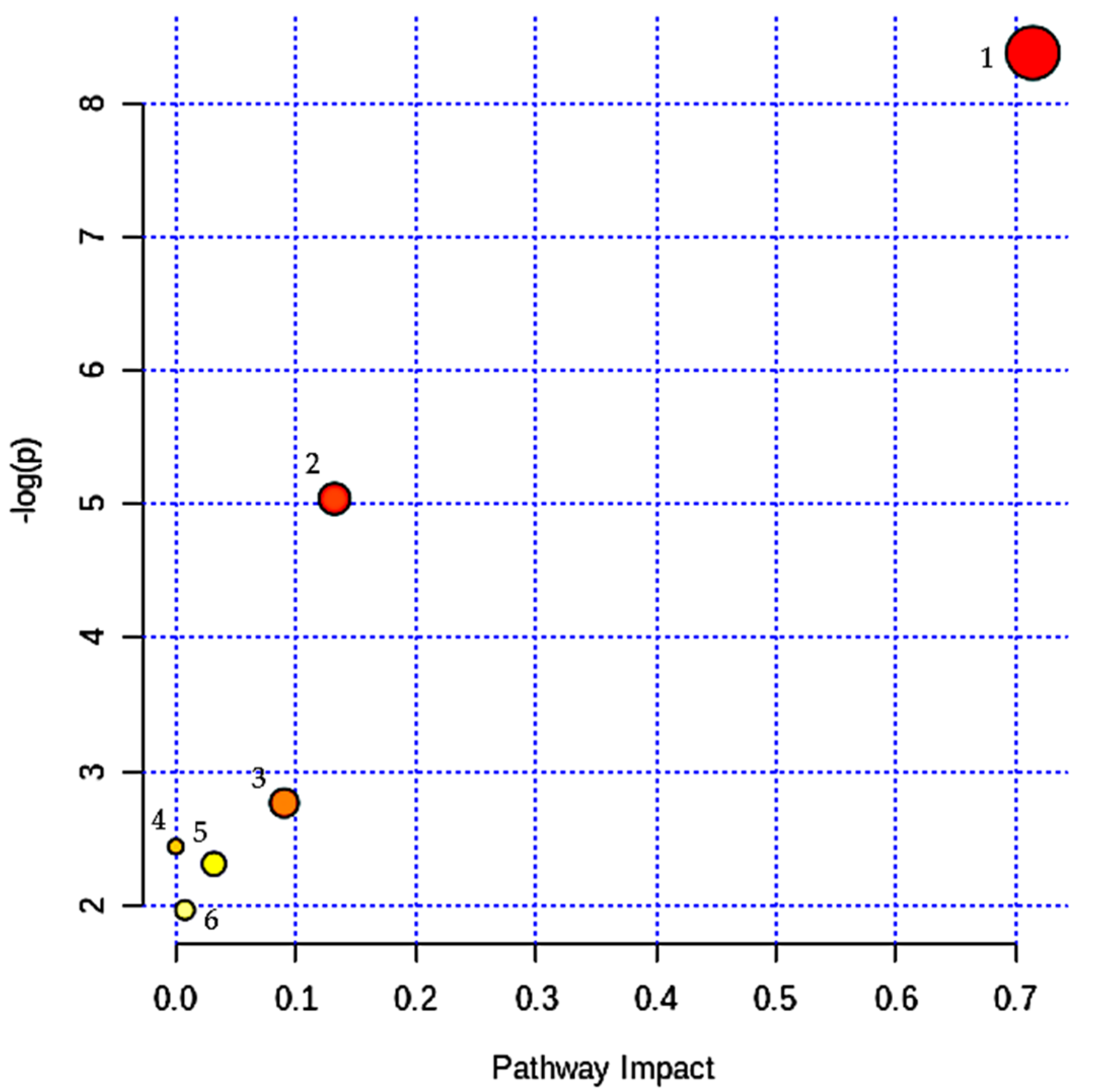
3.6. Probable Metabolic Pathways for Successful Aging (SA)
3.7. Relationship between Dietary Pattern and SA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Trushina, E.; Dutta, T.; Persson, X.M.T.; Mielke, M.; Petersen, R.C. Identification of Altered Metabolic Pathways in Plasma and CSF in Mild Cognitive Impairment and Alzheimer’s Disease Using Metabolomics. PLoS ONE 2013, 8, e63644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pamplona, R.; Barja, G. An evolutionary comparative scan for longevity-related oxidative stress resistance mechanisms in homeotherms. Biogerontology 2011, 12, 409–435. [Google Scholar] [CrossRef] [PubMed]
- Lau, H.; Ludin, A.F.M.; Rajab, N.F.; Shahar, S. Identification of Neuroprotective Factors Associated with Successful Ageing and Risk of Cognitive Impairment among Malaysia Older Adults. Curr. Gerontol. Geriatr. Res. 2017, 2017, 4218756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badrasawi, M.; Shahar, S.; Rajab, N.F.; Singh, D.K.A. Blood Inflammatory Markers, Anabolic Hormone, Vitamin D and L-Carnitine Status According to Frailty Status among Older Adults. Sains Malays. 2019, 48, 613–619. [Google Scholar] [CrossRef]
- Graham, S.F.; Chevallier, O.P.; Elliott, C.T.; Hölscher, C.; Johnston, J.; McGuinness, B.; Kehoe, P.G.; Passmore, A.P.; Green, B.D. Untargeted Metabolomic Analysis of Human Plasma Indicates Differentially Affected Polyamine and L-Arginine Metabolism in Mild Cognitive Impairment Subjects Converting to Alzheimer’s Disease. PLoS ONE 2015, 10, e0119452. [Google Scholar] [CrossRef]
- Zheng, J.; Dixon, R.A.; Li, L. Development of Isotope Labeling LC–MS for Human Salivary Metabolomics and Application to Profiling Metabolome Changes Associated with Mild Cognitive Impairment. Anal. Chem. 2012, 84, 10802–10811. [Google Scholar] [CrossRef] [Green Version]
- Figueira, J.; Jönsson, P.; Adolfsson, R.; Adolfsson, R.; Nyberg, L.; Öhman, A. NMR analysis of the human saliva metabolome distinguishes dementia patients from matched controls. Mol. BioSyst. 2016, 12, 2562–2571. [Google Scholar] [CrossRef] [Green Version]
- Tukianen, T.; Tynkkynen, T.; Mäkinen, V.P.; Jylänki, P.; Kangas, A.J.; Hokkanen, J.; Vehtari, A.; Gröhn, O.; Hallikainen, M.; Soininen, H.; et al. A multi-metabolite analysis of serum by 1H NMR spectroscopy: Early systemic signs of Alzheimer’s disease. Biochem. Biophys. Res. Commun. 2008, 375, 356–361. [Google Scholar] [CrossRef]
- Wang, G.; Zhou, Y.; Huang, F.J.; Tang, H.D.; Xu, X.H.; Liu, J.J.; Wang, Y.; Deng, Y.L.; Ren, R.J.; Xu, W.; et al. Plasma Metabolite Profiles of Alzheimer’s Disease and Mild Cognitive Impairment. J. Proteome Res. 2014, 13, 2649–2658. [Google Scholar] [CrossRef]
- Johnson, L.C.; Parker, K.; Aguirre, B.F.; Nemkov, T.G.; D’Alessandro, A.; Johnson, S.A.; Seals, D.R.; Martens, C.R. The plasma metabolome as a predictor of biological aging in humans. GeroScience 2019, 41, 895–906. [Google Scholar] [CrossRef]
- Johnson, L.C.; Martens, C.R.; Santos-Parker, J.R.; Bassett, C.J.; Strahler, T.R.; Cruickshank-Quinn, C.; Reisdorph, N.; McQueen, M.B.; Seals, D.R. Amino acid and lipid associated plasma metabolomic patterns are related to healthspan indicators with aging in human. Clin. Sci. 2018, 132, 1765–1777. [Google Scholar] [CrossRef]
- Robinson, O.; Hyam, M.C.; Karaman, I.; Pinto, R.C.; Ala-Korpela, M.; Handakas, E.; Fiorito, G.; Gao, H.; Heard, A.; Järvelin, M.R.; et al. Determinants of accelerated metabolomic and epigenetic aging in a UK cohort. Aging Cell 2020, 19, e13149. [Google Scholar] [CrossRef]
- Fakhruddin, N.N.I.N.M.; Shahar, S.; Rajikan, R.; Omar, M.A.; Din, N.C.; Razali, R.; Harith, S.; Mohamed, H.J.J.; Hakim, B.N.A. Identification of Dietary Patterns associated with Characteristics of Successful Aging. Malays. J. Nutr. 2019, 25, 47–57. [Google Scholar] [CrossRef]
- Hodge, A.M.; O’Dea, K.; English, D.R.; Giles, G.; Flicker, L. Dietary patterns as predictors of successful ageing. J. Nutr. Health Aging 2013, 18, 221–227. [Google Scholar] [CrossRef]
- Koulman, A.; Volmer, D. Perspectives for Metabolomics in Human Nutrition: An Overview. Nutr. Bull. 2008, 33, 324–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, M.B.S.; Rinnan, Å.; Manach, C.; Poulsen, S.K.; Pujos-Guillot, E.; Larsen, T.M.; Astrup, A.; Dragsted, L.O. Untargeted Metabolomics as a Screening Tool for Estimating Compliance to a Dietary Pattern. J. Proteome Res. 2014, 13, 1405–1418. [Google Scholar] [CrossRef] [PubMed]
- Konstantinova, S.V.; Tell, G.S.; Vollset, S.E.; Ulvik, A.; Drevon, C.A.; Ueland, P.M. Dietary patterns, food groups, and nutrients as predictors of plasma choline and betaine in middle-aged and elderly men and women. Am. J. Clin. Nutr. 2008, 88, 1663–1669. [Google Scholar] [CrossRef]
- O’Sullivan, A.; Gibney, M.J.; Brennan, L. Dietary intake patterns are reflected in metabolomic profiles: Potential role in dietary assessment studies. Am. J. Clin. Nutr. 2011, 93, 314–321. [Google Scholar] [CrossRef] [Green Version]
- Shahar, S.; Omar, A.; Vanoh, D.; Hamid, T.A.; Mukari, S.Z.M.S.; Din, N.C.; Rajab, N.F.; Mohammed, Z.; Ibrahim, R.; Loo, W.H.; et al. Approaches in methodology for population-based longitudinal study on neuroprotective model for healthy longevity (TUA) among Malaysian Older Adults. Aging Clin. Exp. Res. 2015, 28, 1089–1104. [Google Scholar] [CrossRef]
- Fakhruddin, N.N.I.; Shahar, S.; Aziz, N.A.; Yahya, H.M.; Rajikan, R. Which aging group prone to has inadequate nutrient intake? LRGS TUA Study. Sains Malays. 2015, 45, 1381–1391. [Google Scholar]
- Razali, R.; Jean-Li, L.; Jaffar, A.; Ahmad, M.; Shah, S.A.; Ibrahim, N.; Din, N.C.; Jaafar, N.R.N.; Midin, M.; Sidi, H.; et al. Is the Bahasa Malaysia version of the Montreal Cognitive Assessment (MoCA-BM) a better instrument than the Malay version of the Mini Mental State Examination (M-MMSE) in screening for mild cognitive impairment (MCI) in the elderly? Compr. Psychiatry 2014, 55, S70–S75. [Google Scholar] [CrossRef]
- Wechsler, D. The Measurement and Appraisal of Adult Intelligence, 4th ed.; Williams & Wilkins Co.: Baltimore, MD, USA, 1958. [Google Scholar]
- Schmidt, M. Rey Auditory Verbal Learning Test: RAVLT: A Handbook; Western Psychological Services: Los Angeles, CA, USA, 1996. [Google Scholar]
- Katz, S.; Grotz, R.C.; Downs, T.D.; Cash, H.R. Progress in Development of the Index of ADL. Gerontologist 1970, 10, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Graf, C. The Lawton Instrumental Activities of Daily Living Scale. Am. J. Nurs. 2008, 108, 52–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, L.; Sun, W. Human urine proteome: A powerful source for clinical research. In Urine Proteomics in Kidney Disease Biomarker Discovery; Springer: Dordrecht, The Netherland, 2015; Volume 845, pp. 31–41. [Google Scholar]
- Beckonert, O.; Keun, H.C.; Ebbels, T.M.D.; Bundy, J.G.; Holmes, E.; Lindon, J.C.; Nicholson, J.K. Metabolic profiling, metabolomic and metabonomic procedures for NMR spectroscopy of urine, plasma, serum and tissue extracts. Nat. Protoc. 2007, 2, 2692–2703. [Google Scholar] [CrossRef] [PubMed]
- Keun, H.C.; Ebbels, T.M.D.; Antti, H.; Bollard, M.E.; Beckonert, O.; Schlotterbeck, G.; Senn, H.; Niederhauser, U.; Holmes, E.; Lindon, J.C.; et al. Analytical Reproducibility in1H NMR-Based Metabonomic Urinalysis. Chem. Res. Toxicol. 2002, 15, 1380–1386. [Google Scholar] [CrossRef]
- Salek, R.M.; Steinbeck, C.; Viant, M.R.; Goodacre, R.; Dunn, W.B. The role of reporting standards for metabolite annotation and identification in metabolomic studies. GigaScience 2013, 2, 13. [Google Scholar] [CrossRef] [PubMed]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.D.; Daykin, C.A.; Fan, T.W.M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Lindon, J.C.; Nicholson, J.K.; Everett, J.R. NMR Spectroscopy of Biofluids. In Annual Reports on NMR Spectroscopy; Academic Press: Cambridge, MA, USA, 1999; Volume 38, pp. 1–88. [Google Scholar]
- Tulipani, S.; Llorach, R.; Jáuregui, O.; López-Uriarte, P.J.; Garcia-Aloy, M.; Bulló, M.; Salas-Salvadó, J.; Andres-Lacueva, C. Metabolomics Unveils Urinary Changes in Subjects with Metabolic Syndrome following 12-Week Nut Consumption. J. Proteome Res. 2011, 10, 5047–5058. [Google Scholar] [CrossRef]
- Wang, X.J.; Zhang, A.; Han, Y.; Wang, P.; Sun, H.; Song, G.; Dong, T.; Yuan, Y.; Yuan, X.; Zhang, M.; et al. Urine metabolomics analysis for biomarker discovery and detection of jaundice syndrome in patients with liver disease. Mol. Cell. Proteom. 2012, 11, 370–380. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, L.; Trygg, J.; Wold, S. CV-ANOVA for significance testing of PLS and OPLS® models. J. Chemom. 2008, 22, 594–600. [Google Scholar] [CrossRef]
- Verwaest, K.A.; Vu, T.N.; Laukens, K.; Clemens, L.E.; Nguyen, H.H.P.; Van Gasse, B.; Martins, J.C.; Van Der Linden, A.M.; Dommisse, R. 1H NMR based metabolomics of CSF and blood serum: A metabolic profile for a transgenic rat model of Huntington disease. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2011, 1812, 1371–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scalbert, A.; Brennan, L.; Manach, C.; Andres-Lacueva, C.; Dragsted, L.O.; Draper, J.; Rappaport, S.M.; Van Der Hooft, J.J.; Wishart, D.S. The food metabolome: A window over dietary exposure. Am. J. Clin. Nutr. 2014, 99, 1286–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolliffe, I.T. Principal Components in Regression Analysis. In Principal Component Analysis; Springer: Berlin/Heidelberg, Germany, 2002; pp. 167–198. [Google Scholar]
- Eriksson, L.; Johansson, E.; Kettaneh-Wold, N.; Trygg, J.; Wikström, C.; Wold, S. Multi- and Megavariate Data Analysis; Umetrics: Umea, Sweden, 2005; Volume 1, pp. 263–336. [Google Scholar]
- Van Der Lee, S.J.; Teunissen, C.E.; Pool, R.; Shipley, M.J.; Teumer, A.; Chouraki, V.; Van Lent, D.M.; Tynkkynen, J.; Fischer, K.; Hernesniemi, J.; et al. Circulating metabolites and general cognitive ability and dementia: Evidence from 11 cohort studies. Alzheimer’s Dement. 2018, 14, 707–722. [Google Scholar] [CrossRef] [PubMed]
- Tynkkynen, J.; Chouraki, V.; Van Der Lee, S.J.; Hernesniemi, J.; Yang, Q.; Li, S.; Beiser, A.; Larson, M.G.; Sääksjärvi, K.; Shipley, M.J.; et al. Association of branched-chain amino acids and other circulating metabolites with risk of incident dementia and Alzheimer’s disease: A prospective study in eight cohorts. Alzheimer’s Dement. 2018, 14, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Ezza, H.A.S.; Khadrawyb, Y.A. Glutamate excitotoxicity and neurodegeneration. J. Mol. Genet. Med. 2014, 8, 141. [Google Scholar]
- Wu, J.Y.; Prentice, H. Role of taurine in the central nervous system. J. Biomed. Sci. 2010, 17, S1–S6. [Google Scholar] [CrossRef] [Green Version]
- Mapstone, M.; Cheema, A.K.; Fiandaca, M.; Zhong, X.; Mhyre, T.R.; MacArthur, L.H.; Hall, W.J.; Fisher, S.G.; Peterson, D.R.; Haley, J.M.; et al. Plasma phospholipids identify antecedent memory impairment in older adults. Nat. Med. 2014, 20, 415–418. [Google Scholar] [CrossRef]
- Menzie, J.; Pan, C.; Prentice, H.; Wu, J.Y. Taurine and central nervous system disorders. Amino Acids 2012, 46, 31–46. [Google Scholar] [CrossRef]
- El Idrissi, A.; Trenkner, E. Taurine as a modulator of excitatory and inhibitory neurotransmission. Neurochem. Res. 2004, 29, 189–197. [Google Scholar] [CrossRef]
- Graham, S.F.; Hölscher, C.; Green, B.D. Metabolic signatures of human Alzheimer’s disease (AD): 1H NMR analysis of the polar metabolome of post-mortem brain tissue. Metabolomics 2014, 10, 744–753. [Google Scholar] [CrossRef]
- Chouraki, V.; Preis, S.R.; Yang, Q.; Beiser, A.; Li, S.; Larson, M.G.; Weinstein, G.; Wang, T.J.; Gerszten, R.E.; Vasan, R.S.; et al. Association of amine biomarkers with incident dementia and Alzheimer’s disease in the Framingham Study. Alzheimer’s Dement. 2017, 13, 1327–1336. [Google Scholar] [CrossRef] [PubMed]
- Zepf, F.D.; Sánchez, C.L.; Biskup, C.S.; Kötting, W.F.; Bubenzer, S.; Helmbold, K.; Eisert, A.; Gaber, T.J.; Landgraf, M.; Dahmen, B.; et al. Acute tryptophan depletion—Converging evidence for decreasing central nervous serotonin synthesis in rodents and humans. Acta Psychiatr. Scand. 2014, 129, 157–159. [Google Scholar] [CrossRef] [PubMed]
- Strasser, B.; Gostner, J.M.; Fuchs, D. Mood, food, and cognition: Role of tryptophan and serotonin. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Fukushige, H.; Fukuda, Y.; Tanaka, M.; Inami, K.; Wada, K.; Tsumura, Y.; Kondo, M.; Harada, T.; Wakamura, T.; Morita, T. Effects of tryptophan-rich breakfast and light exposure during the daytime on melatonin secretion at night. J. Physiol. Anthr. 2014, 33, 33. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Li, Y.; Li, S.; Zhou, Y.; Gan, R.Y.; Xu, D.P.; Li, H.B. Dietary Sources and Bioactivities of Melatonin. Nutrients 2017, 9, 367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sae-Teaw, M.; Johns, J.; Johns, N.P.; Subongkot, S. Serum melatonin levels and antioxidant capacities after consumption of pineapple, orange, or banana by healthy male volunteers. J. Pineal Res. 2013, 55, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Tsou, M.T. Association between Sleep Duration and Health Outcome in Elderly Taiwanese. Int. J. Gerontol. 2011, 5, 200–205. [Google Scholar] [CrossRef] [Green Version]
- Sokolenko, S.; McKay, R.T.; Blondeel, E.J.M.; Lewis, M.J.; Chang, D.; George, B.; Aucoin, M.G. Understanding the variability of compound quantification from targeted profiling metabolomics of 1D-1H-NMR spectra in synthetic mixtures and urine with additional insights on choice of pulse sequences and robotic sampling. Metabolomics 2013, 9, 887–903. [Google Scholar] [CrossRef]
- Roberts, L.D.; Souza, A.L.; Gerszten, R.E.; Clish, C.B. Targeted Metabolomics. Curr. Protoc. Mol. Boil. 2012, 98, 30–32. [Google Scholar] [CrossRef]
- Kohl, S.M.; Klein, M.S.; Hochrein, J.; Oefner, P.J.; Spang, R.; Gronwald, W. State-of-the art data normalization methods improve NMR-based metabolomic analysis. Metabolomics 2012, 8, 146–160. [Google Scholar] [CrossRef] [Green Version]
- Iida, M.; Harada, S.; Takebayashi, T. Application of Metabolomics to Epidemiological Studies of Atherosclerosis and Cardiovascular Disease. J. Atheroscler. Thromb. 2019, 26, 747–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Successful Cognitive Aging | Usual Aging | Mild Cognitive Impairment |
|---|---|---|
|
|
|
|
|
|
|
|
|
|
| |
|
| |
|
|
| Key | HMDB ID | Urinary Metabolites | Source | Chemical Shift (Multiplicity, J Value) |
|---|---|---|---|---|
| 1. | HMDB0000754 | 3-Hydroxyisovalerate | Endogenous and food | 1.26 (s), 2.36 (s) |
| 2. | HMDB0001311 | Lactate | Endogenous and food | 1.32 (d, 6.9), 4.11 (q, 6.9) |
| 3. | HMDB0000161 | Alanine | Endogenous and food | 1.44 (d, 7.1), 3.79 (q, 7.2) |
| 4. | HMDB0001389 | Melatonin | Endogenous and food | 1.91 (s), 2.93 (t, 6.8), 3.87 (s) |
| 5. | HMDB0000042 | Acetate | Endogenous and food | 1.93 (s) |
| 6. | HMDB0000201 | O-acetylcarnitine | Endogenous and food | 2.13 (s), 3.18 (s) |
| 7. | HMDB0000254 | Succinate | Endogenous and food | 2.40 (s) |
| 8. | HMDB0000094 | Citrate | Endogenous and food | 2.54 (d), 2.68 (d, 15.2) |
| 9. | HMDB0000965 | Hypotaurine | Endogenous | 2.60 (t, 6.9), 3.32 (t, 6.9) |
| 10. | HMDB0000251 | Taurine | Endogenous and food | 2.69 (s), 3.23 (t, 6.6), 3.42 (t, 6.6) |
| 11. | HMDB0000156 | Malate | Endogenous and food | 2.73 (dd, 15.4) |
| 12. | HMDB0000259 | Serotonin | Endogenous and food | 3.11 (t, 7.1), 7.41 (d, 8.7) |
| 13. | HMDB0004983 | Dimethyl sulfone | Endogenous and food | 3.14 (s) |
| 14. | HMDB0000925 | Trimethylamine-N-oxide | Endogenous | 3.26 (s) |
| 15. | HMDB0001964 | Caffein | Endogenous and food | 3.29 (s), 3.50 (s) |
| 16. | HMDB0000123 | Glycine | Endogenous and food | 3.55 (s) |
| 17. | HMDB0000684 | Kynurenine | Endogenous and food | 3.73 (d), 4.14 (t, 6.5, 4.2) |
| 18. | HMDB0000639 | Galactarate | Endogenous and food | 3.95 (s), 4.24 (s) |
| 19. | HMDB0000714 | Hippurate | Endogenous and food | 3.96 (d, 5.8), 7.54 (m), 7.62 (tt, 7.5, 1.5), 7.82 (dd, 8.4, 1.2) |
| 20. | HMDB0000875 | Trigonelline | Endogenous and food | 4.42 (s), 8.83 (m), 9.11 (s) |
| 21. | HMDB0000755 | 4-Hydroxyphenyllactate | Endogenous | 6.85 (d, 8.3), 7.15 (d, 8.2) |
| 22. | HMDB30396 | Tryptophan | Endogenous and food | 7.18 (d), 7.29 (s) |
| 23. | HMDB0000142 | Formate | Endogenous and food | 8.44 (s) |
| Chemical Shift (ppm) | Changes | |||
|---|---|---|---|---|
| SA vs. UA | SA vs. MCI | UA vs. MCI | ||
| 3-hydroxyisovalerate | 1.26 | − | + * | + * |
| Acetate | 1.93 | + | + * | + * |
| Malate | 2.73 | + | + * | + * |
| Alanine | 3.79 | − | + * | + * |
| Caffeine | 3.29 | + | + * | + * |
| Kynurenine | 4.14 | − | + * | + * |
| Lactate | 4.11 | − | + * | + * |
| Hippurate | 7.62 | + | + | + * |
| Tryptophan | 7.18 | + | + * | + * |
| Trigonelline | 8.81 | − | + * | + * |
| Succinate | 2.40 | + | + * | + * |
| Citrate | 2.54 | + * | + * | + |
| Hypotaurine | 2.60 | + * | + * | + * |
| Taurine | 2.69 | + * | + * | + * |
| Melatonin | 2.93 | + * | + * | + * |
| Serotonin | 3.11 | + * | + * | + |
| Urinary Metabolites | Chemical Shift (ppm) | Dietary Pattern Score | Oats | Tropical Fruits | |||
|---|---|---|---|---|---|---|---|
| r | p-Value | r | p-Value | r | p-Value | ||
| Citrate | 2.54 | 0.05 | 0.83 | 0.03 | 0.92 | 0.03 | 0.90 |
| Hypotaurine | 2.60 | 0.31 | 0.19 | 0.23 | 0.34 | 0.12 | 0.62 |
| Taurine | 2.69 | 0.14 | 0.56 | 0.13 | 0.58 | −0.09 | 0.70 |
| Melatonin | 2.93 | 0.37 | 0.11 | 0.47 | 0.04 * | 0.13 | 0.60 |
| Serotonin | 3.11 | 0.23 | 0.32 | 0.48 | 0.04 * | 0.10 | 0.70 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nik Mohd Fakhruddin, N.N.I.; Shahar, S.; Ismail, I.S.; Ahmad Azam, A.; Rajab, N.F. Urine Untargeted Metabolomic Profiling Is Associated with the Dietary Pattern of Successful Aging among Malaysian Elderly. Nutrients 2020, 12, 2900. https://doi.org/10.3390/nu12102900
Nik Mohd Fakhruddin NNI, Shahar S, Ismail IS, Ahmad Azam A, Rajab NF. Urine Untargeted Metabolomic Profiling Is Associated with the Dietary Pattern of Successful Aging among Malaysian Elderly. Nutrients. 2020; 12(10):2900. https://doi.org/10.3390/nu12102900
Chicago/Turabian StyleNik Mohd Fakhruddin, Nik Nur Izzati, Suzana Shahar, Intan Safinar Ismail, Amalina Ahmad Azam, and Nor Fadilah Rajab. 2020. "Urine Untargeted Metabolomic Profiling Is Associated with the Dietary Pattern of Successful Aging among Malaysian Elderly" Nutrients 12, no. 10: 2900. https://doi.org/10.3390/nu12102900
APA StyleNik Mohd Fakhruddin, N. N. I., Shahar, S., Ismail, I. S., Ahmad Azam, A., & Rajab, N. F. (2020). Urine Untargeted Metabolomic Profiling Is Associated with the Dietary Pattern of Successful Aging among Malaysian Elderly. Nutrients, 12(10), 2900. https://doi.org/10.3390/nu12102900