Total Dietary Antioxidant Intake Including Polyphenol Content: Is It Capable to Fight against Increased Oxidants within the Body of Ultra-Endurance Athletes?

 ,
, 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Study Design
2.3. Dietary Records Assessment and Dietary Total Antioxidant Capacity Analysis
2.4. Maximal Oxygen uptake (VO2max) Measurement
2.5. Exercise Protocol
2.6. Exercise Performance Measurement
2.7. Plasma Total Oxidant Status (TOS) and Plasma Total Antioxidant Status (TAS) Analysis
Oxidative Stress Index (OSI) Calculation
2.8. Plasma 8-isoprostaglandin F2alpha (8-iso PGF2a) Analysis
2.9. Lactate Analysis
2.10. Statistical Analysis
3. Results
3.1. Subjects
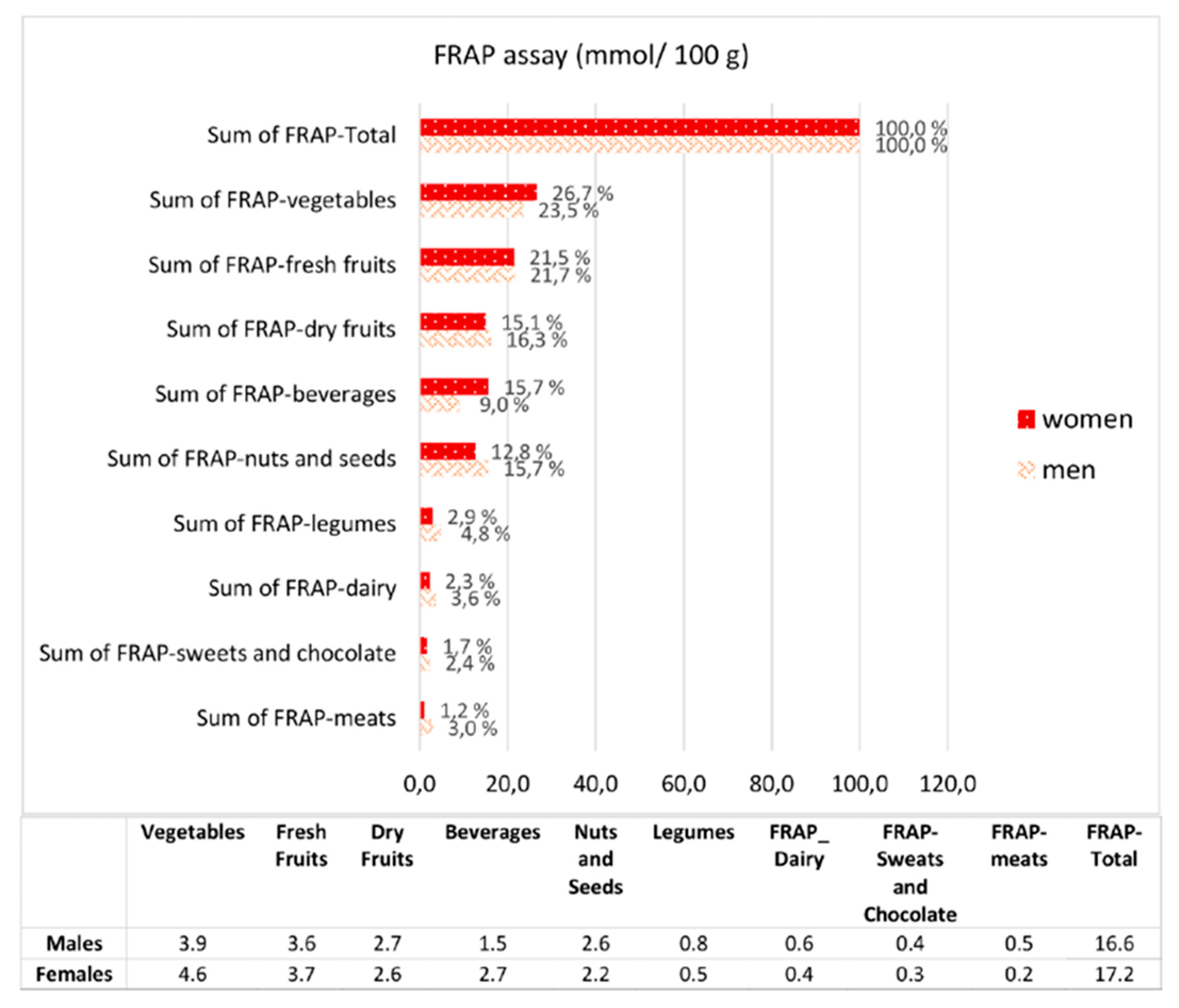
3.2. Dietary Antioxidant Intake
3.3. Changes in Plasma Oxidative Stress Biomarkers after Exercise Testing
3.4. Dietary Antioxidants and Exercise Performance
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Laursen, P.B. Long distance triathlon: Demands, preparation and performance. J. Hum. Sport Exerc. 2011, 6, 247–263. [Google Scholar] [CrossRef] [Green Version]
- Knechtle, B.; Nikolaidis, P.T. Physiology and pathophysiology in ultra-marathon running. Front. Physiol. 2018, 9, 634–667. [Google Scholar] [CrossRef] [Green Version]
- Sachdev, S.; Davies, K.J.A. Production, detection, and adaptive responses to free radicals in exercise. Free Radic. Biol. Med. 2008, 44, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, V.H.; Valente, H.F.; Casal, S.I.; Marques, A.F.; Moreira, P.A. Antioxidants do not prevent postexercise peroxidation and may delay muscle recovery. Med. Sci. Sports Exerc. 2009, 41, 1752–1760. [Google Scholar] [CrossRef] [PubMed]
- Nocella, C.; Cammisotto, V.; Pigozzi, F.; Borrione, P.; Fossati, C.; D’amico, A.; Cangemi, R.; Peruzzi, M.; Gobbi, G.; Ettorre, E.; et al. Impairment between oxidant and antioxidant systems: Short- and Long-Term implications for athletes’ health. Nutrients 2019, 11, 1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.W.; Kao, H.H.; Wu, C.H. Exercise training upregulates SIRT1 to attenuate inflammation and metabolic dysfunction in kidney and liver of diabetic db/db mice. Nutr. Metab. 2019, 16. [Google Scholar] [CrossRef]
- Pyne, D.B. Exercise-induced muscle damage and inflammation: A review. Aust. J. Sci. Med. Sport 1994, 26, 49–58. [Google Scholar] [PubMed]
- Powers, S.K.; Jackson, M.J. Exercise-induced oxidative stress: Cellular mechanisms and impact on muscle force production. Physiol. Rev. 2008, 88, 1243–1276. [Google Scholar] [CrossRef] [Green Version]
- Barclay, J.K.; Hansel, M. Free radicals may contribute to oxidative skeletal muscle fatigue. Can. J. Physiol. Pharm. 1991, 69, 279–284. [Google Scholar] [CrossRef]
- Reichhold, S.; Neubauer, O.; Bulmer, A.C.; Knasmü, S.; Wagner, K.-H. Endurance exercise and DNA stability: Is there a link to duration and intensity? Mutat. Res. 2009, 682, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Alessio, H.M.; Goldfarb, A.H.; Cao, G. Exercise-induced oxidative stress before and after vitamin C supplementation. Int. J. Sport Nutr. Exerc. Metab. 1997, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- McLeay, Y.; Stannard, S.; Houltham, S.; Starck, C. Dietary thiols in exercise: Oxidative stress defence, exercise performance, and adaptation. J. Int. Soc. Sports Nutr. 2017, 14, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, D.; Williams, C.; McGregor, S.J.; Nicholas, C.W.; McArdle, F.; Jackson, M.J.; Powell, J.R. Prolonged vitamin C supplementation and recovery from demanding exercise. Int. J. Sport Nutr. 2001, 11, 466–481. [Google Scholar] [CrossRef] [PubMed]
- Mastaloudis, A.; Morrow, J.D.; Hopkins, D.W.; Devaraj, S.; Traber, M.G. Antioxidant supplementation prevents exercise-induced lipid peroxidation, but not inflammation, in ultramarathon runners. Free Radic. Biol. Med. 2004, 36, 1329–1341. [Google Scholar] [CrossRef]
- Gauche, E.; Lepers, R.; Rabita, G.; Leveque, J.M.; Bishop, D.; Brisswalter, J.; Hausswirth, C. Vitamin and mineral supplementation and neuromuscular recovery after a running race. Med. Sci. Sports Exerc. 2006, 38, 2110–2117. [Google Scholar] [CrossRef] [Green Version]
- Vitale, K.C.; Hueglin, S.; Broad, E. Tart Cherry Juice in Athletes. Curr. Sports Med. Rep. 2017, 16, 230–239. [Google Scholar] [CrossRef]
- McAnulty, S.R.; McAnulty, L.S.; Nieman, D.C.; Dumke, C.L.; Morrow, J.D.; Utter, A.C.; Henson, D.A.; Proulx, W.R.; George, G.L. Consumption of blueberry polyphenols reduces exercise-induced oxidative stress compared to vitamin C. Nutr. Res. 2004, 24, 209–221. [Google Scholar] [CrossRef]
- Gaeini, A.A.; Rahnama, N.; Hamedinia, M.R. Effects of vitamin E supplementation on oxidative stress at rest and after exercise to exhaustion in athletic students. J. Sport. Med. Phys. Fit. 2006, 46, 458–461. [Google Scholar]
- Peternelj, T.T.; Coombes, J.S. Antioxidant supplementation during exercise training: Beneficial or detrimental? Sport. Med. 2011, 41, 1043–1069. [Google Scholar] [CrossRef]
- Draeger, C.L.; Naves, A.; Marques, N.; Baptistella, A.B.; Carnauba, R.A.; Paschoal, V.; Nicastro, H. Controversies of antioxidant vitamins supplementation in exercise: Ergogenic or ergolytic effects in humans? J. Int. Soc. Sports Nutr. 2014, 11, 4. [Google Scholar] [CrossRef] [Green Version]
- Michailidis, Y.; Karagounis, L.G.; Terzis, G.; Jamurtas, A.Z.; Spengos, K.; Tsoukas, D.; Chatzinikolaou, A.; Mandalidis, D.; Stefanetti, R.J.; Papassotiriou, I.; et al. Thiol-based antioxidant supplementation alters human skeletal muscle signaling and attenuates its inflammatory response and recovery after intense eccentric exercise. Am. J. Clin. Nutr. 2013, 98, 233–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, A.C.; McKenna, M.J.; Medved, I.; Murphy, K.T.; Brown, M.J.; Della Gatta, P.; Cameron-Smith, D. Infusion with the antioxidant N-acetylcysteine attenuates early adaptive responses to exercise in human skeletal muscle. Acta Physiol. 2012, 204, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Watson, T.A.; Callister, R.; Taylor, R.D.; Sibbritt, D.W.; Macdonald-Wicks, L.K.; Garg, M.L. Antioxidant restriction and oxidative stress in short-duration exhaustive exercise. Med. Sci. Sports Exerc. 2005, 37, 63–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radak, Z.; Taylor, A.W.; Ohno, H.; Goto, S. Adaptation to exercise-induced oxidative stress: From muscle to brain. Exerc. Immunol. Rev. 2001, 7, 90–107. [Google Scholar]
- Margaritis, I.; Rousseau, A.S. Does physical exercise modify antioxidant requirements? Nutr. Res. Rev. 2008, 21, 3–12. [Google Scholar] [CrossRef] [Green Version]
- He, F.; Li, J.; Liu, Z.; Chuang, C.-C.; Yang, W.; Zuo, L. Redox Mechanism of Reactive Oxygen Species in Exercise. Front. Physiol. 2016, 7, 486. [Google Scholar] [CrossRef] [Green Version]
- Radak, Z.; Zhao, Z.; Koltai, E.; Ohno, H.; Atalay, M. Oxygen consumption and usage during physical exercise: The balance between oxidative stress and ROS-dependent adaptive signaling. Antioxid. Redox Signal. 2013, 18, 1208–1246. [Google Scholar] [CrossRef] [Green Version]
- Yavari, A.; Javadi, M.; Mirmiran, P.; Bahadoran, Z. Exercise-Induced Oxidative Stress and Dietary Antioxidants. Asian J. Sport. Med. 2015, 6, 24898. [Google Scholar] [CrossRef] [Green Version]
- Carlsen, M.H.; Halvorsen, B.L.; Holte, K.; Bøhn, S.K.; Dragland, S.; Sampson, L.; Willey, C.; Senoo, H.; Umezono, Y.; Sanada, C.; et al. The total antioxidant content of more than 3100 foods, beverages, spices, herbs and supplements used worldwide. Nutr. J. 2010, 9, 3. [Google Scholar] [CrossRef]
- Schneider, C.D.; Bock, P.M.; Becker, G.F.; Moreira, J.C.F.; Bello-Klein, A.; Oliveira, A.R. Comparison of the effects of two antioxidant diets on oxidative stress markers in triathletes. Biol. Sport 2018, 35, 181–189. [Google Scholar] [CrossRef]
- Lee, E.C.; Fragala, M.S.; Kavouras, S.A.; Queen, R.M.; Pryor, J.L.; Casa, D.J. Biomarkers in sports and exercise: Tracking health, performance, and recovery in athletes. J. Strength Cond. Res. 2017, 31, 2920–2937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katerji, M.; Filippova, M. Duerksen-Hughes Penelope Approaches and Methods to Measure Oxidative Stress in Clinical Samples: Research Applications in the Cancer Field. Oxid. Med. Cell. Longev. 2019, 1–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2016, 15, 71–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vezzoli, A.; Dellanoce, C.; Mrakic-Sposta, S.; Montorsi, M.; Moretti, S.; Tonini, A.; Pratali, L.; Accinni, R. Oxidative Stress Assessment in Response to Ultraendurance Exercise: Thiols Redox Status and ROS Production according to Duration of a Competitive Race. Oxid. Med. Cell. Longev. 2016, 2016, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Mastaloudis, A.; Leonard, S.W.; Traber, M.G. Oxidative stress in athletes during extreme endurance exercise. Free Radic. Biol. Med. 2001, 31, 911–922. [Google Scholar] [CrossRef]
- Nieman, D.C.; Henson, D.A.; Mcanulty, S.R.; Mcanulty, L.; Swick, N.S.; Utter, A.C.; Vinci, D.M.; Opiela, S.J.; Morrow, J.D.; Mcan-Ulty, S.R.; et al. Influence of vitamin C supplementation on oxidative and immune changes after an ultramarathon. J. Appl. Physiol. 1970, 92. [Google Scholar] [CrossRef] [Green Version]
- Mrakic-Sposta, S.; Gussoni, M.; Moretti, S.; Pratali, L.; Giardini, G.; Tacchini, P.; Dellanoce, C.; Tonacci, A.; Mastorci, F.; Borghini, A.; et al. Effects of Mountain Ultra-Marathon Running on ROS Production and Oxidative Damage by Micro-Invasive Analytic Techniques. PLoS ONE 2015, 10, e0141780. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, G.; Spence, V.A.; McLaren, M.; Hill, A.; Underwood, C.; Belch, J.J.F. Oxidative stress levels are raised in chronic fatigue syndrome and are associated with clinical symptoms. Free Radic. Biol. Med. 2005, 39, 584–589. [Google Scholar] [CrossRef]
- Janse, D.E.; Jonge, X.; Thompson, B.; Han, A. Methodological Recommendations for Menstrual Cycle Research in Sports and Exercise. Med. Sci. Sports Exerc. 2019, 51, 2610–2617. [Google Scholar] [CrossRef]
- Smekal, G.; Von Duvillard, S.P.; Frigo, P.; Tegelhofer, T.; Pokan, R.; Hofmann, P.; Tschan, H.; Baron, R.; Wonisch, M.; Renezeder, K.; et al. Menstrual cycle: No effect on exercise cardiorespiratory variables or blood lactate concentration. Med. Sci. Sports Exerc. 2007, 39, 1098–1106. [Google Scholar] [CrossRef]
- Joyce, K.M.; Stewart, S.H. Standardization of Menstrual Cycle Dataor the Analysis of Intensive Longitudinal Data. In Menstrual Cycle; IntechOpen Limited: London, UK, 2019. [Google Scholar]
- Borrás, C.; Gambini, J.; López-Grueso, R.; Pallardó, F.V.; Viña, J. Direct antioxidant and protective effect of estradiol on isolated mitochondria. Biochim. Biophys. Acta 2010, 1802, 205–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oosthuyse, T.; Bosch, A.N. The effect of the menstrual cycle on exercise metabolism: Implications for exercise performance in eumenorrhoeic women. Sport. Med. 2010, 40, 207–227. [Google Scholar] [CrossRef] [PubMed]
- Marventano, S.; Mistretta, A.; Platania, A.; Galvano, F.; Grosso, G. Reliability and relative validity of a food frequency questionnaire for Italian adults living in Sicily, Southern Italy. Int. J. Food Sci. Nutr. 2016, 67, 857–864. [Google Scholar] [CrossRef] [PubMed]
- Rakicioglu, N.; Acar Tek, N.; Ayaz, A.; Pekcan, G. Photographic Atlas of Food and Meal Portion Sizes; Ata Publications: Ankara, Turkey, 2012; ISBN 978-9944-5508-0-2. [Google Scholar]
- Mancini, F.R.; Affret, A.; Dow, C.; Balkau, B.; Bonnet, F.; Boutron-Ruault, M.C.; Fagherazzi, G. Dietary antioxidant capacity and risk of type 2 diabetes in the large prospective E3N-EPIC cohort. Diabetologia 2018, 61, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Institute of Medicine; Food and Nutrition Board. Nutrient Recommendations: Dietary Reference Intakes (DRI). Available online: https://ods.od.nih.gov/Health_Information/Dietary_Reference_Intakes.aspx (accessed on 25 April 2020).
- Hue, O.; Le Gallais, D.; Chollet, D.; Préfaut, C. Ventilatory threshold and maximal oxygen uptake in present triathletes. Can. J. Appl. Physiol. 2000, 25, 102–113. [Google Scholar] [CrossRef] [Green Version]
- Shing, C.M.; Peake, J.M.; Lim, C.L.; Briskey, D.; Walsh, N.P.; Fortes, M.B.; Ahuja, K.D.K.; Vitetta, L. Effects of probiotics supplementation on gastrointestinal permeability, inflammation and exercise performance in the heat. Eur. J. Appl. Physiol. 2014, 114, 93–103. [Google Scholar] [CrossRef]
- Borg, G. Psychophysical bases of perceived exertion. Med. Sci. Sports Exerc. 1982, 14, 377–381. [Google Scholar] [CrossRef]
- Howley, E.; Bassett, D.; Welch, G. Criteria for maximal oxygen uptake: Review and commentary. Med. Sci. Sports Exerc. 1995, 27, 1292–1300. [Google Scholar] [CrossRef]
- Montenegro, C.F.; Kwong, D.A.; Minow, Z.A.; Davis, B.A.; Lozada, C.F.; Casazza, G.A. Betalain-rich supplementation improves exercise performance and recovery in competitive triathletes. Appl. Physiol. Nutr. Metab. 2017, 42, 166–172. [Google Scholar] [CrossRef] [Green Version]
- Erel, O. A new automated colorimetric method for measuring total oxidant status. Clin. Biochem. 2005, 38, 1103–1111. [Google Scholar] [CrossRef]
- Gass, G.C.; Rogers, S.; Mitchell, R. Blood lactate concentration following maximum exercise in trained subjects. Br. J. Sports Med. 1981, 15, 172–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, S.-L.; Chang, H.; Kim, H.-J.; Kim, Y.-A.; Kim, D.-S.; Ho, S.-H.; Kim, S.-H.; Song, W. Effect of HX108-CS supplementation on exercise capacity and lactate accumulation after high-intensity exercise. J. Int. Soc. Sports Nutr. 2013, 10, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Masi, F.; De Souza Vale, R.G.; Dantas, E.H.M.; Barreto, A.C.L.; Novaes, J.D.S.; Reis, V.M. Is blood lactate removal during water immersed cycling faster than during cycling on land? J. Sport. Sci. Med. 2007, 6, 188–192. [Google Scholar]
- Viechtbauer, W.; Smits, L.; Kotz, D.; Budé, L.; Spigt, M.; Serroyen, J.; Crutzen, R. A simple formula for the calculation of sample size in pilot studies. J. Clin. Epidemiol. 2015, 68, 1375–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Lawrence Erlbaum Associates Publishers: Hillsdale, NJ, USA, 1988. [Google Scholar]
- Volpe, S.L. Micronutrient Requirements for Athletes. Clin. Sports Med. 2007, 26, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Koivisto, A.E.; Olsen, T.; Paur, I.; Paulsen, G.; Bastani, N.E.; Garthe, I.; Raastad, T.; Matthews, J.; Blomhoff, R.; Bøhn, S.K. Effects of antioxidant-rich foods on altitude-induced oxidative stress and inflammation in elite endurance athletes: A randomized controlled trial. PLoS ONE 2019, 14, e0217895. [Google Scholar] [CrossRef] [Green Version]
- Skenderi, K.P.; Tsironi, M.; Lazaropoulou, C.; Anastasiou, C.A.; Matalas, A.-L.; Kanavaki, I.; Thalmann, M.; Goussetis, E.; Papassotiriou, I.; Chrousos, G.P. Changes in free radical generation and antioxidant capacity during ultramarathon foot race. Eur. J. Clin. Invest. 2008, 38, 159–165. [Google Scholar] [CrossRef]
- McAnulty, S.R.; McAnulty, L.S.; Nieman, D.C.; Morrow, J.D.; Shooter, L.A.; Holmes, S.; Heward, C.; Henson, D.A. Effect of alpha-tocopherol supplementation on plasma homocysteine and oxidative stress in highly trained athletes before and after exhaustive exercise. J. Nutr. Biochem. 2005, 16, 530–537. [Google Scholar] [CrossRef]
- Briviba, K.; Watzl, B.; Nickel, K.; Kulling, S.; Bös, K.; Haertel, S.; Rechkemmer, G.; Bub, A. A half-marathon and a marathon run induce oxidative DNA damage, reduce antioxidant capacity to protect DNA against damage and modify immune function in hobby runners. Redox Rep. 2005, 10, 325–331. [Google Scholar] [CrossRef]
- Kawamura, T.; Muraoka, I. Exercise-induced oxidative stress and the effects of antioxidant intake from a physiological viewpoint. Antioxidants 2018, 7, 119. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Xun, P.; Wang, R.; Mao, L.; He, K. Can Magnesium Enhance Exercise Performance? Nutrients 2017, 9, 946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Parra, G.; Mora, R.; Hoeger, B. Maximal oxygen consumption in national elite triathletes that train in high altitude. J. Hum. Sport Exerc. 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Daniels, J.; Daniels, N. Running economy of elite male and elite female runners. Med. Sci. Sports Exerc. 1992, 24, 483–489. [Google Scholar] [CrossRef]
- Devlin, J.; Paton, B.; Poole, L.; Sun, W.; Ferguson, C.; Wilson, J.; Kemi, O.J. Blood lactate clearance after maximal exercise depends on active recovery intensity. J. Sports Med. Phys. Fitness 2014, 54, 271–278. [Google Scholar] [PubMed]
- Moxnes, J.F.; Sandbakk, Ø. The kinetics of lactate production and removal during whole-body exercise. Theor. Biol. Med. Model. 2012, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eston, R. Use of ratings of perceived exertion in sports. Int. J. Sports Physiol. Perform. 2012, 7, 175–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doherty, M.; Smith, P.M. Effects of caffeine ingestion on rating of perceived exertion during and after exercise: A meta-analysis. Scand. J. Med. Sci. Sport. 2005, 15, 69–78. [Google Scholar] [CrossRef]
- Handzlik, M.K.; Gleeson, M. Likely Additive Ergogenic Effects of Combined Preexercise Dietary Nitrate and Caffeine Ingestion in Trained Cyclists. ISRN Nutr. 2013, 2013. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Parameter | Males (n = 12) | Females (n = 12) |
|---|---|---|
| Age (y) | 38.5 (31.3–40.0) | 38.0 ((31.6–44.5) |
| Height (cm) | 179.0 (173.5–184.5) | 162.0 (160.0–166.5) * |
| Body mass (kg) | 72.6 (69.6–81.0) | 56.5 (53.5–59.0) * |
| Fat mass percentage (%) | 12.9 (10.0–16.3) | 19.4 (9.2–12.9) * |
| Fat-free mass (kg) | 63.8 (60.0–68.3) | 45.5 (42.8–47.9) * |
| Maximum oxygen consumption (VO2 max), mL·min−1·kg−1 | 60.7 (52.7–65.1) | 51.0 (48.7–52.2) * |
| Baseline training (h·week−1) | 16.3 (15.0–17.6) | 16.4 (15.5–17.0) |
| Years in ultra-endurance sports (y) | 6.0 (2.75–20.0) | 10.5 (4.0–20.0) |
| Dietary intake per day | %RDA ** | %RDA ** |
| Energy (kcal) | 2571.8 (2057.4–3355.5) | 1871.7 (1589.7–2020.0) * |
| Carbohydrate (%) | 36.1 (33.2–41.5) | 34.0 (29.6–35.9) |
| Carbohydrate (g−1·kg−1·d) | 3.4 (2.3–3.8) | 2.7 (2.1–3.2) |
| Protein (%) | 19.9 (16.7–21.8) | 16.3 (14.8–18.2) * |
| Protein (g−1·kg−1·d) | 1.6 (1.3–2.2) | 1.3 (1.2–1.3) |
| Fat (%) | 43.8 (40.9–49.9) | 49.8 (48.9–52.9) * |
| Omega 3 (g) | 2.9 (2.4–4.1) | 2.6 (1.9–3.6) |
| Omega 6 (g) | 25.4 (19.1–32.0) | 20.0 (14.6– 24.5) |
| Vitamin C (mg) | 145.6 (84.9–186.1) 161.7 | 98.9 (68.0–121.3) 136.7 |
| Vitamin E (mg) | 28.7 (20.8–31.7) 143.6 | 18.9 (12.5–25.2) * 94.4 |
| Vitamin A (RAE) | 729.2 (592.1–815.9) 81.0 | 505.0 (403.5–915.0) 72.1 |
| Selenium (μg) | 52.2 (43.4–65.0) 95.0 | 57.5 (45.2–62.1) 104.5 |
| Zinc (mg) | 14.8 (13.5–20.8) 134.5 | 10.6 (10.1–11.7) * 132.0 |
| Plasma Parameters | Pre-Exercise | Post-Exercise | ∆ (Post- vs. Pre-) | Effect Size (d) | Change (%; Range) | p | |
|---|---|---|---|---|---|---|---|
| TOS a (μmol H2O2eq/L) | Men | 3.9 (3.6–4.5) | 5.0 (4.8–7.3) | 1.1 (0.5–2.9) | 0.50 | 30.3 (11.0–84.0) | 0.015 * |
| Women | 3.4 (3.2–4.1) | 5,0 (4.6–5.6) | 1.7 (0.6–2.3) | 0.59 | 48.2 (18.0–65.4) | 0.004 * | |
| TAS b (μmol Trolox equivalent/L) | Men | 1.6 (1.5–1.6) | 1.8 (1.7–1.8) | 0.2 (0.2–0.3) | 0.62 | 13.6 (11.1–16.1) | 0.002 * |
| Women | 1.4 (1.3–1.5) | 1.6 (1.5–1.7) | 0.2 (0.1–0.2) | 0.63 | 12.0 (9.5–15.9) | 0.002 * | |
| OSI c | Men | 0.3 (0.3–0.4) | 0.4 (0.3 0.4) | 0.1 (0.1–0.1) | 0.63 | 27.5 (19.7–40.1) | 0.002 * |
| Women | 0.3 (0.2–0.3) | 0.3 (0.3– 0.4) | 0.1 (0.0–0.1) | 0.60 | 20.7 (14.1–36.2) | 0.003 * | |
| 8-iso PGF2α (pg/mL) | Men | 230.7 (217.9–305.0) | 444.6 (410.6–482.3) | 193.5 (171.0–228.0) | 0.60 | 80.5 (58.7–112.6) | 0.003 * |
| Women | 223.8 (214.2–268.3) | 458.0 (447.2–556.0) | 219.6 (179.0–290.7) | 0.60 | 99.1 (58.6–143.4) | 0.003 * |
| Dietary Antioxidant Intake (FRAP-mmol/day) | |||||
|---|---|---|---|---|---|
| Males (n = 12) | Females (n = 12) | ||||
| r | p | r | p | ||
| TOSa (μmol H2O2eq/L) | Pre | −0.641 | 0.025 * | −0.741 | 0.006 ** |
| Post | 0.147 | 0.648 | −0.077 | 0.812 | |
| ∆(Post- vs. Pre-) | 0.343 | 0.216 | −0.497 | 0.101 | |
| TAS b (μmol Trolox equivalent/L) | Pre | 0.225 | 0.483 | 0.077 | 0.812 |
| Post | 0.514 | 0.087 | 0.417 | 0.178 | |
| ∆(Post- vs. Pre-) | 0.893 | 0.001 ** | 0.769 | 0.002 ** | |
| OSI c (arbitrary unit) | Pre | 0.320 | 0.311 | −0.451 | 0.141 |
| Post | 0.056 | 0.863 | −0.077 | 0.811 | |
| ∆(Post- vs. Pre-) | −0.409 | 0.187 | 0.266 | 0.404 | |
| 8-isoPGF2a (pg/mL) | Pre | −0.417 | 0.201 | −0.373 | 0.259 |
| Post | −0.573 | 0.066 | 0.055 | 0.873 | |
| ∆(Post- vs. Pre-) | −0.702 | 0.016 * | −0.682 | 0.024 * | |
| Time-to-exhaustion (min) | 0.692 | 0.013 * | −0.028 | 0.931 | |
| Lactate (mmol/L) | ∆(Post- 3rd La-–6th La-) | 0.795 | 0.006 ** | 0.642 | 0.024 * |
| Heart rate (HR) (bpm) | ∆(Post- 3rd La-–6th La-) | 0.246 | 0.440 | 0.074 | 0.819 |
| RPE d | Post | −0.525 | 0.079 | −0.022 | 0.945 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Devrim-Lanpir, A.; Bilgic, P.; Kocahan, T.; Deliceoğlu, G.; Rosemann, T.; Knechtle, B. Total Dietary Antioxidant Intake Including Polyphenol Content: Is It Capable to Fight against Increased Oxidants within the Body of Ultra-Endurance Athletes? Nutrients 2020, 12, 1877. https://doi.org/10.3390/nu12061877
Devrim-Lanpir A, Bilgic P, Kocahan T, Deliceoğlu G, Rosemann T, Knechtle B. Total Dietary Antioxidant Intake Including Polyphenol Content: Is It Capable to Fight against Increased Oxidants within the Body of Ultra-Endurance Athletes? Nutrients. 2020; 12(6):1877. https://doi.org/10.3390/nu12061877
Chicago/Turabian StyleDevrim-Lanpir, Aslı, Pelin Bilgic, Tuğba Kocahan, Gökhan Deliceoğlu, Thomas Rosemann, and Beat Knechtle. 2020. "Total Dietary Antioxidant Intake Including Polyphenol Content: Is It Capable to Fight against Increased Oxidants within the Body of Ultra-Endurance Athletes?" Nutrients 12, no. 6: 1877. https://doi.org/10.3390/nu12061877
APA StyleDevrim-Lanpir, A., Bilgic, P., Kocahan, T., Deliceoğlu, G., Rosemann, T., & Knechtle, B. (2020). Total Dietary Antioxidant Intake Including Polyphenol Content: Is It Capable to Fight against Increased Oxidants within the Body of Ultra-Endurance Athletes? Nutrients, 12(6), 1877. https://doi.org/10.3390/nu12061877