The Central Role of Iron in Human Nutrition: From Folk to Contemporary Medicine

 ,
,  ,
,  ,
,  ,
,  ,
,  ,
,
Abstract
:1. Etymology and Ages of Iron Myths
2. The Martial Status in Humans
3. Folk Medicine
4. Current Knowledge on Iron Homeostasis
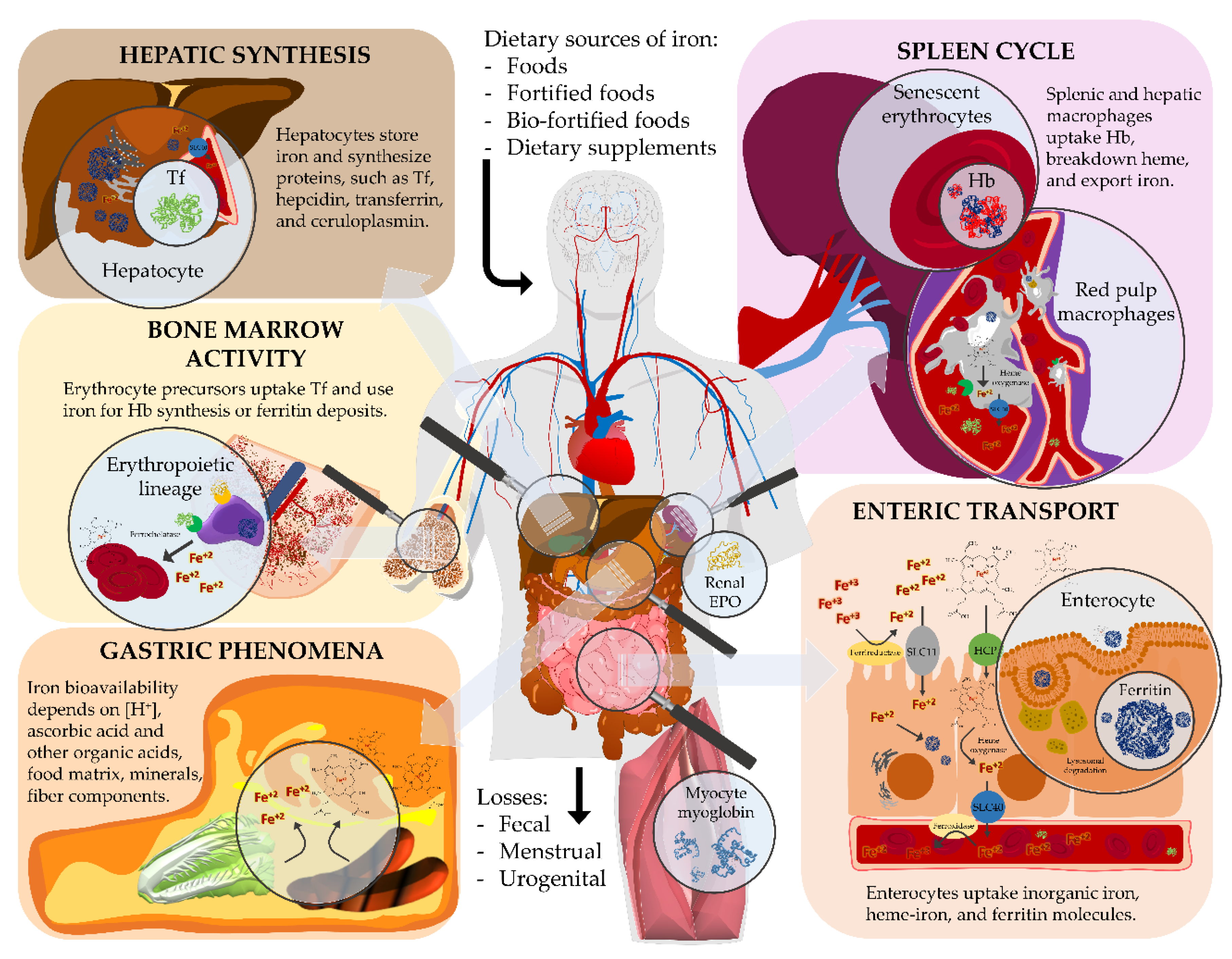
4.1. Overview of Iron Metabolism
4.1.1. Gastric Processing
4.1.2. Intestinal Passage
4.1.3. Systemic Delivery
4.1.4. Physiological Roles
4.1.5. Homeostatic Regulation
4.2. Absorption Influencers
4.3. Diagnostics of Iron Deficiency
4.3.1. Understanting the Iron Deficiency
4.3.2. The Martial Status Biomarkers during Iron Deficiency
- Iron storage depletion. During the first phase of iron depletion, the deposits in the bone marrow, liver, and spleen are becoming exhausted (no stainable bone marrow iron), but no consequences on erythropoiesis are detectable yet. This early depletion is characterized by low ferritin (<35 μg/L), but normal Hb and other martial status indices [36]. The bone marrow is a major site for iron storage, but all the local metal is used for erythropoiesis, easily impairing RBC generation upon iron depletion at this site. The absence of stainable iron in the bone marrow is the gold standard for iron deficiency diagnosis, but it is used only in certain circumstances due to the invasive nature of the procedure [49]. It is based on the Prussian blue staining of aspirates to detect both hemosiderin in macrophages and iron granules in sideroblasts. The analysis requires an experienced observer and careful attention to detail [50]. The serum fraction of ferritin represents a portion of the total body pool that is stored in cells specialized in storing the metal and processing heme (e.g., hepatocytes and macrophages). In healthy individuals, the normal concentrations range between 15 and 300 μg/L, with lower values in children vs. adults, in women vs. men, and in fertile vs. post-menopausal women. Normally, 1 μg/L of serum ferritin corresponds to 8–10 mg of stored iron as a direct proportion. Values comprised between 12 and 15 μg/L indicate a depletion of iron stores. The ferritin measurement is widely available, standardized, and methodologically robust, and is based on colorimetric/fluorescent enzyme-linked immunoassays (ELISA) or on chemiluminescent immunoassays (CLIA) ran on automated analyzers [51]. The serum is the best matrix for a proper ferritin measurement, although plasma is also suitable depending on the analytical method.
- Iron supply discrepancies. In the second stage of deficient erythropoiesis, the decreased rate is ascribed to inadequate iron supply to the bone marrow. While Hb has still normal values (>115g/L), ferritin further reduces (<20 μg/L) together with Tf saturation (<16%). Contrariwise, there is an increase of the sTfR (>1.75 mg/dL) [36]. When the functional requirements are not met by dietary absorption or storage release, serum iron (i.e., the amount of Fe3+ in the blood bound to Tf) decreases while Tf increases. Because of this liaison, three assays that measure the potential of iron supply are generally performed concomitantly, being the serum iron, the Tf concentration (reported as the quantity of iron that can be bound to Tf = total iron binding capacity, TIBC), and the percentage of Tf saturation (serum iron × 100/TIBC) [52]. Serum iron can be measured by either colorimetric assays (most used) or atomic absorption spectrophotometry [53]. The concentration of serum transferrin can be measured by immunologic methods (direct) or throughout the determination of TIBC, whose assay is identical to the serum iron assay, but applies an additional step (saturation of iron-binding sites of the transferrin molecule with excess iron) followed by the removal of the unbound iron. Several analyzers measure also the unsaturated iron binding capacity (UIBC), with TIBC being subsequently calculated by summing UIBC to serum iron [54]. Serum iron, TIBC, and transferrin saturation are indexes of an adequate iron supply, but their utility as screening tools for iron deficiency is limited by several factors, such as the circadian rhythm (e.g., morning peak of serum iron and Tf saturation), diet, and oral contraceptive use [55]. Nevertheless, a Tf saturation < 16% is known to reflect a suboptimal iron supply for the proper erythrocyte development [52]. Normal values of serum iron range between 65 μg/dL to 170 μg/dL in adult males and 50 μg/dL to 170 μg/dL in adult females. TIBC and Tf saturation normal ranges are 250–450 μg/dL and 20–60%, respectively, in both adult males and females [48]. The serum is the best sample matrix, but also heparin-plasma may be used, whilst EDTA- and citrate-plasma are unsuitable due to the chelating properties of these anticoagulants. Cellular ion demands [56], the erythroid proliferation rate [57], and the stainable bone marrow iron [58] are known to be linked to the concentrations of the soluble form of the serine protease-cleaved membrane receptor (sTfR) that circulates in plasma bound to Tf. Several lifestyle factors affect sTfR, such as smoking, alcoholic drinking, sedentary behaviors, and hypernutrition [36]. Latex-enhanced immunoassays (nephelometry and turbidimetry) and the more recent immunofluorometric assays have been implemented to evaluate sTfR. However, the usefulness of commercial kits is limited by the poor comparability between different tools, comprising the calibrators (free vs. transferrin-complexed form, tissue origin), the antibodies (monoclonal vs. polyclonal), and reporting units (mg/L vs. nmol/L) [59]. This lack of commutability together with the relatively high cost of reagents are some of the reasons why sTfR measurements have not been widely adopted in clinical practice. Normal range of sTfR are 0.30–1.75 mg/dL. The serum is the best matrix and it should be separated within 8 h from blood drawings in order to get reliable results [48]. Of note, the sTfR/serum ferritin ratio may be more reliable than each parameter alone for the identification of iron deficiency [60].
- Iron-deficient anemia. The third stage of iron-deficient anemia is characterized by a reduction of both Hb concentrations and RBCs below-optimal levels (i.e., functional iron deficiency = iron supply is inadequate to meet the requirements for erythropoiesis). In the absence of ongoing inflammatory processes, the biochemical features are low ferritin (<12 μg/L), Tf saturation (<16%), and Hb (<115 g/L), but high sTfR (>1.75 mg/dL) and RBC protoporphyrin (>80 μg/dL). During the ferrochelatase-dependent insertion of ferrous iron in the proto-porphyrin ring, zinc can alternatively be incorporated to form zinc protoporphyrin, which is normally found in trace amounts [61]. In the early stages of reduced erythropoiesis, erythrocyte zinc protoporphyrin progressively rises, thereby providing to be a useful parameter for detecting uncomplicated functional iron deficiency. Importantly, its measure represents the average iron availability for erythropoiesis during the preceding 3–4 months since they are established during erythrocyte maturation and remain unaltered for the mature RBC lifespan. This value can be measured directly by hematofluorometer (porphyrins fluoresce in the red wavelengths when opportunely excited) or after extraction of the zinc moiety using ethyl acetate and hydrochloric acid. In this latter case, the zinc-free erythrocyte protoporphyrin is measured by conventional fluorometry. Values > 150 μmol/mol heme are highly suggestive of iron deficiency [62]. Although RBCs represent the largest functional compartment, their associated indices are not representative of the individual’s martial status. Hb concentration is usually relevant for assessing the degree of severity of iron deficiency, but its sensitivity is low because of the rather inconsistent variations between healthy and iron-deficient individuals. In addition, the specificity of this test is poor. The packed cell volume (hematocrit, Hct), although widely used in the past, does not provide any additional information to Hb concentration. Altered RBC indices, meaning a reduction of mean corpuscular volume (MCV), a reduction of mean corpuscular hemoglobin (MCH), and an increase of red blood cell distribution width (RDW), are usually a feature of iron-deficient erythropoiesis, but they lack specificity [36,48]. Conversely, modern analyzers can measure reticulocyte and hypochromic cell parameters, such as the reticulocyte Hb and the proportion of hypochromic erythrocytes, which may be useful for a proper assessment of anemia in chronic conditions characterized by a generalized inflammatory state. For instance, the biochemical feature of functional iron deficiency in chronic heart failure can show normal Hb values [63] and higher cut-off limits for both Tf saturation (<20%) and ferritin (<300 μg/L) [64]. Heightened values of ferritin may be also found in chronic kidney disease patients, where the concomitant proteinuria, low-iron diet, and inflammation expose them to veiled iron-deficient conditions [65]. The proportion of hypochromic erythrocytes with the reticulocyte Hb count could be used in these cases though, also for predicting the responsiveness to iron therapy [66].
5. The Present of Iron Medicine
5.1. Iron Foods
5.2. Dietary Patterns
5.3. Fortified Foods
5.4. Hands-On Approaches
5.5. Dietary Supplements
5.6. Parenteral Routes: Transfusions and Injections
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- History.com. Iron Age: A&E Television Networks. 2020. Available online: https://www.history.com/topics/pre-history/iron-age (accessed on 1 May 2020).
- Dickinson, O. The Aegean from Bronze Age to Iron Age: Continuity and Change between the Twelfth and Eighth Centuries BC.; Routledge: London, UK; New York, NY, USA, 2007. [Google Scholar]
- RSC. Iron: The Royal Society of Chemistry. 2020. Available online: https://www.rsc.org/periodic-table/element/26/iron (accessed on 1 May 2020).
- NASA. Mars: The National Aeronautics and Space Administration. 2020. Available online: https://solarsystem.nasa.gov/planets/mars/overview/ (accessed on 1 May 2020).
- Stearn, W.T. The Origin of the Male and Female Symbols of Biology. Taxon 1962, 11, XI. [Google Scholar] [CrossRef]
- Römheld, V. Different strategies for iron acquisition in higher plants. Physiol. Plant. 1987, 70, 231–234. [Google Scholar] [CrossRef]
- Ross, A.C.; Caballero, B.; Cousins, R.J.; Tucker, K.L.; Ziegler, T.R. Modern Nutrition in Health and Disease, 11th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2014. [Google Scholar]
- Gyana, R.; Rout Sahoo, S. Role of iron in plant growth and metabolism. Rev. Agric. Sci. 2015, 3, 1–24. [Google Scholar]
- Stevens, G.A.; Finucane, M.M.; De-Regil, L.M.; Paciorek, C.J.; Flaxman, S.R.; Branca, F.; Peña-Rosas, J.P.; Bhutta, Z.A.; Ezzati, M.; on behalf of Nutrition Impact Model Study Group (Anaemia). Global, regional, and national trends in haemoglobin concentration and prevalence of total and severe anaemia in children and pregnant and non-pregnant women for 1995–2011, a systematic analysis of population-representative data. Lancet Glob Health 2013, 1, e16–e25. [Google Scholar] [CrossRef] [Green Version]
- Camaschella, C. Iron-deficiency anemia. N. Engl. J. Med. 2015, 372, 1832–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chappell, C. Grandma’s Remedies: A Guide to Traditional Cures and Treatments from Mustard Poultices to Rosehip Syrup: Cornerstone Digital; Penguin Random House: London, UK, 2009. [Google Scholar]
- Arienti, G. Le Basi Molecolari della Nutrizione, 3rd ed.; Piccin Nuova Libraria: Padova, Italy, 2011. [Google Scholar]
- Green, J. Joey Green’s Magic Health Remedies: 1363 Quick-and-Easy Cures Using Brand-Name Products, 4th ed.; Rodale Press | Penguin Random House: London, UK, 2013. [Google Scholar]
- Jain, R.; Venkatasubramanian, P. Sugarcane Molasses—A Potential Dietary Supplement in the Management of Iron Deficiency Anemia. J. Diet. Suppl. 2017, 14, 589–598. [Google Scholar] [CrossRef]
- Anderson, G.J.; Frazer, D.M. Current understanding of iron homeostasis. Am. J. Clin. Nutr. 2017, 106 (Suppl. 6), 1559S–1566S. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, J. Mechanisms of cellular iron acquisition: Another iron in the fire. Cell 2002, 111, 603–606. [Google Scholar] [CrossRef] [Green Version]
- Gulec, S.; Anderson, G.J.; Collins, J.F. Mechanistic and regulatory aspects of intestinal iron absorption. Am. J. Physiol. Gastrointest Liver Physiol. 2014, 307, G397–G409. [Google Scholar] [CrossRef] [Green Version]
- Le Blanc, S.; Garrick, M.D.; Arredondo, M. Heme carrier protein 1 transports heme and is involved in heme-Fe metabolism. Am. J. Physiol. Cell Physiol. 2012, 302, C1780–C1785. [Google Scholar] [CrossRef]
- Kalgaonkar, S.; Lonnerdal, B. Receptor-mediated uptake of ferritin-bound iron by human intestinal Caco-2 cells. J. Nutr. Biochem. 2009, 20, 304–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Attieh, Z.K.; Su, T.; Syed, B.A.; Gao, H.; Alaeddine, R.M.; Fox, T.C.; Usta, J.; Naylor, C.E.; Evans, R.W.; et al. Hephaestin is a ferroxidase that maintains partial activity in sex-linked anemia mice. Blood 2004, 103, 3933–3939. [Google Scholar] [CrossRef] [PubMed]
- Dautry-Varsat, A. Receptor-mediated endocytosis: The intracellular journey of transferrin and its receptor. Biochimie 1986, 68, 375–381. [Google Scholar] [CrossRef]
- Luck, A.N.; Mason, A.B. Transferrin-mediated cellular iron delivery. Curr. Top. Membr. 2012, 69, 3–35. [Google Scholar] [PubMed] [Green Version]
- Grant, B.D.; Donaldson, J.G. Pathways and mechanisms of endocytic recycling. Nat. Rev. Mol. Cell Biol. 2009, 10, 597–608. [Google Scholar] [CrossRef] [Green Version]
- Abbaspour, N.; Hurrell, R.; Kelishadi, R. Review on iron and its importance for human health. J. Res. Med. Sci. 2014, 19, 164–174. [Google Scholar]
- Nagelkerke, S.Q.; Bruggeman, C.W.; den Haan, J.M.M.; Mul, E.P.J.; van den Berg, T.K.; van Bruggen, R.; Kuijpers, T.W. Red pulp macrophages in the human spleen are a distinct cell population with a unique expression of Fc-gamma receptors. Blood Adv. 2018, 2, 941–953. [Google Scholar] [CrossRef] [Green Version]
- Ganz, T. Macrophages and systemic iron homeostasis. J. Innate Immun. 2012, 4, 446–453. [Google Scholar] [CrossRef]
- Nemeth, E.; Ganz, T. Regulation of iron metabolism by hepcidin. Annu. Rev. Nutr. 2006, 26, 323–342. [Google Scholar] [CrossRef]
- Nunez, M.T. Regulatory mechanisms of intestinal iron absorption-uncovering of a fast-response mechanism based on DMT1 and ferroportin endocytosis. Biofactors 2010, 36, 88–97. [Google Scholar]
- De Domenico, I.; Ward, D.M.; Kaplan, J. Hepcidin regulation: Ironing out the details. J. Clin. Investig. 2007, 117, 1755–1758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, G.J. Mechanisms of iron loading and toxicity. Am. J. Hematol. 2007, 82 (Suppl. 12), 1128–1131. [Google Scholar] [CrossRef] [PubMed]
- Dumic, I.; Nordin, T.; Jecmenica, M.; Stojkovic Lalosevic, M.; Milosavljevic, T.; Milovanovic, T. Gastrointestinal Tract Disorders in Older Age. Can. J. Gastroenterol. Hepatol. 2019, 2019, 6757524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annibale, B.; Capurso, G.; Martino, G.; Grossi, C.; Delle Fave, G. Iron deficiency anaemia and Helicobacter pylori infection. Int. J. Antimicrob. Agents 2000, 16, 515–519. [Google Scholar] [CrossRef]
- Chasteen, N.D.; Harrison, P.M. Mineralization in ferritin: An efficient means of iron storage. J. Struct. Biol. 1999, 126, 182–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lonnerdal, B. Calcium and iron absorption--mechanisms and public health relevance. Int. J. Vitam. Nutr. Res. 2010, 80, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Suchdev, P.S.; Namaste, S.M.; Aaron, G.J.; Raiten, D.J.; Brown, K.H.; Flores-Ayala, R.; the BRINDA Working Group. Overview of the Biomarkers Reflecting Inflammation and Nutritional Determinants of Anemia (BRINDA) Project. Adv. Nutr. 2016, 7, 349–356. [Google Scholar] [CrossRef] [Green Version]
- Lynch, S.; Pfeiffer, C.M.; Georgieff, M.K.; Brittenham, G.; Fairweather-Tait, S.; Hurrell, R.F.; McArdle, H.J.; Raiten, D.J. Biomarkers of Nutrition for Development (BOND)-Iron Review. J. Nutr. 2018, 148 (Suppl. 1), 1001S–1067S. [Google Scholar] [CrossRef] [Green Version]
- WHO. Iron Deficiency Anaemia—Assessment, Prevention and Control. In A Guide for Programme Managers; WHO/NHD/01.3; WHO Publications: Geneva, Switzerland, 2001. [Google Scholar]
- Munoz, M.; Villar, I.; Garcia-Erce, J.A. An update on iron physiology. World J. Gastroenterol. 2009, 15, 4617–4626. [Google Scholar] [CrossRef]
- Joint WHO/CDC Technical Consultation on the Assessment of Iron Status at the Population Level. In Proceedings of the Assessing the Iron Status of Populations: Including Literature Reviews: Report of a Joint World Health Organization/Centers for Disease Control and Prevention Technical Consultation on the Assessment of Iron Status at the Population Level; Geneva, Switzerland, 6–8 April 2004.
- Harms, K.; Kaiser, T. Beyond soluble transferrin receptor: Old challenges and new horizons. Best Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 799–810. [Google Scholar] [CrossRef]
- Shih, Y.J.; Baynes, R.D.; Hudson, B.G.; Flowers, C.H.; Skikne, B.S.; Cook, J.D. Serum transferrin receptor is a truncated form of tissue receptor. J. Biol. Chem. 1990, 265, 19077–19081. [Google Scholar] [PubMed]
- Worwood, M. Indicators of the Iron Status of Populations: Ferritin. Reference Manual for Laboratory Considerations—Iron Status Indicators for Population Assessments; WHO: Geneva, Switzerland, 2007. [Google Scholar]
- Thurnham, D.I.; McCabe, L.D.; Haldar, S.; Wieringa, F.T.; Northrop-Clewes, C.A.; McCabe, G.P. Adjusting plasma ferritin concentrations to remove the effects of subclinical inflammation in the assessment of iron deficiency: A meta-analysis. Am. J. Clin. Nutr. 2010, 92, 546–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomkins, A. Assessing micronutrient status in the presence of inflammation. J. Nutr. 2003, 133 (Suppl. 5), 1649S–1655S. [Google Scholar] [CrossRef] [PubMed]
- Dignass, A.; Farrag, K.; Stein, J. Limitations of Serum Ferritin in Diagnosing Iron Deficiency in Inflammatory Conditions. Int. J. Chronic Dis. 2018, 2018, 9394060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munoz, M.; Acheson, A.G.; Auerbach, M.; Besser, M.; Habler, O.; Kehlet, H.; Liumbruno, G.M.; Lasocki, S.; Meybohm, P.; Baikady, R.R.; et al. International consensus statement on the peri-operative management of anaemia and iron deficiency. Anaesthesia 2017, 72, 233–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briguglio, M.; Gianola, S.; Aguirre, M.F.I.; Sirtori, P.; Perazzo, P.; Pennestri, F.; Brayda-Bruno, M.; Sansone, V.; Banfi, G. Nutritional support for enhanced recovery programs in orthopedics: Future perspectives for implementing clinical practice. Nutr. Clin. Metab. 2019, 33, 190–198. [Google Scholar] [CrossRef]
- Lombardi, G.; Lippi, G.; Banfi, G. Iron requirements and iron status of athletes. In Encyclopaedia of Sports Medicine XIX.; Maughan, R.J., Ed.; John Wiley & Sons: Oxford, UK, 2014; pp. 229–241. [Google Scholar]
- Gale, E.; Torrance, J.; Bothwell, T. The quantitative estimation of total iron stores in human bone marrow. J. Clin. Investig. 1963, 42, 1076–1082. [Google Scholar] [CrossRef] [PubMed]
- Barron, B.A.; Hoyer, J.D.; Tefferi, A. A bone marrow report of absent stainable iron is not diagnostic of iron deficiency. Ann. Hematol. 2001, 80, 166–169. [Google Scholar] [CrossRef]
- Zimmermann, M.B. Methods to assess iron and iodine status. Br. J. Nutr. 2008, 99 (Suppl. 3), S2–S9. [Google Scholar] [CrossRef]
- Beard, J. Indicators of the iron status of populations: Free erythrocyte protoporphyrin and zinc protoporphyrin; serum and plasma iron, total iron binding capacity and transferrin saturation; and serum transferrin receptor. In Reference Manual for Laboratory Considerations—Iron Status Indicators for Population Assessments; WHO: Geneva, Switzerland, 2007. [Google Scholar]
- Hedayati, M.; Abubaker-Sharif, B.; Khattab, M.; Razavi, A.; Mohammed, I.; Nejad, A.; Wabler, M.; Zhou, H.; Mihalic, J.; Gruettner, C.; et al. An optimised spectrophotometric assay for convenient and accurate quantitation of intracellular iron from iron oxide nanoparticles. Int. J. Hyperther. 2018, 34, 373–381. [Google Scholar] [CrossRef]
- Adams, P.C.; Reboussin, D.M.; Leiendecker-Foster, C.; Moses, G.C.; McLaren, G.D.; McLaren, C.E.; Dawkins, F.W.; Kasvosve, I.; Acton, R.T.; Barton, J.C.; et al. Comparison of the unsaturated iron-binding capacity with transferrin saturation as a screening test to detect C282Y homozygotes for hemochromatosis in 101,168 participants in the hemochromatosis and iron overload screening (HEIRS) study. Clin. Chem. 2005, 51, 1048–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bullock, G.C.; Delehanty, L.L.; Talbot, A.L.; Gonias, S.L.; Tong, W.H.; Rouault, T.A.; Dewar, B.; Macdonald, J.M.; Chruma, J.J.; Goldfarb, A.N. Iron control of erythroid development by a novel aconitase-associated regulatory pathway. Blood 2010, 116, 97–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skikne, B.S.; Flowers, C.H.; Cook, J.D. Serum transferrin receptor: A quantitative measure of tissue iron deficiency. Blood 1990, 75, 1870–1876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohgo, Y.; Niitsu, Y.; Kondo, H.; Kato, J.; Tsushima, N.; Sasaki, K.; Hirayama, M.; Numata, T.; Nishisato, T.; Urushizaki, I. Serum transferrin receptor as a new index of erythropoiesis. Blood 1987, 70, 1955–1958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.; Bird, R.; Clague, A.; Carter, A. Clinical utility of serum soluble transferrin receptor levels and comparison with bone marrow iron stores as an index for iron-deficient erythropoiesis in a heterogeneous group of patients. Pathology 2007, 39, 349–353. [Google Scholar] [CrossRef]
- Speeckaert, M.M.; Speeckaert, R.; Delanghe, J.R. Biological and clinical aspects of soluble transferrin receptor. Crit. Rev. Clin. Lab. Sci. 2010, 47, 213–228. [Google Scholar] [CrossRef]
- Skikne, B.S.; Punnonen, K.; Caldron, P.H.; Bennett, M.T.; Rehu, M.; Gasior, G.H.; Chamberlin, J.S.; Sullivan, L.A.; Bray, K.R.; Southwick, P.C. Improved differential diagnosis of anemia of chronic disease and iron deficiency anemia: A prospective multicenter evaluation of soluble transferrin receptor and the sTfR/log ferritin index. Am. J. Hematol. 2011, 86, 923–927. [Google Scholar] [CrossRef]
- Labbe, R.F.; Vreman, H.J.; Stevenson, D.K. Zinc protoporphyrin: A metabolite with a mission. Clin. Chem. 1999, 45, 2060–2072. [Google Scholar] [CrossRef] [Green Version]
- Tillyer, M.L.; Tillyer, C.R. Zinc protoporphyrin assays in patients with alpha and beta thalassaemia trait. J. Clin. Pathol. 1994, 47, 205–208. [Google Scholar] [CrossRef] [Green Version]
- Mordi, I.R.; Tee, A.; Lang, C.C. Iron Therapy in Heart Failure: Ready for Primetime? Card. Fail. Rev. 2018, 4, 28–32. [Google Scholar] [CrossRef]
- McDonagh, T.; Macdougall, I.C. Iron therapy for the treatment of iron deficiency in chronic heart failure: Intravenous or oral? Eur. J. Heart Fail. 2015, 17, 248–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalantar-Zadeh, K.; Kalantar-Zadeh, K.; Lee, G.H. The fascinating but deceptive ferritin: To measure it or not to measure it in chronic kidney disease? Clin. J. Am. Soc. Nephrol. 2006, 1 (Suppl. 1), S9–S18. [Google Scholar] [CrossRef] [PubMed]
- Ratcliffe, L.E.; Thomas, W.; Glen, J.; Padhi, S.; Pordes, B.A.; Wonderling, D.; Connell, R.; Stephens, S.; Mikhail, A.I.; Fogarty, D.G.; et al. Diagnosis and Management of Iron Deficiency in CKD: A Summary of the NICE Guideline Recommendations and Their Rationale. Am. J. Kidney Dis. 2016, 67, 548–558. [Google Scholar] [CrossRef]
- The National Heart Lung and Blood Institute (NHLBI). Iron-Deficiency Anemia. Available online: https://www.nhlbi.nih.gov/health-topics/iron-deficiency-anemia (accessed on 1 May 2020).
- WHO/FAO. Vitamin and Mineral Requirements in Human Nutrition; WHO: Geneva, Switzerland, 2004. [Google Scholar]
- EFSA NDA Panel (EFSA Panel on Dietetic Products Nutrition and Allergies). Scientific Opinion on Dietary Reference Values for Iron. EFSA J. 2015, 13, 4254. [Google Scholar] [CrossRef]
- Lombardi-Boccia, G.; Lanzi, S.; Lucarini, M.; Di Lullo, G. Meat and meat products consumption in Italy: Contribution to trace elements, heme iron and selected B vitamins supply. Int. J. Vitam. Nutr. Res. 2004, 74, 247–251. [Google Scholar] [CrossRef]
- Hunt, J.R. Bioavailability of iron, zinc, and other trace minerals from vegetarian diets. Am. J. Clin. Nutr. 2003, 78 (Suppl. 3), 633S–639S. [Google Scholar] [CrossRef]
- Souci, S.W.; Fachmann, W.; Kraut, H. Tabelle Complete degli Alimenti. In Edizione Italiana a Cura di M. Carruba; Mattioli: Fidenza, Italy, 1885. [Google Scholar]
- Società Italiana di Nutrizione Umana. LARN—Livelli di Assunzione di Riferimento di Nutrienti ed Energia per la Popolazione Italiana, IV ed.; SICS Editore Srl: Rome, Italy, 2014; pp. 482–502. [Google Scholar]
- Briguglio, M.; Dell’Osso, B.; Panzica, G.; Malgaroli, A.; Banfi, G.; Zanaboni Dina, C.; Galentino, R.; Porta, M. Dietary Neurotransmitters: A Narrative Review on Current Knowledge. Nutrients 2018, 10, 591. [Google Scholar] [CrossRef] [Green Version]
- Hurrell, R.; Egli, I. Iron bioavailability and dietary reference values. Am. J. Clin. Nutr. 2010, 91, 1461S–1467S. [Google Scholar] [CrossRef]
- Semba, R.D.; Bloem, M.W. The anemia of vitamin A deficiency: Epidemiology and pathogenesis. Eur. J. Clin. Nutr. 2002, 56, 271–281. [Google Scholar] [CrossRef] [Green Version]
- EFSA (European Food Safety Authority). Dietary Reference Values for Nutrients: Summary Report; EFSA Supporting Publication: Parma, Italy, 2017; Volume 98, p. e15121. [Google Scholar]
- Hurrell, R.; Bothwell, T.; Cook, J.D.; Dary, O.; Davidsson, L.; Fairweather-Tait, S.; Hallberg, L.; Lynch, S.; Rosado, J.; Walter, T.; et al. The usefulness of elemental iron for cereal flour fortification: A SUSTAIN Task Force report. Sharing United States Technology to Aid in the Improvement of Nutrition. Nutr. Rev. 2002, 60, 391–406. [Google Scholar] [CrossRef]
- Lynch, S.R.; Bothwell, T.; STFoI, P. A comparison of physical properties, screening procedures and a human efficacy trial for predicting the bioavailability of commercial elemental iron powders used for food fortification. Int. J. Vitam. Nutr. Res. 2007, 77, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.; Thankachan, P.; Muthayya, S.; Goud, R.B.; Kurpad, A.V.; Hurrell, R.F.; Zimmermann, M.B. Dual fortification of salt with iodine and iron: A randomized, double-blind, controlled trial of micronized ferric pyrophosphate and encapsulated ferrous fumarate in southern India. Am. J. Clin. Nutr. 2008, 88, 1378–1387. [Google Scholar]
- Henare, S.J.; Nur Singh, N.; Ellis, A.M.; Moughan, P.J.; Thompson, A.K.; Walczyk, T. Iron bioavailability of a casein-based iron fortificant compared with that of ferrous sulfate in whole milk: A randomized trial with a crossover design in adult women. Am. J. Clin. Nutr. 2019, 110, 1362–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, M.; Sharma, N.; Sharma, S.; Kapoor, P.; Kumar, A.; Chunduri, V.; Arora, P. Biofortified Crops Generated by Breeding, Agronomy, and Transgenic Approaches Are Improving Lives of Millions of People around the World. Front. Nutr. 2018, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Sperotto, R.A.; Ricachenevsky, F.K.; Waldow Vde, A.; Fett, J.P. Iron biofortification in rice: It’s a long way to the top. Plant Sci. 2012, 190, 24–39. [Google Scholar] [CrossRef]
- Connorton, J.M.; Balk, J. Iron Biofortification of Staple Crops: Lessons and Challenges in Plant Genetics. Plant Cell Physiol. 2019, 60, 1447–1456. [Google Scholar] [CrossRef] [Green Version]
- Finkelstein, J.L.; Fothergill, A.; Hackl, L.S.; Haas, J.D.; Mehta, S. Iron biofortification interventions to improve iron status and functional outcomes. Proc. Nutr. Soc. 2019, 78, 197–207. [Google Scholar] [CrossRef]
- Campos Ponce, M.; Polman, K.; Roos, N.; Wieringa, F.T.; Berger, J.; Doak, C.M. What Approaches are Most Effective at Addressing Micronutrient Deficiency in Children 0-5 Years? A Review of Systematic Reviews. Matern. Child Health J. 2019, 23 (Suppl. 1), 4–17. [Google Scholar] [CrossRef] [Green Version]
- Charles, C.V.; Dewey, C.E.; Daniell, W.E.; Summerlee, A.J. Iron-deficiency anaemia in rural Cambodia: Community trial of a novel iron supplementation technique. Eur. J. Public Health 2011, 21, 43–48. [Google Scholar] [CrossRef]
- Tara, M. Scrap Metal Pots an Unrecognized Source of Lead Poisoning; The Epoch Times: New York, NY, USA, 2017; Available online: https://www.theepochtimes.com/cooking-pots-made-from-recycled-metal-a-source-of-lead-poisoning_2215631.html (accessed on 1 May 2020).
- Casey, G.J.; Montresor, A.; Cavalli-Sforza, L.T.; Thu, H.; Phu, L.B.; Tinh, T.T.; Tien, N.T.; Phuc, T.Q.; Biggs, B.-A. Elimination of iron deficiency anemia and soil transmitted helminth infection: Evidence from a fifty-four month iron-folic acid and de-worming program. PLoS Negl. Trop. Dis. 2013, 7, e2146. [Google Scholar] [CrossRef] [Green Version]
- Cercamondi, C.I.; Egli, I.M.; Ahouandjinou, E.; Dossa, R.; Zeder, C.; Salami, L.; Tjalsma, H.; Wiegerinck, E.; Tanno, T.; Hurrell, R.F.; et al. Afebrile Plasmodium falciparum parasitemia decreases absorption of fortification iron but does not affect systemic iron utilization: A double stable-isotope study in young Beninese women. Am. J. Clin. Nutr. 2010, 92, 1385–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briguglio, M.; Pregliasco, F.E.; Lombardi, G.; Perazzo, P.; Banfi, G. The Malnutritional Status of the Host as a Virulence Factor for New Coronavirus SARS-CoV-2. Front. Med. 2020, 7, 146. [Google Scholar] [CrossRef] [PubMed]
- Troesch, B.; Jing, H.; Laillou, A.; Fowler, A. Absorption studies show that phytase from Aspergillus niger significantly increases iron and zinc bioavailability from phytate-rich foods. Food Nutr. Bull. 2013, 34 (Suppl. 2), S90–S101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jefferds, M.E.; Irizarry, L.; Timmer, A.; Tripp, K. UNICEF-CDC global assessment of home fortification interventions 2011, current status, new directions, and implications for policy and programmatic guidance. Food Nutr. Bull. 2013, 34, 434–443. [Google Scholar] [CrossRef] [Green Version]
- De-Regil, L.M.; Suchdev, P.S.; Vist, G.E.; Walleser, S.; Pena-Rosas, J.P. Home fortification of foods with multiple micronutrient powders for health and nutrition in children under two years of age (Review). Evid. Based Child Health 2013, 8, 112–201. [Google Scholar] [CrossRef]
- De-Regil, L.M.; Jefferds, M.E.D.; Pena-Rosas, J.P. Point-of-use fortification of foods with micronutrient powders containing iron in children of preschool and school-age. Cochrane Database Syst. Rev. 2017, 11, CD009666. [Google Scholar] [CrossRef]
- Jaeggi, T.; Kortman, G.A.; Moretti, D.; Chassard, C.; Holding, P.; Dostal, A.; Boekhorst, J.; Timmerman, H.M.; Swinkels, D.W.; Tjalsma, H.; et al. Iron fortification adversely affects the gut microbiome, increases pathogen abundance and induces intestinal inflammation in Kenyan infants. Gut 2015, 64, 731–742. [Google Scholar] [CrossRef]
- Sazawal, S.; Black, R.E.; Ramsan, M.; Chwaya, H.M.; Stoltzfus, R.J.; Dutta, A.; Dhingra, U.; Kabole, I.; Deb, S.; Othmanet, M.K.; et al. Effects of routine prophylactic supplementation with iron and folic acid on admission to hospital and mortality in preschool children in a high malaria transmission setting: Community-based, randomised, placebo-controlled trial. Lance 2006, 367, 133–143. [Google Scholar] [CrossRef]
- Harrington, M.; Hotz, C.; Zeder, C.; Polvo, G.O.; Villalpando, S.; Zimmermann, M.B.; Walczyk, T.; Rivera, J.A.; Hurrell, R.F. A comparison of the bioavailability of ferrous fumarate and ferrous sulfate in non-anemic Mexican women and children consuming a sweetened maize and milk drink. Eur. J. Clin. Nutr. 2011, 65, 20–25. [Google Scholar] [CrossRef] [Green Version]
- Briguglio, M.; Hrelia, S.; Malaguti, M.; De Vecchi, E.; Lombardi, G.; Banfi, G.; Riso, P.; Porrini, M.; Romagnoli, S.; Pino, F.; et al. Oral Supplementation with Sucrosomial Ferric Pyrophosphate Plus L-Ascorbic Acid to Ameliorate the martial Status: A Randomized Controlled Trial. Nutrients 2020, 12, 386. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, R.; Toblli, J.E.; Romero, J.D.; Monterrosa, B.; Frer, C.; Macagno, E.; Breymann, C. Efficacy and safety of oral iron(III) polymaltose complex versus ferrous sulfate in pregnant women with iron-deficiency anemia: A multicenter, randomized, controlled study. J. Matern. Fetal. Neonatal. Med. 2011, 24, 1347–1352. [Google Scholar] [CrossRef] [PubMed]
- Asperti, M.; Gryzik, M.; Brilli, E.; Castagna, A.; Corbella, M.; Gottardo, R.; Girelli, D.; Tarantino, G.; Arosio, P.; Poli, M. Sucrosomial((R)) Iron Supplementation in Mice: Effects on Blood Parameters, Hepcidin, and Inflammation. Nutrients 2018, 10, 1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briguglio, M.; Dell’Osso, B.; Galentino, R.; Zanaboni Dina, C.; Banfi, G.; Porta, M. Tics and obsessive-compulsive disorder in relation to diet: Two case reports. Encephale 2018, 44, 479–481. [Google Scholar] [CrossRef] [PubMed]
- Briguglio, M.; Hrelia, S.; Malaguti, M.; Serpe, L.; Canaparo, R.; Dell’Osso, B.; Galentino, R.; De Michele, S.; Zanaboni Dina, C.; Porta, M. Food Bioactive Compounds and Their Interference in Drug Pharmacokinetic/Pharmacodynamic Profiles. Pharmaceutics 2018, 10, 277. [Google Scholar] [CrossRef] [Green Version]
- Maluf, N.S.R. History of Blood Transfusion: The Use of Blood from Antiquity through the Eighteenth Century. J. Hist. Med. Allied Sci. 1954, 9, 59–107. [Google Scholar] [CrossRef] [PubMed]
- Winkelmayer, W.C.; Goldstein, B.A.; Mitani, A.A.; Ding, V.Y.; Airy, M.; Mandayam, S.; Chang, T.I.; Brookhart, M.A.; Fishbane, S. Safety of Intravenous Iron in Hemodialysis: Longer-term Comparisons of Iron Sucrose Versus Sodium Ferric Gluconate Complex. Am. J. Kidney Dis. 2017, 69, 771–779. [Google Scholar] [CrossRef]
- Nikravesh, N.; Borchard, G.; Hofmann, H.; Philipp, E.; Fluhmann, B.; Wick, P. Factors influencing safety and efficacy of intravenous iron-carbohydrate nanomedicines: From production to clinical practice. Nanomedicine 2020, 26, 102178. [Google Scholar] [CrossRef]
- Neogi, S.B.; Devasenapathy, N.; Singh, R.; Bhushan, H.; Shah, D.; Divakar, H.; Zodpey, S.; Malik, S.; Nanda, S.; Mittal, P.; et al. Safety and effectiveness of intravenous iron sucrose versus standard oral iron therapy in pregnant women with moderate-to-severe anaemia in India: A multicentre, open-label, phase 3, randomised, controlled trial. Lancet Glob Health 2019, 7, e1706–e1716. [Google Scholar] [CrossRef] [Green Version]
- Del Vecchio, L.; Longhi, S.; Locatelli, F. Safety concerns about intravenous iron therapy in patients with chronic kidney disease. Clin. Kidney J. 2016, 9, 260–267. [Google Scholar] [CrossRef] [Green Version]
- Auerbach, M.; Macdougall, I. The available intravenous iron formulations: History, efficacy, and toxicology. Hemodial. Int. 2017, 21 (Suppl. 1), S83–S92. [Google Scholar] [CrossRef] [Green Version]
- New recommendations to manage risk of allergic reactions with intravenous iron-containing medicines. In European Medicines Agency’s Committee for Medicinal Products for Human Use (CHMP); Report No.: EMA/579491/2013; 2013; Available online: https://www.ema.europa.eu/en/news/new-recommendations-manage-risk-allergic-reactions-intravenous-iron-containing-medicines (accessed on 11 June 2020).
- Grzywacz, A.; Lubas, A.; Fiedor, P.; Fiedor, M.; Niemczyk, S. Safety and Efficacy of Intravenous Administration of Iron Preparations. Acta Pol. Pharm. 2017, 74, 13–24. [Google Scholar] [PubMed]
- Janbek, J.; Sarki, M.; Specht, I.O.; Heitmann, B.L. A systematic literature review of the relation between iron status/anemia in pregnancy and offspring neurodevelopment. Eur. J. Clin. Nutr. 2019, 73, 1561–1578. [Google Scholar] [CrossRef] [PubMed]
- Mattiello, V.; Schmugge, M.; Hengartner, H.; von der Weid, N.; Renella, R.; Group SPHW. Diagnosis and management of iron deficiency in children with or without anemia: Consensus recommendations of the SPOG Pediatric Hematology Working Group. Eur. J. Pediatr. 2020, 179, 527–545. [Google Scholar] [CrossRef] [PubMed]
- Vetrano, D.L.; Zucchelli, A.; Marconi, E.; Levi, M.; Pegoraro, V.; Cataldo, N.; Heiman, G.; Cricelli, C.; Lapi, F. Predictors of iron-deficiency anemia in primary care older adults: A real-world European multi-country longitudinal study. Aging Clin. Exp. Res. 2020. [Google Scholar] [CrossRef]
- Briguglio, M.; Gianturco, L.; Stella, D.; Colombo, C.; Bonadies, M.; Sala, O.; Anselmi, M.; Banfi, G.; Turiel, M. Correction of hypovitaminosis D improved global longitudinal strain earlier than left ventricular ejection fraction in cardiovascular older adults after orthopaedic surgery. J. Geriatr. Cardiol. 2018, 15, 519–522. [Google Scholar]
- Briguglio, M.; Dell’Osso, B.; Galentino, R.; Banfi, G.; Porta, M. Higher adherence to the Mediterranean diet is associated with reduced tics and obsessive-compulsive symptoms: A series of nine boys with Obsessive-Compulsive Tic Disorder. Nutr. Clin. Metab. 2019, 33, 227–230. [Google Scholar] [CrossRef]
- Briguglio, M.; Vitale, J.A.; Galentino, R.; Banfi, G.; Zanaboni Dina, C.; Bona, A.; Panzica, G.; Porta, M.; Dell’Osso, B.; Glick, I.D. Healthy Eating, Physical Activity, and Sleep Hygiene (HEPAS) as the Winning Triad for Sustaining Physical and Mental Health in Patients at Risk for or with Neuropsychiatric Disorders: Considerations for Clinical Practice. Neuropsychiatr. Dis. Treat. 2020, 16, 55–70. [Google Scholar] [CrossRef] [Green Version]
- Mistry, R.; Hosoya, H.; Kohut, A.; Ford, P. Iron deficiency in heart failure, an underdiagnosed and undertreated condition during hospitalization. Ann. Hematol. 2019, 98, 2293–2297. [Google Scholar] [CrossRef]
- Gebremedhin, S.; Samuel, A.; Mamo, G.; Moges, T.; Assefa, T. Coverage, compliance and factors associated with utilization of iron supplementation during pregnancy in eight rural districts of Ethiopia: A cross-sectional study. BMC Public Health 2014, 14, 607. [Google Scholar] [CrossRef] [Green Version]
- Camaschella, C. Iron-Deficiency Anemia. N. Engl. J. Med. 2015, 373, 485–486. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Dietary Source | Average Contents of Iron, mg/100g Step |
|---|---|
| (Daily Needs for Adults: 10–11 mg) | |
| Animal foods | |
| Veal and other mammal liver, raw | 20 |
| Yolk of chicken eggs, raw | 5 |
| Fishes, raw | 5 |
| Meats (veal, beef), raw | 4 |
| Milk (cow), whole | 0.2 |
| Plant foods | |
| Common oregano, dried | 18 |
| Bitter cocoa, powder | 14.3 |
| Arabica coffee, powder | 12 |
| Dried pulses (lentils, beans), dried | 9 |
| Wheat bran, soy flour, dried | 8 |
| Walnuts, almonds, pistachios, dried | 7 |
| Edible mushrooms, raw | 1–2 |
| Red wine | 0.9–1.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Briguglio, M.; Hrelia, S.; Malaguti, M.; Lombardi, G.; Riso, P.; Porrini, M.; Perazzo, P.; Banfi, G. The Central Role of Iron in Human Nutrition: From Folk to Contemporary Medicine. Nutrients 2020, 12, 1761. https://doi.org/10.3390/nu12061761
Briguglio M, Hrelia S, Malaguti M, Lombardi G, Riso P, Porrini M, Perazzo P, Banfi G. The Central Role of Iron in Human Nutrition: From Folk to Contemporary Medicine. Nutrients. 2020; 12(6):1761. https://doi.org/10.3390/nu12061761
Chicago/Turabian StyleBriguglio, Matteo, Silvana Hrelia, Marco Malaguti, Giovanni Lombardi, Patrizia Riso, Marisa Porrini, Paolo Perazzo, and Giuseppe Banfi. 2020. "The Central Role of Iron in Human Nutrition: From Folk to Contemporary Medicine" Nutrients 12, no. 6: 1761. https://doi.org/10.3390/nu12061761
APA StyleBriguglio, M., Hrelia, S., Malaguti, M., Lombardi, G., Riso, P., Porrini, M., Perazzo, P., & Banfi, G. (2020). The Central Role of Iron in Human Nutrition: From Folk to Contemporary Medicine. Nutrients, 12(6), 1761. https://doi.org/10.3390/nu12061761