A Mix of Natural Bioactive Compounds Reduces Fat Accumulation and Modulates Gene Expression in the Adipose Tissue of Obese Rats Fed a Cafeteria Diet



Abstract
:1. Introduction
2. Materials and Methods
2.1. Mix Composition
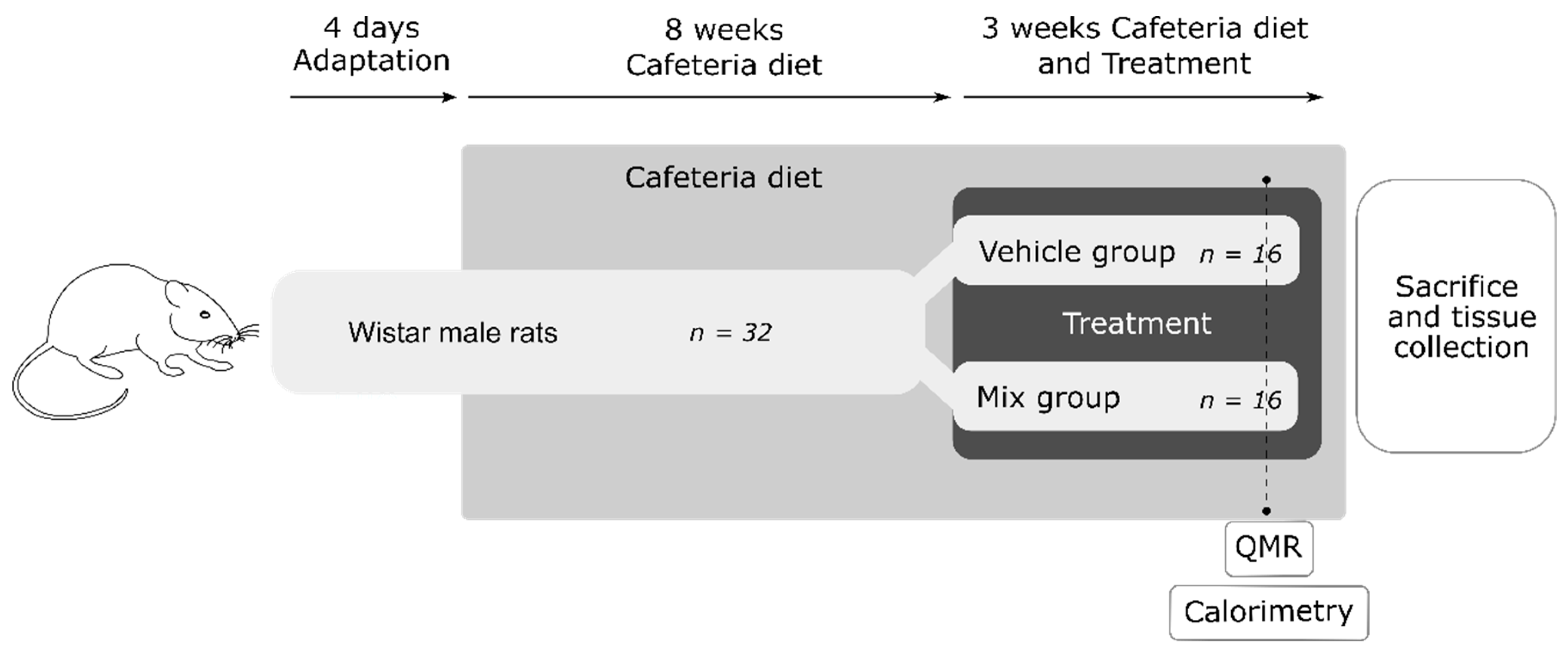
2.2. Animals and Treatments
2.3. Indirect Calorimetry
2.4. Cell Culture and Treatment
2.5. RNA Extraction and Quantification by Real-Time qRT-PCR
2.6. Histology Analysis
2.7. Western Blot Analysis
2.8. Stromal Vascular Fraction Extraction
2.9. Statistical Analysis
3. Results
3.1. Biometric Measures
3.2. Indirect Calorimetry
3.3. Gene Expression in IWAT, EWAT, RWAT, BAT, and SVF
3.4. Histology of the IWAT
3.5. 3T3-L1 Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO). Obesity and overweight, Factsheet No. 311. Available online: http://www.who.int/mediacentre/factsheets/fs311/en/ (accessed on 13 February 2018).
- Poirier, P.; Giles, T.D.; Bray, G.A.; Hong, Y.; Stern, J.S.; Pi-Sunyer, F.X.; Eckel, R.H. Obesity and cardiovascular disease: Pathophysiology, evaluation, and effect of weight loss: An update of the 1997 American Heart Association Scientific Statement on obesity and heart disease from the Obesity Committee of the Council on Nutrition, Physical. Circulation 2006, 113, 898–918. [Google Scholar] [CrossRef] [Green Version]
- Weinstock, R.S.; Dai, H.; Wadden, T.A. Diet and exercise in the treatment of obesity. Arch. Intern. Med. 1998, 158, 2477–2483. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.; Gavrilova, O.; Pack, S.; Jou, W.; Mullen, S.; Sumner, A.E.; Cushman, S.W.; Periwal, V. Hypertrophy and/or Hyperplasia: Dynamics of Adipose Tissue Growth. PLoS Comput. Biol. 2009, 5. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Vela, M.E.F.; Torres, N.; Tovar, A.R. White Adipose Tissue as Endocrine Organ and Its Role in Obesity. Arch. Med. Res. 2008, 39, 715–728. [Google Scholar] [CrossRef]
- Sanchez-Gurmaches, J.; Guertin, D.A. Adipocyte lineages: Tracing back the origins of fat. Biochim. Biophys. Acta Mol. Basis Dis. 2014, 1842, 340–351. [Google Scholar] [CrossRef] [Green Version]
- Pinent, M.; Bladé, M.C.; Salvadó, M.J.; Arola, L.; Hackl, H.; Quackenbush, J.; Trajanoski, Z.; Ardévol, A. Grape-seed derived procyanidins interfere with adipogenesis of 3T3-L1 cells at the onset of differentiation. Int. J. Obes. 2005, 29, 934–941. [Google Scholar] [CrossRef] [Green Version]
- Han, M.H.; Kim, H.J.; Jeong, J.-W.; Park, C.; Kim, B.W.; Choi, Y.H. Inhibition of Adipocyte Differentiation by Anthocyanins Isolated from the Fruit of Vitis coignetiae Pulliat is Associated with the Activation of AMPK Signaling Pathway. Toxicol. Res. 2018, 34, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Yeganeh, A.; Taylor, C.G.; Tworek, L.; Poole, J.; Zahradka, P. Trans-10,cis-12 conjugated linoleic acid (CLA) interferes with lipid droplet accumulation during 3T3-L1 preadipocyte differentiation. Int. J. Biochem. Cell Biol. 2016, 76, 39–50. [Google Scholar] [CrossRef]
- Chechi, K.; Carpentier, A.C.; Richard, D. Understanding the brown adipocyte as a contributor to energy homeostasis. Trends Endocrinol. Metab. 2013, 24, 408–420. [Google Scholar] [CrossRef] [PubMed]
- Moroshko, I.; Brennan, L.; Brien, P.O. Predictors of dropout in weight loss interventions: A systematic review of the literature. Obes. Rev. 2011, 912–934. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.G.; Park, C.-Y. Anti-Obesity Drugs: A Review about Their Effects and Safety. Diabetes Metab. J. 2012, 36, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Torres-Fuentes, C.; Schellekens, H.; Dinan, T.G.; Cryan, J.F. A natural solution for obesity: Bioactives for the prevention and treatment of weight gain. A review. Am. J. Clin. Nutr. 2015, 18, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Bravo Vázquez, F.I.; Suárez Recio, M.; Arola Arnal, A.; Muguerza Marquínez, M.B.; Salvadó Rovira, M.J.; Arola Ferrer, L.M.; Crescenti Savall, A.; Pascual Serrano, A.; Ibars Serra, M.; Ardid Ruiz, A.; et al. Composición hipoleptinémica y su uso; P201930732; Oficina Española de Patentes y Marcas: Madrid, Spain, 2019; p. 15.
- Caimari, A.; Del Bas, J.; Crescenti, A.; Arola, L. Low doses of grape seed procyanidins reduce adiposity and improve the plasma lipid profile in hamsters. Int. J. Obes. 2013, 37, 576–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascual-Serrano, A.; Arola-Arnal, A.; Suárez-García, S.; Bravo, F.I.; Suárez, M.; Arola, L.; Bladé, C. Grape seed proanthocyanidin supplementation reduces adipocyte size and increases adipocyte number in obese rats. Int. J. Obes. 2017, 41, 1246–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.; Jiang, Z.; Yin, J.; Long, H.; Zheng, X. Anti-obesity effects of artificial planting blueberry (Vaccinium ashei) anthocyanin in high-fat diet-treated mice. Int. J. Food Sci. Nutr. 2016, 67, 257–264. [Google Scholar] [CrossRef]
- Den Hartigh, L.J.; Wang, S.; Goodspeed, L.; Wietecha, T.; Houston, B.; Omer, M.; Ogimoto, K.; Subramanian, S.; Gowda, G.A.N.; O’Brien, K.D.; et al. Metabolically distinct weight loss by 10,12 CLA and caloric restriction highlight the importance of subcutaneous white adipose tissue for glucose homeostasis in mice. PLoS ONE 2017, 12, e0172912. [Google Scholar] [CrossRef] [Green Version]
- Bravo, F.I.; Mas-Capdevila, A.; Margalef, M.; Arola-Arnal, A.; Muguerza, B. Novel Antihypertensive Peptides Derived from Chicken Foot Proteins. Mol. Nutr. Food Res. 2019, 63, 1801176. [Google Scholar] [CrossRef]
- Mas-Capdevila, A.; Pons, Z.; Aleixandre, A.; Bravo, F.; Muguerza, B. Dose-Related Antihypertensive Properties and the Corresponding Mechanisms of a Chicken Foot Hydrolysate in Hypertensive Rats. Nutrients 2018, 10, 1295. [Google Scholar] [CrossRef] [Green Version]
- Salvadó, M.J.; Casanova, E.; Fernández-Iglesias, A.; Arola, L.; Bladé, C. Roles of proanthocyanidin rich extracts in obesity. Food Funct. 2015, 6, 1053–1071. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Ling, W. The update of anthocyanins on obesity and type 2 diabetes: Experimental evidence and clinical perspectives. Rev. Endocr. Metab. Disord. 2015, 16, 1–13. [Google Scholar] [CrossRef]
- Yamasaki, M.; Yanagita, T. Adipocyte response to conjugated linoleic acid. Obes. Res. Clin. Pract. 2013, 7, e235–e242. [Google Scholar] [CrossRef] [PubMed]
- Margalef, M.; Pons, Z.; Iglesias-Carres, L.; Bravo, F.I.; Muguerza, B.; Arola-Arnal, A. Lack of Tissue Accumulation of Grape Seed Flavanols after Daily Long-Term Administration in Healthy and Cafeteria-Diet Obese Rats. J. Agric. Food Chem. 2015, 63, 9996–10003. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Xia, M.; Ma, J.; Hao, Y.; Liu, J.; Mou, H.; Cao, L.; Ling, W. Anthocyanin supplementation improves serum LDL- and HDL-cholesterol concentrations associated with the inhibition of cholesteryl ester transfer protein in dyslipidemic subjects. Am. J. Clin. Nutr. 2009, 90, 485–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bravo Vázquez, F.I.; Arola Ferrer, L.; Muguerza Marquínez, M.B. Method for Obtaining a Chicken Feet Hydrolysate with Antihypertensive Activity, the Hydrolysate That Is Obtained and the Peptides it Contains. WO Patent WO/2017/050775, 30 March 2017. [Google Scholar]
- Gomez-Smith, M.; Karthikeyan, S.; Jeffers, M.S.; Janik, R.; Thomason, L.A.; Stefanovic, B.; Corbett, D. A physiological characterization of the Cafeteria diet model of metabolic syndrome in the rat. Physiol. Behav. 2016, 167, 382–391. [Google Scholar] [CrossRef]
- Nair, A.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27. [Google Scholar] [CrossRef] [Green Version]
- Crescenti, A.; del Bas, J.M.; Arola-Arnal, A.; Oms-Oliu, G.; Arola, L.; Caimari, A. Grape seed procyanidins administered at physiological doses to rats during pregnancy and lactation promote lipid oxidation and up-regulate AMPK in the muscle of male offspring in adulthood. J. Nutr. Biochem. 2015, 26, 912–920. [Google Scholar] [CrossRef]
- Frayn, K.N. Calculation of substrate oxidation rates in vivo from gaseous exchange. J. Appl. Physiol. 1983, 55, 628–634. [Google Scholar] [CrossRef] [Green Version]
- Carraro, F.; Stuart, C.A.; Hartl, W.H.; Rosenblatt, J.; Wolfe, R.R. Effect of exercise and recovery on muscle protein synthesis in human subjects. Am. J. Physiol. 1990, 259, E470–E476. [Google Scholar] [CrossRef]
- Atwater, W.O. Coefficients of digestibility and availability of the nutrients of food. Proc Am Physiol Soc 1909, 30, 14–19. [Google Scholar]
- Watanabe, N.; Inagawa, K.; Shibata, M.; Osakabe, N. Flavan-3-ol fraction from cocoa powder promotes mitochondrial biogenesis in skeletal muscle in mice. Lipids Health Dis. 2014, 13, 64. [Google Scholar] [CrossRef] [Green Version]
- You, Y.; Yuan, X.; Liu, X.; Liang, C.; Meng, M.; Huang, Y.; Han, X.; Guo, J.; Guo, Y.; Ren, C.; et al. Cyanidin-3-glucoside increases whole body energy metabolism by upregulating brown adipose tissue mitochondrial function. Mol. Nutr. Food Res. 2017, 61, 1700261. [Google Scholar] [CrossRef] [PubMed]
- Ardévol, A.; Bladé, C.; Salvadó, M.J.; Arola, L. Changes in lipolysis and hormone-sensitive lipase expression caused by procyanidins in 3T3-L1 adipocytes. Int. J. Obes. 2000, 24, 319–324. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Zacarías, J.L.; Castro-Muñozledo, F.; Kuri-Harcuch, W. Quantitation of adipose conversion and triglycerides by staining intracytoplasmic lipids with Oil red O. Histochemistry 1992, 97, 493–497. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Gibert-Ramos, A.; Palacios-Jordan, H.; Salvadó, M.J.; Crescenti, A. Consumption of out-of-season orange modulates fat accumulation, morphology and gene expression in the adipose tissue of Fischer 344 rats. Eur. J. Nutr. 2019. [Google Scholar] [CrossRef] [Green Version]
- Weiss, E.P.; Jordan, R.C.; Frese, E.M.; Albert, S.G.; Villareal, D.T. Effects of Weight Loss on Lean Mass, Strength, Bone, and Aerobic Capacity. Med Sci Sport. Exerc. 2017, 49, 206–217. [Google Scholar] [CrossRef]
- Guglielmi, V.; Sbraccia, P. Obesity phenotypes: Depot-differences in adipose tissue and their clinical implications. Eat. Weight Disord. 2018, 23, 3–14. [Google Scholar] [CrossRef]
- Tchkonia, T.; Tchoukalova, Y.D.; Giorgadze, N.; Pirtskhalava, T.; Karagiannides, I.; Forse, R.A.; Koo, A.; Stevenson, M.; Chinnappan, D.; Cartwright, A.; et al. Abundance of two human preadipocyte subtypes with distinct capacities for replication, adipogenesis, and apoptosis varies among fat depots. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E267–E277. [Google Scholar] [CrossRef]
- Rodríguez, E.; Monjo, M.; Rodríguez-Cuenca, S.; Pujol, E.; Amengual, B.; Roca, P.; Palou, A. Sexual dimorphism in the adrenergic control of rat brown adipose tissue response to overfeeding. Pflugers Arch. Eur. J. Physiol. 2001, 442, 396–403. [Google Scholar] [CrossRef]
- Roca, P.; Rodriguez, A.M.; Oliver, P.; Bonet, M.L.; Quevedo, S.; Picó, C.; Palou, A. Brown adipose tissue response to cafeteria diet-feeding involves induction of the UCP2 gene and is impaired in female rats as compared to males. Pflügers Arch. Eur. J. Physiol. 1999, 438, 628–634. [Google Scholar] [CrossRef]
- Matamala, J.C.; Gianotti, M.; Pericás, J.; Quevedo, S.; Roca, P.; Palou, A.; García-Palmer, F.J. Changes induced by fasting and dietetic obesity in thermogenic parameters of rat brown adipose tissue mitochondrial subpopulations. Biochem. J. 1996, 319, 529–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sene-Fiorese, M.; Duarte, F.O.; Scarmagnani, F.R.R.; Cheik, N.C.; Manzoni, M.S.J.; Nonaka, K.O.; Rossi, E.A.; De Duarte, A.C.G.O.; Dâmaso, A.R. Efficiency of Intermittent Exercise on Adiposity and Fatty Liver in Rats Fed with High-fat Diet. Obesity 2008, 16, 2217–2222. [Google Scholar] [CrossRef] [PubMed]
- Pajuelo, D.; Quesada, H.; Díaz, S.; Fernández-Iglesias, A.; Arola-Arnal, A.; Bladé, C.; Salvadó, J.; Arola, L. Chronic dietary supplementation of proanthocyanidins corrects the mitochondrial dysfunction of brown adipose tissue caused by diet-induced obesity in Wistar rats. Br. J. Nutr. 2012, 107, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Nomura, S.; Ichinose, T.; Jinde, M.; Kawashima, Y.; Tachiyashiki, K.; Imaizumi, K. Tea catechins enhance the mRNA expression of uncoupling protein 1 in rat brown adipose tissue. J. Nutr. Biochem. 2008. [Google Scholar] [CrossRef] [PubMed]
- Serrano, J.; Casanova, À.; Andreu, M.; Anna, G.; Pérez, M. A specific dose of grape seed derived proanthocyanidins to inhibit body weight gain limits food intake and increases energy expenditure in rats. Eur. J. Nutr. 2017, 56, 1629–1636. [Google Scholar] [CrossRef]
- Casanova, E.; Baselga-Escudero, L.; Ribas-Latre, A.; Cedó, L.; Arola-Arnal, A.; Pinent, M.; Bladé, C.; Arola, L.; Salvadó, M.J. Chronic intake of proanthocyanidins and docosahexaenoic acid improves skeletal muscle oxidative capacity in diet-obese rats. J. Nutr. Biochem. 2014, 25, 1003–1010. [Google Scholar] [CrossRef] [PubMed]
- Terpstra, A.H.M.; Beynen, A.C.; Everts, H.; Kocsis, S.; Katan, M.B.; Zock, P.L. The Decrease in Body Fat in Mice Fed Conjugated Linoleic Acid Is Due to Increases in Energy Expenditure and Energy Loss in the Excreta. J. Nutr. 2002, 132, 940–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, R.E.; Ahmadian, M.; Jaworski, K.; Sarkadi-Nagy, E.; Sul, H.S. Regulation of Lipolysis in Adipocytes. Annu. Rev. Nutr. 2007, 27, 79–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caimari, A.; Oliver, P.; Palou, A. Adipose triglyceride lipase expression and fasting regulation are differently affected by cold exposure in adipose tissues of lean and obese Zucker rats. J. Nutr. Biochem. 2012, 23, 1041–1050. [Google Scholar] [CrossRef] [PubMed]
- Ahmadian, M.; Duncan, R.E.; Varady, K.A.; Frasson, D.; Hellerstein, M.K.; Birkenfeld, A.L.; Samuel, V.T.; Shulman, G.I.; Wang, Y.; Kang, C.; et al. Adipose overexpression of desnutrin promotes fatty acid use and attenuates diet-induced obesity. Diabetes 2009, 58, 855–866. [Google Scholar] [CrossRef] [Green Version]
- Blanchard, P.; Festuccia, W.T.; Houde, V.P.; St-pierre, P.; Brûlé, S.; Turcotte, V.; Côté, M.; Bellmann, K.; Marette, A.; Deshaies, Y. Major involvement of mTOR in the PPARγ-induced stimulation of adipose tissue lipid uptake and fat accretion. J. Lipid Res. 2012, 53, 1117–1125. [Google Scholar] [CrossRef] [Green Version]
- Satory, D.L.; Smith, S.B. Conjugated Linoleic Acid Inhibits Proliferation but Stimulates Lipid Filling of Murine 3T3-L1 Preadipocytes. J. Nutr. 1999, 129, 92–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakuma, S.; Nishioka, Y.; Imanishi, R.; Nishikawa, K.; Sakamoto, H.; Fujisawa, J.; Wada, K.; Kamisaki, Y.; Fujimoto, Y. cis9, trans11-Conjugated Linoleic Acid Differentiates Mouse 3T3-L1 Preadipocytes into Mature Small Adipocytes through Induction of Peroxisome Proliferator-activated Receptor. GAMMA. J. Clin. Biochem. Nutr. 2010, 47, 167–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, J.J.; Liu, Z.L.; Li, J.M.; Chen, J.P.; Jiang, L.; Wang, D.M.; Yuan, J.; Shen, J.G.; Yang, D.P.; Chen, J.Q. Different mechanisms of cis-9,trans-11- and trans-10,cis-12- conjugated linoleic acid affecting lipid metabolism in 3T3-L1 cells. J. Nutr. Biochem. 2010, 21, 1099–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Zhang, Y.; Liu, Y.; Sun, R.; Xia, M. Purified anthocyanin supplementation reduces dyslipidemia, enhances antioxidant capacity, and prevents insulin resistance in diabetic patients. J. Nutr. 2015, 145, 742–748. [Google Scholar] [CrossRef]
- Shen, W.; Baldwin, J.; Collins, B.; Hixson, L.; Lee, K.-T.; Herberg, T.; Starnes, J.; Cooney, P.; Chuang, C.-C.; Hopkins, R.; et al. Low level of trans-10, cis-12 conjugated linoleic acid decreases adiposity and increases browning independent of inflammatory signaling in overweight Sv129 mice. J. Nutr. Biochem. 2015, 26, 616–625. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Boström, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.H.; Khandekar, M.; Virtanen, K.A.; Nuutila, P.; Schaart, G.; et al. Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell 2012, 150, 366–376. [Google Scholar] [CrossRef] [Green Version]
- House, R.L.; Cassady, J.P.; Eisen, E.J.; Eling, T.E.; Collins, J.B.; Grissom, S.F.; Odle, J. Functional genomic characterization of delipidation elicited by trans-10, cis-12-conjugated linoleic acid (t10c12-CLA) in a polygenic obese line of mice. Physiol. Genomics 2005, 21, 351–361. [Google Scholar] [CrossRef] [Green Version]
- Wendel, A.A.; Purushotham, A.; Liu, L.-F.; Belury, M.A. Conjugated Linoleic Acid Induces Uncoupling Protein 1 in White Adipose Tissue of ob/ob Mice. Lipids 2009, 44, 975–982. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VH | Mix | p-Value | |
|---|---|---|---|
| Initial weight (g) | 172.60 ± 4.43 | 172.80 ± 5.30 | 0.976 |
| Weight before treatment (g) | 504.50 ± 11.98 | 481.90 ± 7.96 | 0.123 |
| Weight after treatment (g) | 552.27 ± 14.17 | 508.13 ± 9.13 * | 0.013 |
| Weight gain before treatment (%) | 193.82 ± 7.40 | 181.84 ± 7.65 | 0.270 |
| Weight gain during treatment (%) | 9.41 ± 0.52 | 5.42 ± 0.54 * | <0.001 |
| Accumulated caloric intake (Kcal) | 4305.43 ± 245.68 | 3826.15 ± 220.22 | 0.150 |
| Fat (g) | 132.43 ± 6.93 | 109.53 ± 10.76 # | 0.095 |
| Lean (g) | 366.64 ± 7.19 | 373.64 ± 6.14 | 0.471 |
| Fat (%) | 25.01 ± 0.91 | 21.19 ± 1.59 * | 0.054 |
| Lean (%) | 69.58 ± 1.05 | 73.41 ± 1.25 * | 0.035 |
| EWAT (g) | 26.09 ± 1.83 | 19.37 ± 1.37 * | 0.006 |
| IWAT (g) | 10.21 ± 0.68 | 7.99 ± 0.57 * | 0.019 |
| RWAT (g) | 27.13 ± 1.84 | 22.75 ± 1.43 # | 0.068 |
| MWAT (g) | 12.49 ± 1.28 | 10.29 ± 1.00 | 0.182 |
| BAT (g) | 1.064 ± 0.07 | 0.869 ± 0.037 * | 0.017 |
| Adiposity Index (%) | 13.61 ± 0.65 | 11.93 ± 0.52 * | 0.050 |
| EWAT (%) | 4.68 ± 0.25 | 3.78 ± 0.22 * | 0.012 |
| IWAT (%) | 1.84 ± 0.1 | 1.57 ± 0.11 # | 0.086 |
| RWAT (%) | 4.87 ± 0.25 | 4.45 ± 0.24 | 0.233 |
| MWAT (%) | 2.22 ± 0.19 | 1.99 ± 0.16 | 0.351 |
| BAT (%) | 0.192 ± 0.011 | 0.171 ± 0.007 | 0.123 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gibert-Ramos, A.; Martín-González, M.Z.; Crescenti, A.; Salvadó, M.J. A Mix of Natural Bioactive Compounds Reduces Fat Accumulation and Modulates Gene Expression in the Adipose Tissue of Obese Rats Fed a Cafeteria Diet. Nutrients 2020, 12, 3251. https://doi.org/10.3390/nu12113251
Gibert-Ramos A, Martín-González MZ, Crescenti A, Salvadó MJ. A Mix of Natural Bioactive Compounds Reduces Fat Accumulation and Modulates Gene Expression in the Adipose Tissue of Obese Rats Fed a Cafeteria Diet. Nutrients. 2020; 12(11):3251. https://doi.org/10.3390/nu12113251
Chicago/Turabian StyleGibert-Ramos, Albert, Miguel Z. Martín-González, Anna Crescenti, and M. Josepa Salvadó. 2020. "A Mix of Natural Bioactive Compounds Reduces Fat Accumulation and Modulates Gene Expression in the Adipose Tissue of Obese Rats Fed a Cafeteria Diet" Nutrients 12, no. 11: 3251. https://doi.org/10.3390/nu12113251
APA StyleGibert-Ramos, A., Martín-González, M. Z., Crescenti, A., & Salvadó, M. J. (2020). A Mix of Natural Bioactive Compounds Reduces Fat Accumulation and Modulates Gene Expression in the Adipose Tissue of Obese Rats Fed a Cafeteria Diet. Nutrients, 12(11), 3251. https://doi.org/10.3390/nu12113251