Blends of Human Milk Oligosaccharides Confer Intestinal Epithelial Barrier Protection In Vitro

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Cell Culture Model
2.3. HMO Treatments and Inflammation-Induced Epithelial Barrier Dysfunction Model
2.4. Epithelial Permeability Assessment
2.5. Experimental Design and Data Analyses
3. Results
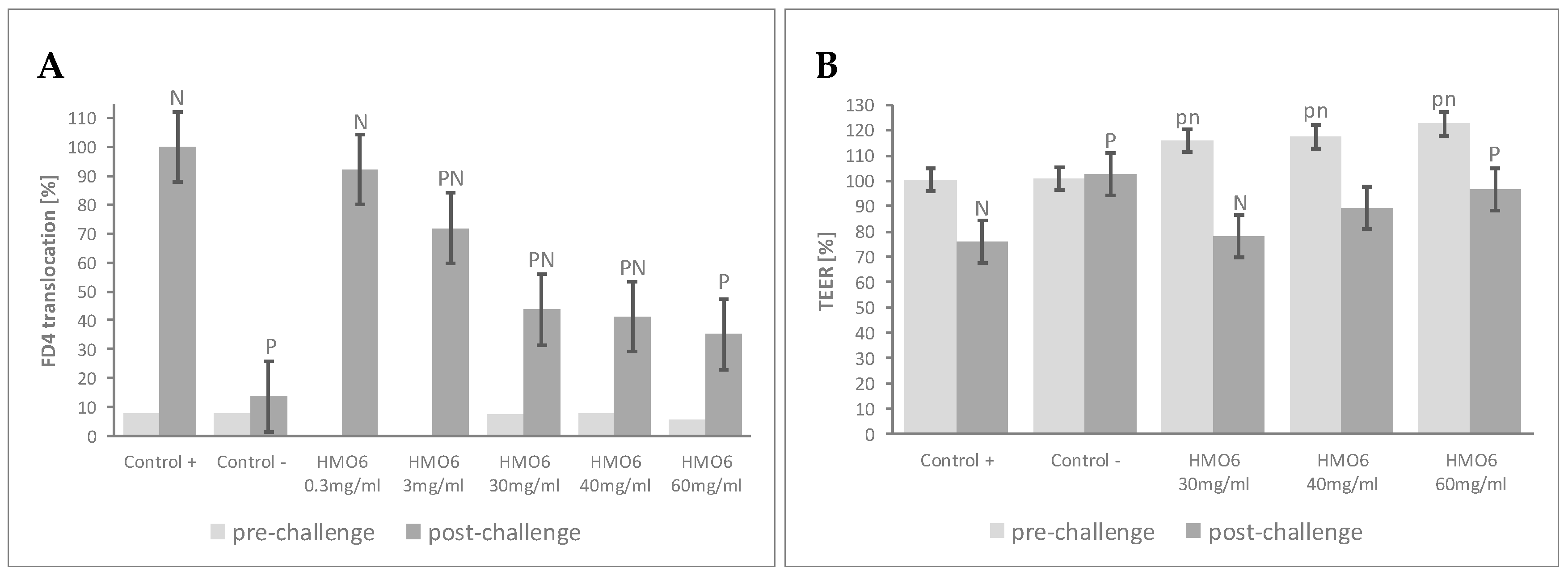
3.1. A Blend of 6 HMOs Highly Represented in Human Milk Conferred Resistance against Inflammatory-Induced Epithelial Barrier Dysfunction
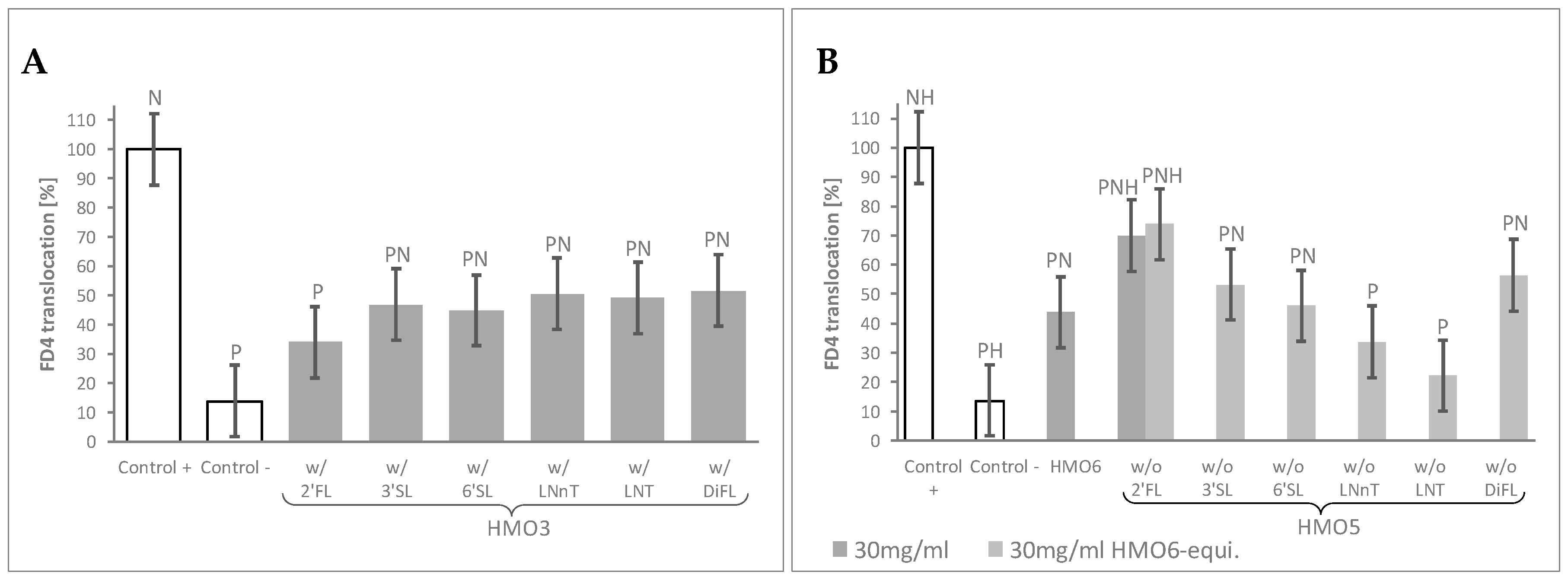
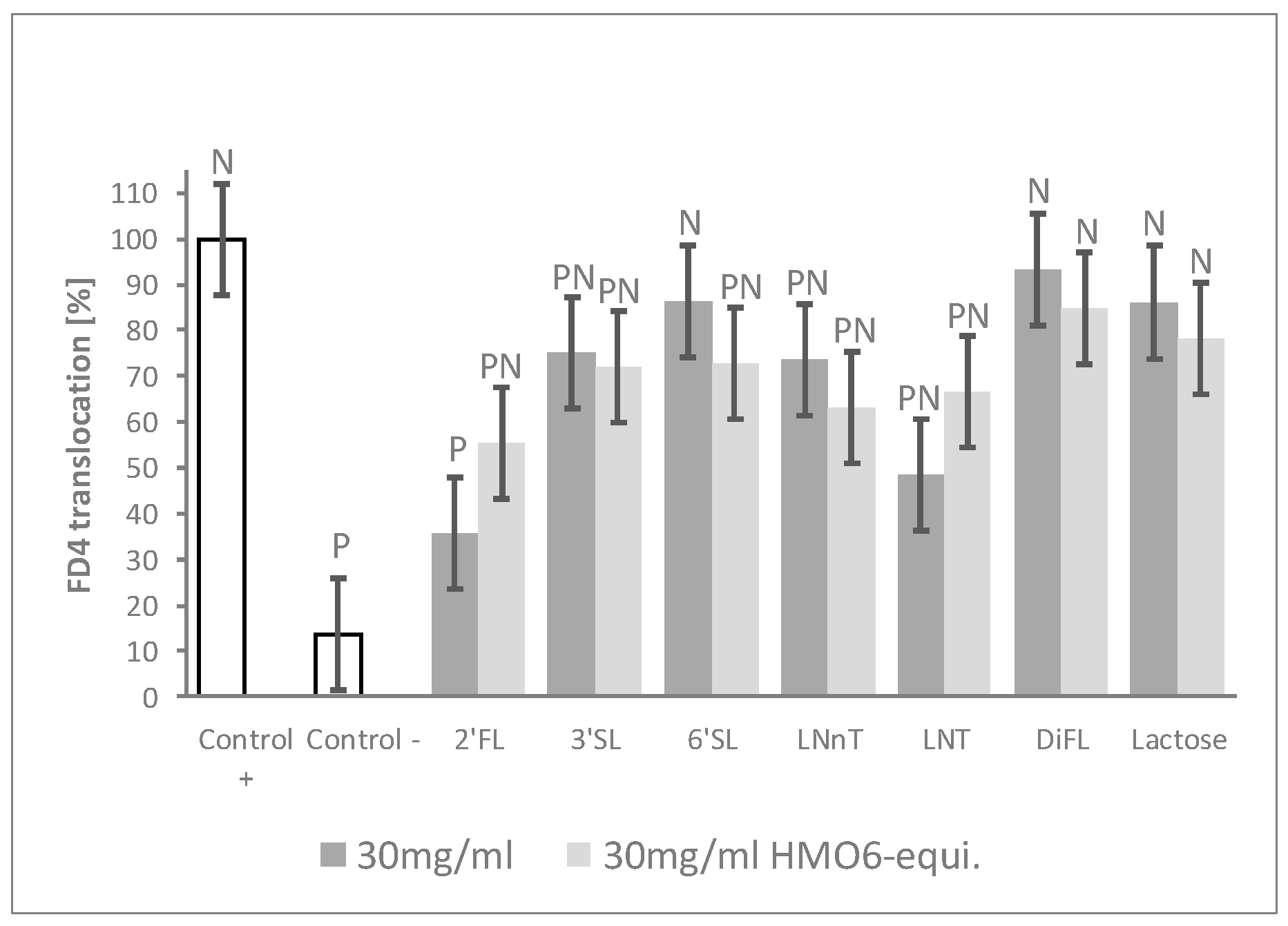
3.2. Combination of HMO Sub-Blends, Particularly Those Containing 2′FL, Protected the Epithelial Barrier
3.3. 2’FL Was the Most Efficient Single HMO in Preventing the Increased Epithelial Permeability after Inflammatory Challenge
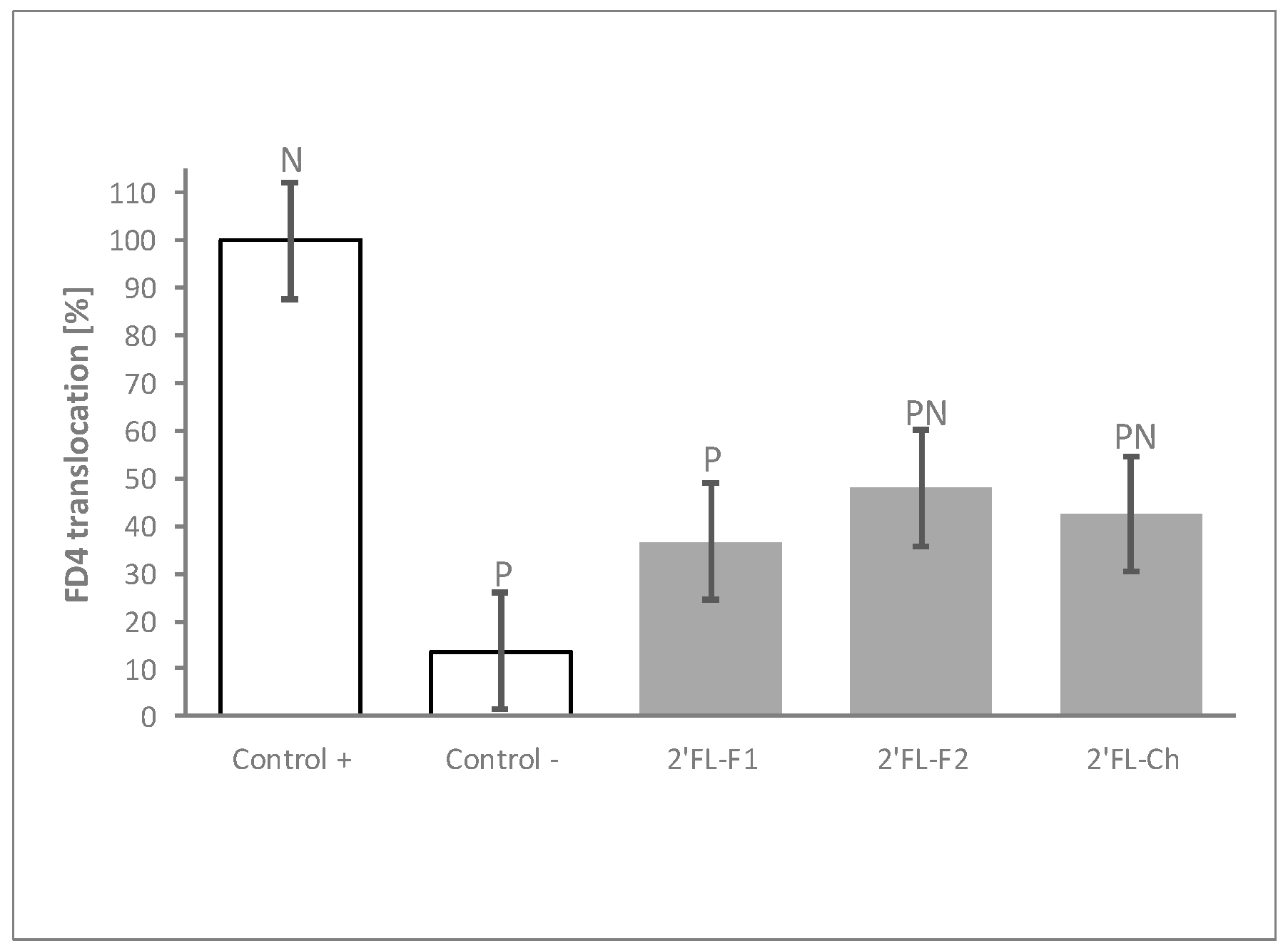
3.4. The Process of 2′FL Synthesis Does Not Influence Its Permeability Benefits
4. Discussion
5. Conclusion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Fifty-Fifth World Health Assembly; World Health Organization: Geneva, Switzerland, 2002. [Google Scholar]
- Herrmann, K.; Carroll, K. An exclusively human milk diet reduces necrotizing enterocolitis. Breastfeed. Med. 2014, 9, 184–190. [Google Scholar] [CrossRef] [Green Version]
- Klement, E.; Cohen, R.V.; Boxman, J.; Joseph, A.; Reif, S. Breastfeeding and risk of inflammatory bowel disease: A systematic review with meta-analysis. Am. J. Clin. Nutr. 2004, 80, 1342–1352. [Google Scholar] [CrossRef] [Green Version]
- Lodge, C.J.; Tan, D.J.; Lau, M.X.; Dai, X.; Tham, R.; Lowe, A.J.; Bowatte, G.; Allen, K.J.; Dharmage, S.C. Breastfeeding and asthma and allergies: A systematic review and meta-analysis. Acta Paediatr. 2015, 104, 38–53. [Google Scholar] [CrossRef]
- Jakaitis, B.M.; Denning, P.W. Human breast milk and the gastrointestinal innate immune system. Clin. Perinatol. 2014, 41, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Jakobsson, I. Intestinal permeability in children of different ages and with different gastrointestinal diseases. Pediatr. Allergy Immunol. 1993, 4, 33–39. [Google Scholar] [CrossRef]
- Catassi, C.; Bonucci, A.; Coppa, G.V.; Carlucci, A.; Giorgi, P.L. Intestinal permeability changes during the first month: Effect of natural versus artificial feeding. J. Pediatr. Gastroenterol. Nutr. 1995, 21, 383–386. [Google Scholar] [CrossRef]
- Saleem, B.; Okogbule-Wonodi, A.C.; Fasano, A.; Magder, L.S.; Ravel, J.; Kapoor, S.; Viscardi, R.M. Intestinal Barrier Maturation in Very Low Birthweight Infants: Relationship to Feeding and Antibiotic Exposure. J. Pediatr. 2017, 183, 31–36.e31. [Google Scholar] [CrossRef] [Green Version]
- Arndt, M.B.; Richardson, B.A.; Ahmed, T.; Mahfuz, M.; Haque, R.; John-Stewart, G.C.; Denno, D.M.; Petri, W.A., Jr.; Kosek, M.; Walson, J.L.; et al. Fecal Markers of Environmental Enteropathy and Subsequent Growth in Bangladeshi Children. Am. J. Trop. Med. Hyg. 2016, 95, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Fukui, H. Increased Intestinal Permeability and Decreased Barrier Function: Does It Really Influence the Risk of Inflammation? Inflamm. Intest. Dis. 2016, 1, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Heymann, J.; Raub, A.; Earle, A. Breastfeeding policy: A globally comparative analysis. Bull. World Health Organ. 2013, 91, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Bode, L. Human milk oligosaccharides: Every baby needs a sugar mama. Glycobiology 2012, 22, 1147–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triantis, V.; Bode, L.; van Neerven, R.J.J. Immunological Effects of Human Milk Oligosaccharides. Front. Pediatr. 2018, 6, 190. [Google Scholar] [CrossRef] [PubMed]
- Marcobal, A.; Sonnenburg, J.L. Human milk oligosaccharide consumption by intestinal microbiota. Clin. Microbiol. Infect. 2012, 18 (Suppl. 4), 12–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, J.C.; Totten, S.M.; Huang, J.O.; Nagshbandi, S.; Kirmiz, N.; Garrido, D.A.; Lewis, Z.T.; Wu, L.D.; Smilowitz, J.T.; German, J.B.; et al. Identification of Oligosaccharides in Feces of Breast-fed Infants and Their Correlation with the Gut Microbial Community. Mol. Cell. Proteom. 2016, 15, 2987–3002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borewicz, K.; Gu, F.; Saccenti, E.; Arts, I.C.W.; Penders, J.; Thijs, C.; van Leeuwen, S.S.; Lindner, C.; Nauta, A.; van Leusen, E.; et al. Correlating Infant Faecal Microbiota Composition and Human Milk Oligosaccharide Consumption by Microbiota of One-Month Old Breastfed Infants. Mol. Nutr. Food Res. 2019, 63, e1801214. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.Y.; Li, B.; Koike, Y.; Maattanen, P.; Miyake, H.; Cadete, M.; Johnson-Henry, K.C.; Botts, S.R.; Lee, C.; Abrahamsson, T.R.; et al. Human Milk Oligosaccharides Increase Mucin Expression in Experimental Necrotizing Enterocolitis. Mol. Nutr. Food Res. 2019, 63, e1800658. [Google Scholar] [CrossRef] [PubMed]
- Holscher, H.D.; Davis, S.R.; Tappenden, K.A. Human milk oligosaccharides influence maturation of human intestinal Caco-2Bbe and HT-29 cell lines. J. Nutr. 2014, 144, 586–591. [Google Scholar] [CrossRef] [Green Version]
- Holscher, H.D.; Bode, L.; Tappenden, K.A. Human Milk Oligosaccharides Influence Intestinal Epithelial Cell Maturation in Vitro. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 296–301. [Google Scholar] [CrossRef]
- Samuel, T.M.; Binia, A.; de Castro, C.A.; Thakkar, S.K.; Billeaud, C.; Agosti, M.; Al-Jashi, I.; Costeira, M.J.; Marchini, G.; Martinez-Costa, C.; et al. Impact of maternal characteristics on human milk oligosaccharide composition over the first 4 months of lactation in a cohort of healthy European mothers. Sci. Rep. 2019, 9, 11767. [Google Scholar] [CrossRef]
- Poquet, L.; Clifford, M.N.; Williamson, G. Transport and metabolism of ferulic acid through the colonic epithelium. Drug Metab. Dispos. 2008, 36, 190–197. [Google Scholar] [CrossRef]
- R 3.5.0 R Foundation for Statistical Computing. Available online: http://www.R-project.org (accessed on 1 October 2020).
- Ayechu-Muruzabal, V.; van Stigt, A.H.; Mank, M.; Willemsen, L.E.M.; Stahl, B.; Garssen, J.; Van’t Land, B. Diversity of Human Milk Oligosaccharides and Effects on Early Life Immune Development. Front. Pediatr. 2018, 6, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ninonuevo, M.R.; Park, Y.; Yin, H.; Zhang, J.; Ward, R.E.; Clowers, B.H.; German, J.B.; Freeman, S.L.; Killeen, K.; Grimm, R.; et al. A strategy for annotating the human milk glycome. J. Agric. Food Chem. 2006, 54, 7471–7480. [Google Scholar] [CrossRef] [PubMed]
- McGuire, M.K.; Meehan, C.L.; McGuire, M.A.; Williams, J.E.; Foster, J.; Sellen, D.W.; Kamau-Mbuthia, E.W.; Kamundia, E.W.; Mbugua, S.; Moore, S.E.; et al. What’s normal? Oligosaccharide concentrations and profiles in milk produced by healthy women vary geographically. Am. J. Clin. Nutr. 2017, 105, 1086–1100. [Google Scholar] [CrossRef] [PubMed]
- Gabrielli, O.; Zampini, L.; Galeazzi, T.; Padella, L.; Santoro, L.; Peila, C.; Giuliani, F.; Bertino, E.; Fabris, C.; Coppa, G.V. Preterm milk oligosaccharides during the first month of lactation. Pediatr.s 2011, 128, e1520–e1531. [Google Scholar] [CrossRef]
- Newburg, D.S.; Ruiz-Palacios, G.M.; Altaye, M.; Chaturvedi, P.; Meinzen-Derr, J.; Guerrero Mde, L.; Morrow, A.L. Innate protection conferred by fucosylated oligosaccharides of human milk against diarrhea in breastfed infants. Glycobiology 2004, 14, 253–263. [Google Scholar] [CrossRef]
- Sprenger, N.; Odenwald, H.; Kukkonen, A.K.; Kuitunen, M.; Savilahti, E.; Kunz, C. FUT2-dependent breast milk oligosaccharides and allergy at 2 and 5 years of age in infants with high hereditary allergy risk. Eur. J. Nutr. 2017, 56, 1293–1301. [Google Scholar] [CrossRef] [Green Version]
- Sodhi, C.P.; Wipf, P.; Yamaguchi, Y.; Fulton, W.B.; Kovler, M.; Nino, D.F.; Zhou, Q.; Banfield, E.; Werts, A.D.; Ladd, M.R.; et al. The human milk oligosaccharides 2′-fucosyllactose and 6′-sialyllactose protect against the development of necrotizing enterocolitis by inhibiting toll-like receptor 4 signaling. Pediatr. Res. 2020, 1–11. [Google Scholar] [CrossRef]
- Kong, C.; Elderman, M.; Cheng, L.; de Haan, B.J.; Nauta, A.; de Vos, P. Modulation of Intestinal Epithelial Glycocalyx Development by Human Milk Oligosaccharides and Non-Digestible Carbohydrates. Mol. Nutr. Food Res. 2019, 63, e1900303. [Google Scholar] [CrossRef]
- Zenhom, M.; Hyder, A.; de Vrese, M.; Heller, K.J.; Roeder, T.; Schrezenmeir, J. Prebiotic oligosaccharides reduce proinflammatory cytokines in intestinal Caco-2 cells via activation of PPARgamma and peptidoglycan recognition protein 3. J. Nutr. 2011, 141, 971–977. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Liu, S.; Kling, D.E.; Leone, S.; Lawlor, N.T.; Huang, Y.; Feinberg, S.B.; Hill, D.R.; Newburg, D.S. The human milk oligosaccharide 2′-fucosyllactose modulates CD14 expression in human enterocytes, thereby attenuating LPS-induced inflammation. Gut 2016, 65, 33–46. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blocks | Treatments | 2′FL | 3′SL | 6′SL | LNnT | LNT | DFL | Lactose | |
|---|---|---|---|---|---|---|---|---|---|
| [mg/mL] | |||||||||
| A | HMO6 | 0.3 mg/mL | 0.165 | 0.021 | 0.027 | 0.015 | 0.054 | 0.018 | 0.000 |
| 3 mg/mL | 1.65 | 0.21 | 0.27 | 0.15 | 0.54 | 0.18 | 0.00 | ||
| 30 mg/mL | 16.5 | 2.1 | 2.7 | 1.5 | 5.4 | 1.8 | 0.0 | ||
| 40 mg/mL | 22.0 | 2.8 | 3.6 | 2.0 | 7.2 | 2.4 | 0.0 | ||
| 60 mg/mL | 33.0 | 4.2 | 5.4 | 3.0 | 10.8 | 3.6 | 0.0 | ||
| B | HMO3_01 | 60 mg/mL HMO6-equivalent | 33.0 | 4.2 | 5.4 | 0.0 | 0.0 | 0.0 | 0.0 |
| HMO3_02 | 33.0 | 4.2 | 0.0 | 3.0 | 0.0 | 0.0 | 0.0 | ||
| HMO3_03 | 33.0 | 4.2 | 0.0 | 0.0 | 10.8 | 0.0 | 0.0 | ||
| HMO3_04 | 33.0 | 4.2 | 0.0 | 0.0 | 0.0 | 3.6 | 0.0 | ||
| HMO3_05 | 33.0 | 0.0 | 5.4 | 3.0 | 0.0 | 0.0 | 0.0 | ||
| HMO3_06 | 33.0 | 0.0 | 5.4 | 0.0 | 10.8 | 0.0 | 0.0 | ||
| HMO3_07 | 33.0 | 0.0 | 5.4 | 0.0 | 0.0 | 3.6 | 0.0 | ||
| HMO3_08 | 33.0 | 0.0 | 0.0 | 3.0 | 10.8 | 0.0 | 0.0 | ||
| HMO3_09 | 33.0 | 0.0 | 0.0 | 3.0 | 0.0 | 3.6 | 0.0 | ||
| HMO3_10 | 33.0 | 0.0 | 0.0 | 0.0 | 10.8 | 3.6 | 0.0 | ||
| HMO3_11 | 0.0 | 4.2 | 5.4 | 3.0 | 0.0 | 0.0 | 0.0 | ||
| HMO3_12 | 0.0 | 4.2 | 5.4 | 0.0 | 10.8 | 0.0 | 0.0 | ||
| HMO3_13 | 0.0 | 4.2 | 5.4 | 0.0 | 0.0 | 3.6 | 0.0 | ||
| HMO3_14 | 0.0 | 4.2 | 0.0 | 3.0 | 10.8 | 0.0 | 0.0 | ||
| HMO3_15 | 0.0 | 4.2 | 0.0 | 3.0 | 0.0 | 3.6 | 0.0 | ||
| HMO3_16 | 0.0 | 4.2 | 0.0 | 0.0 | 10.8 | 3.6 | 0.0 | ||
| HMO3_17 | 0.0 | 0.0 | 5.4 | 3.0 | 10.8 | 0.0 | 0.0 | ||
| HMO3_18 | 0.0 | 0.0 | 5.4 | 3.0 | 0.0 | 3.6 | 0.0 | ||
| HMO3_19 | 0.0 | 0.0 | 5.4 | 0.0 | 10.8 | 3.6 | 0.0 | ||
| HMO3_20 | 0.0 | 0.0 | 0.0 | 3.0 | 10.8 | 3.6 | 0.0 | ||
| C | HMO5 (w/o 2′FL) | 30 mg/mL HMO6-equivalent | 0.0 | 2.1 | 2.7 | 1.5 | 5.4 | 1.8 | 0.0 |
| HMO5 (w/o 3′SL) | 16.5 | 0.0 | 2.7 | 1.5 | 5.4 | 1.8 | 0.0 | ||
| HMO5 (w/o 6′SL) | 16.5 | 2.1 | 0.0 | 1.5 | 5.4 | 1.8 | 0.0 | ||
| HMO5 (w/o LNnT) | 16.5 | 2.1 | 2.7 | 0.0 | 5.4 | 1.8 | 0.0 | ||
| HMO5 (w/o LNT) | 16.5 | 2.1 | 2.7 | 1.5 | 0.0 | 1.8 | 0.0 | ||
| HMO5 (w/o DFL) | 16.5 | 2.1 | 2.7 | 1.5 | 5.4 | 0.0 | 0.0 | ||
| HMO5 (w/o 2′FL) | 30mg/mL | 0.0 | 4.7 | 6.0 | 3.3 | 12.0 | 4.0 | 0.0 | |
| D | 2′FL | 30mg/mL HMO6-equivalent | 16.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| 3′SL | 0.0 | 2.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | ||
| 6′SL | 0.0 | 0.0 | 2.7 | 0.0 | 0.0 | 0.0 | 0.0 | ||
| LNnT | 0.0 | 0.0 | 0.0 | 1.5 | 0.0 | 0.0 | 0.0 | ||
| LNT | 0.0 | 0.0 | 0.0 | 0.0 | 5.4 | 0.0 | 0.0 | ||
| DFL | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.8 | 0.0 | ||
| Lactose | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.5 | ||
| 2′FL | 30 mg/mL | 30.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 3′SL | 0.0 | 30.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | ||
| 6′SL | 0.0 | 0.0 | 30.0 | 0.0 | 0.0 | 0.0 | 0.0 | ||
| LNnT | 0.0 | 0.0 | 0.0 | 30.0 | 0.0 | 0.0 | 0.0 | ||
| LNT | 0.0 | 0.0 | 0.0 | 0.0 | 30.0 | 0.0 | 0.0 | ||
| DFL | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 30.0 | 0.0 | ||
| Lactose | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 30.0 | ||
| 2′FL Ferment. 1 | 30 mg/mL | 30.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 2′FL Ferment. 2 | 30.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | ||
| 2′FL Chemical | 30.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Natividad, J.M.; Rytz, A.; Keddani, S.; Bergonzelli, G.; Garcia-Rodenas, C.L. Blends of Human Milk Oligosaccharides Confer Intestinal Epithelial Barrier Protection In Vitro. Nutrients 2020, 12, 3047. https://doi.org/10.3390/nu12103047
Natividad JM, Rytz A, Keddani S, Bergonzelli G, Garcia-Rodenas CL. Blends of Human Milk Oligosaccharides Confer Intestinal Epithelial Barrier Protection In Vitro. Nutrients. 2020; 12(10):3047. https://doi.org/10.3390/nu12103047
Chicago/Turabian StyleNatividad, Jane M., Andreas Rytz, Sonia Keddani, Gabriela Bergonzelli, and Clara L. Garcia-Rodenas. 2020. "Blends of Human Milk Oligosaccharides Confer Intestinal Epithelial Barrier Protection In Vitro" Nutrients 12, no. 10: 3047. https://doi.org/10.3390/nu12103047
APA StyleNatividad, J. M., Rytz, A., Keddani, S., Bergonzelli, G., & Garcia-Rodenas, C. L. (2020). Blends of Human Milk Oligosaccharides Confer Intestinal Epithelial Barrier Protection In Vitro. Nutrients, 12(10), 3047. https://doi.org/10.3390/nu12103047