Associations between Genotype–Diet Interactions and Weight Loss—A Systematic Review


Abstract
:1. Introduction
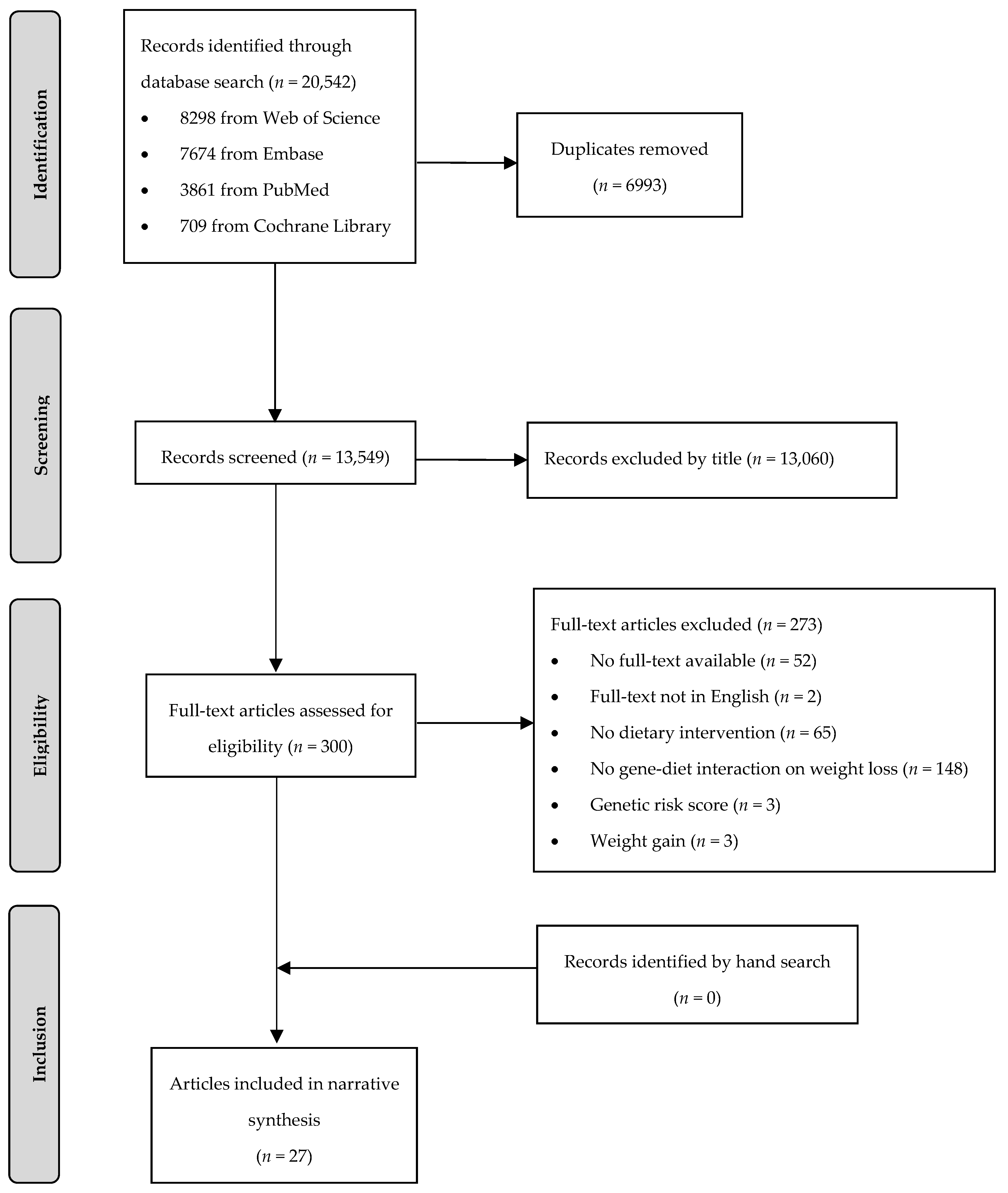
2. Materials and Methods
2.1. Search Strategy
2.2. Study Selection
2.3. Data Extraction
2.4. Reporting Strategy
2.5. Risk of Bias and Quality Assessment
3. Results
3.1. Characteristics of Studies Included
3.2. Study Quality and Risk of Bias
3.3. Main Findings
3.3.1. Interaction of Genotype and Fat Intake on Weight Loss
3.3.2. Interaction of Genotype and Carbohydrate Intake on Weight Loss
3.3.3. Interaction of Genotype and Protein Intake on Weight Loss
4. Discussion
Strengths and Limitations
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bluher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef]
- World Health Organization. Fact Sheet about Obesity and Overweigt. Available online: https://www.who.int/en/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 27 April 2020).
- Sanghera, D.K.; Bejar, C.; Sharma, S.; Gupta, R.; Blackett, P.R. Obesity genetics and cardiometabolic health: Potential for risk prediction. Diabetes Obes. Metab. 2019, 21, 1088–1100. [Google Scholar] [CrossRef]
- Al-Goblan, A.S.; Al-Alfi, M.A.; Khan, M.Z. Mechanism linking diabetes mellitus and obesity. Diabetes Metab Syndr. Obes. 2014, 7, 587–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, J.O.; Wyatt, H.R.; Reed, G.W.; Peters, J.C. Obesity and the environment: Where do we go from here? Science 2003, 299, 853–855. [Google Scholar] [CrossRef] [Green Version]
- Seid, H.; Rosenbaum, M. Low Carbohydrate and Low-Fat Diets: What We Don’t Know and Why we Should Know It. Nutrients 2019, 11, 2749. [Google Scholar] [CrossRef] [Green Version]
- Drabsch, T.; Holzapfel, C. A Scientific Perspective of Personalised Gene-Based Dietary Recommendations for Weight Management. Nutrients 2019, 11, 617. [Google Scholar] [CrossRef] [Green Version]
- Lindgren, C.M.; Heid, I.M.; Randall, J.C.; Lamina, C.; Steinthorsdottir, V.; Qi, L.; Speliotes, E.K.; Thorleifsson, G.; Willer, C.J.; Herrera, B.M.; et al. Genome-wide association scan meta-analysis identifies three Loci influencing adiposity and fat distribution. PLoS Genet. 2009, 5, e1000508. [Google Scholar] [CrossRef]
- Locke, A.E.; Kahali, B.; Berndt, S.I.; Justice, A.E.; Pers, T.H.; Day, F.R.; Powell, C.; Vedantam, S.; Buchkovich, M.L.; Yang, J.; et al. Genetic studies of body mass index yield new insights for obesity biology. Nature 2015, 518, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Speliotes, E.K.; Willer, C.J.; Berndt, S.I.; Monda, K.L.; Thorleifsson, G.; Jackson, A.U.; Lango Allen, H.; Lindgren, C.M.; Luan, J.; Magi, R.; et al. Association analyses of 249,796 individuals reveal 18 new loci associated with body mass index. Nat. Genet. 2010, 42, 937–948. [Google Scholar] [CrossRef] [Green Version]
- Yengo, L.; Sidorenko, J.; Kemper, K.E.; Zheng, Z.; Wood, A.R.; Weedon, M.N.; Frayling, T.M.; Hirschhorn, J.; Yang, J.; Visscher, P.M.; et al. Meta-analysis of genome-wide association studies for height and body mass index in approximately 700000 individuals of European ancestry. Hum. Mol. Genet. 2018, 27, 3641–3649. [Google Scholar] [CrossRef]
- Choquet, H.; Meyre, D. Genetics of Obesity: What have we Learned? Curr. Genom. 2011, 12, 169–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willyard, C. Heritability: The family roots of obesity. Nature 2014, 508, S58–S60. [Google Scholar] [CrossRef] [PubMed]
- Frayling, T.M.; Timpson, N.J.; Weedon, M.N.; Zeggini, E.; Freathy, R.M.; Lindgren, C.M.; Perry, J.R.; Elliott, K.S.; Lango, H.; Rayner, N.W.; et al. A common variant in the FTO gene is associated with body mass index and predisposes to childhood and adult obesity. Science 2007, 316, 889–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, C.D.; Trepanowski, J.F.; Del Gobbo, L.C.; Hauser, M.E.; Rigdon, J.; Ioannidis, J.P.A.; Desai, M.; King, A.C. Effect of Low-Fat vs Low-Carbohydrate Diet on 12-Month Weight Loss in Overweight Adults and the Association with Genotype Pattern or Insulin Secretion: The DIETFITS Randomized Clinical Trial. JAMA 2018, 319, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Celis-Morales, C.; Livingstone, K.M.; Marsaux, C.F.; Macready, A.L.; Fallaize, R.; O’Donovan, C.B.; Woolhead, C.; Forster, H.; Walsh, M.C.; Navas-Carretero, S.; et al. Effect of personalized nutrition on health-related behaviour change: Evidence from the Food4Me European randomized controlled trial. Int. J. Epidemiol. 2017, 46, 578–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grau, K.; Cauchi, S.; Holst, C.; Astrup, A.; Martinez, J.A.; Saris, W.H.; Blaak, E.E.; Oppert, J.M.; Arner, P.; Rossner, S.; et al. TCF7L2 rs7903146-macronutrient interaction in obese individuals’ responses to a 10-wk randomized hypoenergetic diet. Am. J. Clin. Nutr. 2010, 91, 472–479. [Google Scholar] [CrossRef] [Green Version]
- Larsen, T.M.; Dalskov, S.M.; van Baak, M.; Jebb, S.A.; Papadaki, A.; Pfeiffer, A.F.; Martinez, J.A.; Handjieva-Darlenska, T.; Kunesova, M.; Pihlsgard, M.; et al. Diets with high or low protein content and glycemic index for weight-loss maintenance. N. Engl. J. Med. 2010, 363, 2102–2113. [Google Scholar] [CrossRef] [Green Version]
- Xiang, L.; Wu, H.; Pan, A.; Patel, B.; Xiang, G.; Qi, L.; Kaplan, R.C.; Hu, F.; Wylie-Rosett, J.; Qi, Q. FTO genotype and weight loss in diet and lifestyle interventions: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2016, 103, 1162–1170. [Google Scholar] [CrossRef]
- Krug, S.; Kastenmuller, G.; Stuckler, F.; Rist, M.J.; Skurk, T.; Sailer, M.; Raffler, J.; Romisch-Margl, W.; Adamski, J.; Prehn, C.; et al. The dynamic range of the human metabolome revealed by challenges. FASEB J. 2012, 26, 2607–2619. [Google Scholar] [CrossRef]
- Zeevi, D.; Korem, T.; Zmora, N.; Israeli, D.; Rothschild, D.; Weinberger, A.; Ben-Yacov, O.; Lador, D.; Avnit-Sagi, T.; Lotan-Pompan, M.; et al. Personalized Nutrition by Prediction of Glycemic Responses. Cell 2015, 163, 1079–1094. [Google Scholar] [CrossRef] [Green Version]
- Livingstone, K.M.; Celis-Morales, C.; Lara, J.; Ashor, A.W.; Lovegrove, J.A.; Martinez, J.A.; Saris, W.H.; Gibney, M.; Manios, Y.; Traczyk, I.; et al. Associations between FTO genotype and total energy and macronutrient intake in adults: A systematic review and meta-analysis. Obes. Rev. 2015, 16, 666–678. [Google Scholar] [CrossRef] [PubMed]
- Livingstone, K.M.; Celis-Morales, C.; Papandonatos, G.D.; Erar, B.; Florez, J.C.; Jablonski, K.A.; Razquin, C.; Marti, A.; Heianza, Y.; Huang, T.; et al. FTO genotype and weight loss: Systematic review and meta-analysis of 9563 individual participant data from eight randomised controlled trials. BMJ 2016, 354, i4707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booth, A.; Brice, A. Evidence-Based Practice for Information Professionals: A Handbook; Facet Pub.: London, UK, 2004; 304p. [Google Scholar]
- Sterne, J.A.C.; Savovic, J.; Page, M.J.; Elbers, R.G.; Blencowe, N.S.; Boutron, I.; Cates, C.J.; Cheng, H.Y.; Corbett, M.S.; Eldridge, S.M.; et al. RoB 2: A revised tool for assessing risk of bias in randomised trials. BMJ 2019, 366, l4898. [Google Scholar] [CrossRef] [Green Version]
- Sterne, J.A.C.; Hernan, M.A.; Reeves, B.C.; Savovic, J.; Berkman, N.D.; Viswanathan, M.; Henry, D.; Altman, D.G.; Ansari, M.T.; Boutron, I.; et al. ROBINS-I: A tool for assessing risk of bias in non-randomised studies of interventions. BMJ Brit. Med. J. 2016, 355. [Google Scholar] [CrossRef] [Green Version]
- Campbell, H.; Rudan, I. Interpretation of genetic association studies in complex disease. Pharm. J. 2002, 2, 349–360. [Google Scholar] [CrossRef] [Green Version]
- Goni, L.; Qi, L.; Cuervo, M.; Milagro, F.I.; Saris, W.H.; MacDonald, I.A.; Langin, D.; Astrup, A.; Arner, P.; Oppert, J.M.; et al. Effect of the interaction between diet composition and the PPM1K genetic variant on insulin resistance and beta cell function markers during weight loss: Results from the Nutrient Gene Interactions in Human Obesity: Implications for dietary guidelines (NUGENOB) randomized trial. Am. J. Clin. Nutr. 2017, 106, 902–908. [Google Scholar] [CrossRef] [Green Version]
- Grau, K.; Hansen, T.; Holst, C.; Astrup, A.; Saris, W.H.; Arner, P.; Rossner, S.; Macdonald, I.; Polak, J.; Oppert, J.M.; et al. Macronutrient-specific effect of FTO rs9939609 in response to a 10-week randomized hypo-energetic diet among obese Europeans. Int. J. Obes. (Lond.) 2009, 33, 1227–1234. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.L.; De la Cruz, R.; Holst, C.; Grau, K.; Naranjo, C.; Maiz, A.; Astrup, A.; Saris, W.H.; MacDonald, I.; Oppert, J.M.; et al. Allelic variants of melanocortin 3 receptor gene (MC3R) and weight loss in obesity: A randomised trial of hypo-energetic high- versus low-fat diets. PLoS ONE 2011, 6, e19934. [Google Scholar] [CrossRef] [Green Version]
- Sorensen, T.I.; Boutin, P.; Taylor, M.A.; Larsen, L.H.; Verdich, C.; Petersen, L.; Holst, C.; Echwald, S.M.; Dina, C.; Toubro, S.; et al. Genetic polymorphisms and weight loss in obesity: A randomised trial of hypo-energetic high- versus low-fat diets. PLoS Clin. Trials 2006, 1, e12. [Google Scholar] [CrossRef]
- Stocks, T.; Angquist, L.; Banasik, K.; Harder, M.N.; Taylor, M.A.; Hager, J.; Arner, P.; Oppert, J.M.; Martinez, J.A.; Polak, J.; et al. TFAP2B influences the effect of dietary fat on weight loss under energy restriction. PLoS ONE 2012, 7, e43212. [Google Scholar] [CrossRef] [PubMed]
- Svendstrup, M.; Allin, K.H.; Sorensen, T.I.A.; Hansen, T.H.; Grarup, N.; Hansen, T.; Vestergaard, H. Genetic risk scores for body fat distribution attenuate weight loss in women during dietary intervention. Int. J. Obes. (Lond.) 2018, 42, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Goni, L.; Riezu-Boj, J.I.; Milagro, F.I.; Corrales, F.J.; Ortiz, L.; Cuervo, M.; Martinez, J.A. Interaction between an ADCY3 Genetic Variant and Two Weight-Lowering Diets Affecting Body Fatness and Body Composition Outcomes Depending on Macronutrient Distribution: A Randomized Trial. Nutrients 2018, 10, 789. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Lopez, O.; Riezu-Boj, J.I.; Milagro, F.I.; Goni, L.; Cuervo, M.; Martinez, J.A. Differential lipid metabolism outcomes associated with ADRB2 gene polymorphisms in response to two dietary interventions in overweight/obese subjects. Nutr. Metab. Cardiovasc. Dis. 2018, 28, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Goni, L.; Sun, D.; Heianza, Y.; Wang, T.; Huang, T.; Martinez, J.A.; Shang, X.; Bray, G.A.; Smith, S.R.; Sacks, F.M.; et al. A circadian rhythm-related MTNR1B genetic variant modulates the effect of weight-loss diets on changes in adiposity and body composition: The POUNDS Lost trial. Eur. J. Nutr. 2019, 58, 1381–1389. [Google Scholar] [CrossRef]
- Heianza, Y.; Ma, W.; Huang, T.; Wang, T.; Zheng, Y.; Smith, S.R.; Bray, G.A.; Sacks, F.M.; Qi, L. Macronutrient Intake-Associated FGF21 Genotype Modifies Effects of Weight-Loss Diets on 2-Year Changes of Central Adiposity and Body Composition: The POUNDS Lost Trial. Diabetes Care 2016, 39, 1909–1914. [Google Scholar] [CrossRef] [Green Version]
- Heianza, Y.; Sun, D.; Ma, W.; Zheng, Y.; Champagne, C.M.; Bray, G.A.; Sacks, F.M.; Qi, L. Gut-microbiome-related LCT genotype and 2-year changes in body composition and fat distribution: The POUNDS Lost Trial. Int. J. Obes. (Lond.) 2018, 42, 1565–1573. [Google Scholar] [CrossRef]
- Heianza, Y.; Sun, D.; Wang, T.; Huang, T.; Bray, G.A.; Sacks, F.M.; Qi, L. Starch Digestion-Related Amylase Genetic Variant Affects 2-Year Changes in Adiposity in Response to Weight-Loss Diets: The POUNDS Lost Trial. Diabetes 2017, 66, 2416–2423. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.; Wang, T.; Heianza, Y.; Sun, D.; Ivey, K.; Durst, R.; Schwarzfuchs, D.; Stampfer, M.J.; Bray, G.A.; Sacks, F.M.; et al. HNF1A variant, energy-reduced diets and insulin resistance improvement during weight loss: The POUNDS Lost trial and DIRECT. Diabetes Obes. Metab. 2018, 20, 1445–1452. [Google Scholar] [CrossRef]
- Mattei, J.; Qi, Q.; Hu, F.B.; Sacks, F.M.; Qi, L. TCF7L2 genetic variants modulate the effect of dietary fat intake on changes in body composition during a weight-loss intervention. Am. J. Clin. Nutr. 2012, 96, 1129–1136. [Google Scholar] [CrossRef] [Green Version]
- Qi, Q.; Bray, G.A.; Hu, F.B.; Sacks, F.M.; Qi, L. Weight-loss diets modify glucose-dependent insulinotropic polypeptide receptor rs2287019 genotype effects on changes in body weight, fasting glucose, and insulin resistance: The Preventing Overweight Using Novel Dietary Strategies trial. Am. J. Clin. Nutr. 2012, 95, 506–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Q.; Bray, G.A.; Smith, S.R.; Hu, F.B.; Sacks, F.M.; Qi, L. Insulin receptor substrate 1 gene variation modifies insulin resistance response to weight-loss diets in a 2-year randomized trial: The Preventing Overweight Using Novel Dietary Strategies (POUNDS LOST) trial. Circulation 2011, 124, 563–571. [Google Scholar] [CrossRef] [Green Version]
- Qi, Q.; Durst, R.; Schwarzfuchs, D.; Leitersdorf, E.; Shpitzen, S.; Li, Y.; Wu, H.; Champagne, C.M.; Hu, F.B.; Stampfer, M.J.; et al. CETP genotype and changes in lipid levels in response to weight-loss diet intervention in the POUNDS LOST and DIRECT randomized trials. J. Lipid Res. 2015, 56, 713–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Q.; Zheng, Y.; Huang, T.; Rood, J.; Bray, G.A.; Sacks, F.M.; Qi, L. Vitamin D metabolism-related genetic variants, dietary protein intake and improvement of insulin resistance in a 2 year weight-loss trial: POUNDS Lost. Diabetologia 2015, 58, 2791–2799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, D.; Heianza, Y.; Li, X.; Shang, X.; Smith, S.R.; Bray, G.A.; Sacks, F.M.; Qi, L. Genetic, epigenetic and transcriptional variations at NFATC2IP locus with weight loss in response to diet interventions: The POUNDS Lost Trial. Diabetes Obes. Metab. 2018, 20, 2298–2303. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Ng, S.S.; Bray, G.A.; Ryan, D.H.; Sacks, F.M.; Ning, G.; Qi, L. Dietary Fat Intake Modifies the Effect of a Common Variant in the LIPC Gene on Changes in Serum Lipid Concentrations during a Long-Term Weight-Loss Intervention Trial. J. Nutr. 2015, 145, 1289–1294. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Qi, Q.; Liang, J.; Bray, G.A.; Hu, F.B.; Sacks, F.M.; Qi, L. Genetic determinant for amino acid metabolites and changes in body weight and insulin resistance in response to weight-loss diets: The Preventing Overweight Using Novel Dietary Strategies (POUNDS LOST) trial. Circulation 2013, 127, 1283–1289. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Qi, Q.; Bray, G.A.; Hu, F.B.; Sacks, F.M.; Qi, L. APOA5 genotype modulates 2-y changes in lipid profile in response to weight-loss diet intervention: The Pounds Lost Trial. Am. J. Clin. Nutr. 2012, 96, 917–922. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Qi, Q.; Liang, J.; Hu, F.B.; Sacks, F.M.; Qi, L. Neuropeptide Y promoter polymorphism modifies effects of a weight-loss diet on 2-year changes of blood pressure: The preventing overweight using novel dietary strategies trial. Hypertension 2012, 60, 1169–1175. [Google Scholar] [CrossRef] [Green Version]
- Razquin, C.; Martinez, J.A.; Martinez-Gonzalez, M.A.; Fernandez-Crehuet, J.; Santos, J.M.; Marti, A. A Mediterranean diet rich in virgin olive oil may reverse the effects of the -174G/C IL6 gene variant on 3-year body weight change. Mol. Nutr. Food Res. 2010, 54 (Suppl. 1), S75–S82. [Google Scholar] [CrossRef]
- Di Renzo, L.; Cioccoloni, G.; Falco, S.; Abenavoli, L.; Moia, A.; Sinibaldi Salimei, P.; De Lorenzo, A. Influence of FTO rs9939609 and Mediterranean diet on body composition and weight loss: A randomized clinical trial. J. Transl. Med. 2018, 16, 308. [Google Scholar] [CrossRef] [PubMed]
- Garaulet, M.; Smith, C.E.; Hernandez-Gonzalez, T.; Lee, Y.C.; Ordovas, J.M. PPARgamma Pro12Ala interacts with fat intake for obesity and weight loss in a behavioural treatment based on the Mediterranean diet. Mol. Nutr. Food Res. 2011, 55, 1771–1779. [Google Scholar] [CrossRef] [Green Version]
- Petersen, M.; Taylor, M.A.; Saris, W.H.; Verdich, C.; Toubro, S.; Macdonald, I.; Rossner, S.; Stich, V.; Guy-Grand, B.; Langin, D.; et al. Randomized, multi-center trial of two hypo-energetic diets in obese subjects: High- versus low-fat content. Int. J. Obes. (Lond.) 2006, 30, 552–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacks, F.M.; Bray, G.A.; Carey, V.J.; Smith, S.R.; Ryan, D.H.; Anton, S.D.; McManus, K.; Champagne, C.M.; Bishop, L.M.; Laranjo, N.; et al. Comparison of weight-loss diets with different compositions of fat, protein, and carbohydrates. N. Engl. J. Med. 2009, 360, 859–873. [Google Scholar] [CrossRef] [Green Version]
- de Jonge, L.; Bray, G.A.; Smith, S.R.; Ryan, D.H.; de Souza, R.J.; Loria, C.M.; Champagne, C.M.; Williamson, D.A.; Sacks, F.M. Effect of diet composition and weight loss on resting energy expenditure in the POUNDS LOST study. Obesity (Silver Spring) 2012, 20, 2384–2389. [Google Scholar] [CrossRef] [Green Version]
- Shai, I.; Schwarzfuchs, D.; Henkin, Y.; Shahar, D.R.; Witkow, S.; Greenberg, I.; Golan, R.; Fraser, D.; Bolotin, A.; Vardi, H.; et al. Weight loss with a low-carbohydrate, Mediterranean, or low-fat diet. N. Engl. J. Med. 2008, 359, 229–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estruch, R.; Ros, E.; Salas-Salvado, J.; Covas, M.I.; Corella, D.; Aros, F.; Gomez-Gracia, E.; Ruiz-Gutierrez, V.; Fiol, M.; Lapetra, J.; et al. Primary prevention of cardiovascular disease with a Mediterranean diet. N. Engl. J. Med. 2013, 368, 1279–1290. [Google Scholar] [CrossRef] [Green Version]
- Qi, Q.; Kilpelainen, T.O.; Downer, M.K.; Tanaka, T.; Smith, C.E.; Sluijs, I.; Sonestedt, E.; Chu, A.Y.; Renstrom, F.; Lin, X.; et al. FTO genetic variants, dietary intake and body mass index: Insights from 177,330 individuals. Hum. Mol. Genet. 2014, 23, 6961–6972. [Google Scholar] [CrossRef]
- Consortium, S.T.D.; Estrada, K.; Aukrust, I.; Bjorkhaug, L.; Burtt, N.P.; Mercader, J.M.; Garcia-Ortiz, H.; Huerta-Chagoya, A.; Moreno-Macias, H.; Walford, G.; et al. Association of a low-frequency variant in HNF1A with type 2 diabetes in a Latino population. JAMA 2014, 311, 2305–2314. [Google Scholar] [CrossRef] [Green Version]
- Najmi, L.A.; Aukrust, I.; Flannick, J.; Molnes, J.; Burtt, N.; Molven, A.; Groop, L.; Altshuler, D.; Johansson, S.; Bjorkhaug, L.; et al. Functional Investigations of HNF1A Identify Rare Variants as Risk Factors for Type 2 Diabetes in the General Population. Diabetes 2017, 66, 335–346. [Google Scholar] [CrossRef] [Green Version]
- Morita, K.; Saruwatari, J.; Tanaka, T.; Oniki, K.; Kajiwara, A.; Otake, K.; Ogata, Y.; Nakagawa, K. Associations between the common HNF1A gene variant p.I27L (rs1169288) and risk of type 2 diabetes mellitus are influenced by weight. Diabetes Metab. 2015, 41, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Columbus, J.; Chiang, Y.; Shao, W.; Zhang, N.; Wang, D.; Gaisano, H.Y.; Wang, Q.; Irwin, D.M.; Jin, T. Insulin treatment and high-fat diet feeding reduces the expression of three Tcf genes in rodent pancreas. J. Endocrinol. 2010, 207, 77–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harries, L.W.; Brown, J.E.; Gloyn, A.L. Species-specific differences in the expression of the HNF1A, HNF1B and HNF4A genes. PLoS ONE 2009, 4, e7855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Luis, D.A.; Izaola, O.; Primo, D.; Aller, R. Effect of the rs1862513 variant of resistin gene on insulin resistance and resistin levels after two hypocaloric diets with different fat distribution in subjects with obesity. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3865–3872. [Google Scholar] [CrossRef] [PubMed]
- Munafo, M.R.; Clark, T.G.; Flint, J. Assessing publication bias in genetic association studies: Evidence from a recent meta-analysis. Psychiatry Res. 2004, 129, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Orthofer, M.; Valsesia, A.; Magi, R.; Wang, Q.P.; Kaczanowska, J.; Kozieradzki, I.; Leopoldi, A.; Cikes, D.; Zopf, L.M.; Tretiakov, E.O.; et al. Identification of ALK in Thinness. Cell 2020, 181, 1246–1262.e22. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Study Name | Country, Ethnicity | Study Population | Intervention | Duration of Intervention | Weight Loss in kg (Mean ± SD) | Collection of Dietary Data | Reference |
|---|---|---|---|---|---|---|---|
| NUGENOB | United Kingdom, Netherlands, France, Spain, Czech Republic, Sweden, Denmark Caucasian | 771 participants inclusion: BMI ≥ 30 kg/m2; age 20–50 years exclusion: weight change > 3 kg in last 3 months; drug treated hypertension; diabetes mellitus; hyperlipidaemia; untreated thyroid disease; surgically/drug-treated obesity; pregnancy; alcohol/drug abuse; participation in other study. | 600 kcal/day less (1) low-fat diet: 20–25 E% fat, 15 E% protein, 60–65 E% carbs (2) high-fat diet: 40–45 E% fat, 15 E% protein, 40–45 E% carbs | 10 weeks | (1) −6.9 ± 3.4 (2) −6.6 ± 3.5 | Dietary record | [55] |
| Obekit | Spain: Caucasian Hispanics | 147 participants inclusion: BMI 25–40 kg/m2; unrelated exclusion: cardiovascular disease; diabetes mellitus treated with insulin; pregnant and lactating women; use of medications that affect body weight; weight change > 3 kg in last 3 months; unstable use of medication for hyperlipidaemia, type 2 diabetes and treatment of hypertension. | 30% energy restriction (1) low-fat diet: 22 E% fat, 18 E% protein, 60 E% carbs (2) moderately high protein diet: 30 E% fat, 30 E% protein, 40 E% carbs | 16 weeks | (1) −8.1 ± 4.1 (2) −7.6 ± 4.0 | Dietary record | [35] |
| POUNDS Lost | United States: 80% Whites, 15% African Americans, 3% Hispanics, 2% Asians or other | 811 participants inclusion: BMI 25–40 kg/m2; age 30–70 years exclusion: diabetes mellitus; cardiovascular disease; medications that affect body weight; insufficient motivation. | 750 kcal/day less (1) low fat/low-protein diet: 20 E% fat, 15 E% protein, 65 E% carbs (2) low-fat/high-protein diet: 20 E% fat, 25 E% protein, 55 E% carbs (3) high-fat/low-protein diet: 40 E% fat, 15 E% protein, 45 E% carbs (4) high-fat/high-protein diet: 40 E% fat, 25 E% protein, 35 E% carbs | 2 years | 6 months (1) −6.54 ± 0.42 (2) −6.80 ± 0.42 (3) −6.37 ± 0.42 (4) −6.42 ± 0.42 | 24 h recall | [56,57] |
| 2 years (1) −3.26 ± 0.56 (2) −5.03 ± 0.58 (3) −3.87 ± 0.59 (4) −3.98 ± 0.42 | |||||||
| DIRECT | Israel | 322 participants inclusion: BMI ≥ 27 kg/m2; age 40–65 years; presence of type 2 diabetes or coronary heart disease exclusion: pregnant or lactating women; serum creatinine level ≥ 2 mg/dl; liver dysfunction; gastrointestinal problems; active cancer; participating in another diet trial. | (1) low-fat diet: 1500 kcal women, 1800 kcal men, 30 E% fat, 10 E% saturated fats, 300 mg cholesterol intake (2) Mediterranean diet: 1500 kcal women, 1800 kcal men, no more than 35 E% fat, 30 to 45 g of added olive oil and a handful of nuts (3) low-carbohydrate diet: non-restricted calorie diet, 120 g carbohydrates, based on Atkins diet | 2 years | (1) −2.9 ± 4.2 (2) −4.4 ± 6.0 (3) −4.7 ± 6.5 | FFQ and 24 h recall | [58] |
| PREDIMED | Spain: European | 7447 participants inclusion: age 55–80 (men)/60–80 (women) years; diabetes or three or more major cardiovascular risk factors exclusion: history of cardiovascular disease; severe chronic illness; drug or alcohol addiction; history of allergy or intolerance to olive oil or nuts; low predicted likelihood of changing dietary habits. | (1) low-fat diet (2) Mediterranean diet + olive oil (3) Mediterranean diet + nuts | 4 years | (1) −0.10 ± 0.3 (2) −0.21 ± 0.2 (3) −0.07 ± 3.8 | Questionnaire | [52,59] |
| DiOGenes | Netherlands, Denmark, United Kingdom, Greece, Germany, Spain, Bulgaria, Czech Republic | 938 participants inclusion: BMI 27–45 kg/m2; age < 65 years exclusion: > 3 kg weight change within 2 months prior to the study; medication; certain disease. | low-calorie diet: Modifast diet, four items per day, one item between 202–218 kcal, 880 kcal, fat 20 E%, carbs 54 E%, protein 26 E% | 8 weeks | −11.1 ± 3.5 | Dietary record | [18] |
| No acronym | Italy: Caucasian | 300 participants inclusion: Caucasian; Italian; age > 16 years. | (1) control group: general recommendations (2) Mediterranean diet: isocaloric, <25 E% fat, 20 E% protein, 55 E% carbs | 4 weeks | (1) TT genotype: −1.27 ± 3.89 A carriers: −0.62 ± 1.26 (2) TT genotype: −3.41 ± 6.47 A carriers: −2.25 ± 11.79 | n. a. | [53] |
| No acronym | Spain | 1465 participants inclusion: BMI 25–39.99 kg/m2; age 20–65 years exclusion: medication for blood pressure, lowering glucose or lipids; diabetes mellitus; chronic renal failure; hepatic disease; cancer. | 600 kcal less; women: 1200–1800 kcal/day; men: 1500–2000 kcal/day; Mediterranean diet: 35 E% fat (<10 E% saturated fats + 20 E% monounsaturated fats), 15–20 E% protein, 50 E% carbs | different between participants | G carriers: −6.84 ± 5.54 CC genotype: −7.35 ± 5.68 | n. a. | [54] |
| Gene | SNP | Study Name | Sample Size | Study Population of Interaction Term | Time Point of Weight Measurement | Results (p-Value) | Reference |
|---|---|---|---|---|---|---|---|
| ADAMTS9 | rs6795735 | NUGENOB | 559–580 | All participants | 10 weeks | 0.2 1,5,21 | [33] |
| ADCY3 | rs10182181 | Obekit | 101 | 16 weeks | p = 0.02 2,6,21, p = 0.04 2,6,22 Additive model: carriers of the GG (minor allele G) genotype greater weight loss with low-fat diet than carriers of the AG or AA genotypes Co-dominant model: carriers of the GG and AA genotype less weight loss with low-fat diet than carriers of the AG genotype | [35] | |
| ADIPOQ | rs266729 | NUGENOB | 642 | 10 weeks | 0.029 2,7,23 Carriers of the GG and GC (minor allele G) genotype greater weight loss on high-fat diet than carriers of the CC genotype; Carriers of the GC (minor allele G) genotype greater weight loss on low-fat diet than carriers of the CC genotype | [32] | |
| rs2241766 | 0.18 2,7,23 | ||||||
| rs1501299 | 0.14 2,7,23 | ||||||
| rs17300539 | 0.07 2,7,23 | ||||||
| ADRB2 | rs1042713 | Obekit | 107 | 4 months | 0.71 3,8,23 | [36] | |
| rs1042714 | 0.86 3,8,23 | ||||||
| AMY1-AMY2 | rs11185098 | POUNDS Lost | 692 | Whites + Blacks | 2 years | n. s. 2,9,21 | [40] |
| APOA5 | rs964184 | 734 | All participants | n. s. 2,10,21 | [50] | ||
| CART | rs7379701 * | NUGENOB | 642 | 10 weeks | 0.36 2,7,23 | [32] | |
| rs6453132 * | 0.10 2,7,23 | ||||||
| rs17358216 | 0.72 2,7,23 | ||||||
| rs5868607 | 0.68 2,7,23 | ||||||
| CD36 | rs2232169 | 0.42 2,7,23 | |||||
| CETP | rs3764261 | POUNDS Lost + DIRECT | 723 + 171 | Pooled | 2 years | n. s. 2,11,23 | [45] |
| CTNNBL1 | rs9939609 | NUGENOB | 559–580 | All participants | 10 weeks | 0.7 1,5,21 | [33] |
| CYP2R1 | rs10741657 | POUNDS Lost | 732 | 6 months | 0.22 2,12,21 | [46] | |
| 2 years | 0.02 2,12,21 Carriers of the AA (minor allele A) genotype less weight loss with low-fat diet than carriers of the AG or GG genotype | ||||||
| 576 | Whites | 6 months | n. s. 2,11,21 | ||||
| 2 years | < 0.05 2,11,21 Carriers of the AA (minor allele A) genotype less weight loss with low-fat diet than carriers of the AG or GG genotype | ||||||
| DHCR7 | rs12785878 | 732 | All participants | 6 months | 0.80 2,12,21 | ||
| 2 years | 0.22 2,12,21 | ||||||
| 584 | Whites | 6 months | n. s. 2,10,21 | ||||
| 2 years | n. s. 2,10,21 | ||||||
| DNM3-PIGC | rs1011731 | NUGENOB | 559–580 | All participants | 10 weeks | 0.2 1,5,21 | [33] |
| ENPP1 | rs1799774 | 642 | 0.62 2,7,23 | [32] | |||
| rs1044498 | 0.85 2,7,23 | ||||||
| rs7754561 | 0.13 2,7,23 | ||||||
| FANCL | rs887912 | 559–580 | 0.8 1,5,21 | [33] | |||
| FOXC2 | rs34221221 | 642 | 0.75 2,7,23 | [32] | |||
| FTO | rs9939609 | No acronym | 188 | 4 weeks | 0.87 3,13,23 | [53] | |
| NUGENOB | 734 | 10 weeks | 0.55 2,7,24 | [30] | |||
| GAD2 | rs928197 * | 642 | 0.56 2,7,23 | [32] | |||
| rs992990 | 0.78 2,7,23 | ||||||
| rs2236418 * | 0.88 2,7,23 | ||||||
| GC | rs2282679 | POUNDS Lost | 732 | 6 months | 0.17 2,12,21 | [46] | |
| 2 years | 0.08 2,12,21 | ||||||
| 576 | Whites | 6 months | n. s. 2,10,21 | ||||
| 2 years | n. s. 2,10,21 | ||||||
| GHRL | rs696217 | NUGENOB | 642 | All participants | 10 weeks | 0.33 2,7,23 | [32] |
| GHSR | rs2232169 | 0.35 2,7,23 | |||||
| GIPR | rs2287019 | POUNDS Lost | 737 | 6 months | 0.08 2,14,21 | [43] | |
| 2 years | n. s. 2,14,21 | ||||||
| 590 | Whites | 6 months | 0.182,13,21 | ||||
| 2 years | n. s.2,10,21 | ||||||
| GPRC5B | rs12444979 | NUGENOB | 559–580 | All participants | 10 weeks | 0.0461,5,21 | [33] |
| HNF1A | rs7957197 | POUNDS Lost | 722 | 6 months | 0.006 2,12,23 Carriers of the TT and AT (minor allele T) genotype greater weight loss with high-fat diet than carriers of the AA genotype | [41] | |
| 575 | Whites | 0.001 2,15,23 Carriers of the TT and AT (minor allele T) genotype greater weight loss with high-fat diet than carriers of the AA genotype | |||||
| DIRECT | 171 | All participants | 0.03 2,16,23 Carriers of the TT and AT (minor allele T) genotype greater weight loss with high-fat diet than carriers of the AA genotype | ||||
| POUNDS Lost + DIRECT | 722 + 171 | Pooled | 0.001 2,12,23 Carriers of the TT and AT (minor allele T) genotype greater weight loss with high-fat diet than carriers of the AA genotype | ||||
| POUNDS Lost | 722 | All participants | 2 years | n. s. 2,10,23 | |||
| DIRECT | 171 | n. s. 2,10,23 | |||||
| POUNDS Lost + DIRECT | Pooled | n. s. 2,10,23 | |||||
| HSD11B1 | rs846919 | NUGENOB | 642 | All participants | 10 weeks | 0.49 2,7,23 | [32] |
| IGF2 | rs3168310 * | 0.34 2,7,23 | |||||
| rs680 * | 0.59 2,7,23 | ||||||
| rs3842759 | 0.77 2,7,23 | ||||||
| IL6 | rs1800795 | PREDIMED | 737 | 3 years | 0.028 3,17,25 Carriers of the CC (minor allele C) genotype greater weight loss on Mediterranean diet with olive oil than carriers of the CG and GG genotype | [52] | |
| 480 | Non-diabetics | 0.007 3,10,25 Carriers of the CC (minor allele C) genotype greater weight loss on Mediterranean diet with olive oil than carriers of the CG and GG genotype | |||||
| 257 | Diabetics | n. s. 3,10,25 | |||||
| NUGENOB | 642 | All participants | 10 weeks | 0.60 2,7,23 | [32] | ||
| KCNJ11 | rs5219 | 0.10 2,7,23 | |||||
| LEPROTL1 | −2625 C > T | 0.12 2,7,23 | |||||
| LIPC | rs6082 | 0.42 2,7,23 | |||||
| rs1800588 * | 0.67 2,7,23 | ||||||
| rs2070895 * | POUNDS Lost | 734 | 2 years | n. s. 2,15,21 | [48] | ||
| LRRN6C | rs10968576 | NUGENOB | 559–580 | 10 weeks | 0.1 1,5,21 | [33] | |
| LY86 | rs1294421 | 0.6 1,5,21 | |||||
| MAF | rs1424233 | 0.1 1,5,21 | |||||
| MAP2K5 | rs2241423 | 0.3 1,5,21 | |||||
| MC3R | rs6024728 | 760 | 0.89 2,7,21 | [31] | |||
| rs6014646 | 0.48 2,7,21 | ||||||
| rs6024730 | 0.57 2,7,21 | ||||||
| rs6024731 | 0.72 2,7,21 | ||||||
| rs11697509 | 0.20 2,7,21 | ||||||
| rs6127698 | 0.81 2,7,21 | ||||||
| rs3746619 * | 0.81 2,7,21 | ||||||
| rs3827103 * | 0.90 2,7,21 | ||||||
| rs1543873 | 0.42 2,7,21 | ||||||
| rs6099058 | 0.80 2,7,21 | ||||||
| MC4R | rs12970134 | 559–580 | 0.4 1,5,21 | [33] | |||
| MKKS | rs1547 | 642 | 0.47 2,7,23 | [32] | |||
| MTIF3 | rs4771122 | 559–580 | 0.3 1,5,21 | [33] | |||
| MTNR1B | rs10830963 | POUNDS Lost | 575 | Whites | 6 months | < 0.05 2,10,21 Carriers of the GG (minor allele G) genotype greater weight loss with low-fat diet than carriers of the GC or CC genotype | [37] |
| 722 | All participants | 0.01 2,12,21 Carriers of the GG (minor allele G) genotype greater weight loss with low-fat diet than carriers of the GC or CC genotype | |||||
| 2 years | 0.19 2,12,21 | ||||||
| NFATC2IP | rs11150675 | 692 | 0.005 2,18,25 Carriers of the AA (minor allele A) genotype less weight loss with low-fat diet than carriers of the AG and GG genotype | [47] | |||
| NPC1 | rs1805081 | NUGENOB | 559–580 | 10 weeks | 0.2 1,5,21 | [33] | |
| NPY | rs16147 | POUNDS Lost | 723 | 2 years | 0.464 2,15,21 | [51] | |
| 575 | Whites | n. s. 2,10,21 | |||||
| 264 | Hypertensive | 0.688 2,15,21 | |||||
| 459 | Non-hypertensive | 0.547 2,15,21 | |||||
| PCSK1 | rs6235 | NUGENOB | 642 | All participants | 10 weeks | 0.76 2,7,23 | [32] |
| PPARG2 | rs1801282 * | No acronym | 1236 | Different between participants | 0.001 4,19,23 Carriers of the GG and GC (minor allele G) genotype less weight loss (% of baseline weight) on high-fat diet than carriers of the CC genotype | [54] | |
| NUGENOB | 642 | 10 weeks | 0.88 2,7,23 | [32] | |||
| rs3856806 | 0.45 2,7,23 | ||||||
| rs7649970 * | 0.87 2,7,23 | ||||||
| PPARG3 | rs10865710 | 0.76 2,7,23 | |||||
| PPARGC1A | rs8192678 | 0.39 2,7,23 | |||||
| rs2932963 | 0.36 2,7,23 | ||||||
| PPM1K | rs1440581 | 757 | 0.94 2,7,21 | [29] | |||
| POUNDS Lost | 734 | 6 months | 0.002 2,15,21 Carriers of the CC (minor allele C) genotype less weight loss with high-fat diet than carriers of the CT or TT genotype | [49] | |||
| 2 years | 0.008 2,15,21 Carriers of the CC (minor allele C) genotype less weight loss with high-fat diet than carriers of the CT or TT genotype | ||||||
| 587 | Whites | 6 months | 0.02 2,16,21 Carriers of the CC (minor allele C) genotype less weight loss with high-fat diet than carriers of the CT or TT genotype | ||||
| 2 years | 0.012,16,21 Carriers of the CC (minor allele C) genotype less weight loss with high-fat diet than carriers of the CT or TT genotype | ||||||
| RSPO3 | rs9491696 | NUGENOB | 559–580 | All participants | 10 weeks | 0.5 1,5,21 | [33] |
| SERPINE1 | rs1799889 | 642 | 0.29 2,7,23 | [32] | |||
| SLC39A8 | rs13107325 | 559–580 | 0.8 1,5,21 | [33] | |||
| SLC6A14 | rs2011162 | 481 | Women | 0.06 2,7,23 | [32] | ||
| 161 | Men | 0.78 2,7,23 | |||||
| SREBF1 | 17 C > G | 642 | All participants | 0.20 2,7,23 | |||
| TCF7L2 | rs7903146 * | 739 | 0.023 2,7,25 Carriers of the TT (minor allele T) genotype less weight loss on high-fat diet than carriers of the TC and CC genotype | [17] | |||
| POUNDS Lost | 588 | Whites | 6 months | 0.28 2,7,21 | [42] | ||
| 2 years | 0.692 2,7,21 | ||||||
| rs12255372 * | 591 | 6 months | 0.057 2,7,21 | ||||
| 2 years | 0.517 2,7,21 | ||||||
| TFAP2B | rs987237 | DiOGenes | 640 | All participants | 8 weeks | 0.4 1,10,21 | [33] |
| NUGENOB | 559–580 | 10 weeks | 0.03 1,5,21 | ||||
| TNFα | rs1800629 | 642 | 0.04 2,7,23 Carriers of the AA (minor allele A) genotype greater weight loss on a high-fat diet than carriers of the GG genotype; Carriers of the AG genotype greater weight loss on low-fat diet than carriers of the GG genotype | [32] | |||
| UCP2 | rs6593669 | 0.25 2,7,23 | |||||
| UCP3 | rs1900849 | 0.86 2,7,23 | |||||
| VEGFA | rs6905288 | 559–580 | 0.4 1,7,23 | [33] | |||
| rs1358980 | 707 | 0.26 2,13,26 | [34] | ||||
| 174 | Men | 0.58 2,20,26 | |||||
| 533 | Women | 0.06 2,20,26 | |||||
| WAC | rs2807761 | 642 | All participants | 0.17 2,7,23 | [32] | ||
| ZNF608 | rs4836133 | 559–580 | 0.9 1,5,21 | [33] |
| Gene | SNP | Sample Size | Study Population of Interaction Term | Time Point of Weight Measurement | Results (p-Value) 1 | Reference |
|---|---|---|---|---|---|---|
| FGF21 | rs838147 | 715 | All participants | 2 years | 0.07 2,5 | [38] |
| 573 | Whites | n. s. 3,5 | ||||
| IRS1 | rs2943641 | 738 | All participants | 6 months | p = 0.037 2,5; p = 0.058 2,6 Additive model: carriers of the TT (minor allele T) genotype greater weight loss with the highest-carbohydrate diet than carriers of the TC or CC genotype. | [44] |
| 2 years | p = 0.84 2,5; p = 0.59 2,6 | |||||
| 591 | Whites | 6 months | p < 0.05 3,5; p < 0.05 3,6 Additive model: carriers of the TT (minor allele T) genotype greater weight loss with the highest-carbohydrate diet than carriers of the TC or CC genotype. | |||
| 2 years | n. s. 3,5, n. s. 3,6 | |||||
| TCF7L2 | rs7903146 * | 588 | 6 months | 0.811 4,5 | [42] | |
| 2 years | 0.948 4,5 | |||||
| rs12255372 * | 591 | 6 months | 0.21 4,5 | |||
| 2 years | 0.403 4,5 |
| Gene | SNP | Sample Size | Study Population of Interaction Term | Time Point of Weight Measurement | Results (p-Value) 1 | Reference |
|---|---|---|---|---|---|---|
| AMY1-AMY2 | rs11185098 | 692 | All participants | 2 years | n. s. 2 | [40] |
| CYP2R1 | rs10741657 | 732 | 6 months | 0.48 3 | [46] | |
| 2 years | 0.19 3 | |||||
| 576 | Whites | 6 months | n. s. 8 | |||
| 2 years | n. s. 8 | |||||
| DHCR7 | rs12785878 | 732 | All participants | 6 months | 0.25 3 | |
| 2 years | 0.52 3 | |||||
| 584 | Whites | 6 months | n. s. 8 | |||
| 2 years | n. s. 8 | |||||
| GC | rs2282679 | 732 | All participants | 6 months | 0.68 3 | |
| 2 years | 0.41 3 | |||||
| 576 | Whites | 6 months | n. s. 8 | |||
| 2 years | n. s. 8 | |||||
| GIPR | rs2287019 | 737 | All participants | 6 months | > 0.35 4 | [43] |
| 2 years | ||||||
| 590 | Whites | 6 months | > 0.35 9 | |||
| 2 years | ||||||
| LCT | rs4988235 | 583 | n. s. 5 | [39] | ||
| NPY | rs16147 | 723 | All participants | n. s. 6 | [51] | |
| 575 | Whites | n. s. 8 | ||||
| PPM1K | rs1440581 | 734 | All participants | 6 months | > 0.05 6 | [49] |
| 2 years | ||||||
| 587 | Whites | 6 months | > 0.05 10 | |||
| 2 years | ||||||
| TCF7L2 | rs7903146 * | 588 | 6 months | 0.906 7 | [42] | |
| 2 years | 0.515 7 | |||||
| rs12255372 * | 591 | 6 months | 0.746 7 | |||
| 2 years | 0.328 7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bayer, S.; Winkler, V.; Hauner, H.; Holzapfel, C. Associations between Genotype–Diet Interactions and Weight Loss—A Systematic Review. Nutrients 2020, 12, 2891. https://doi.org/10.3390/nu12092891
Bayer S, Winkler V, Hauner H, Holzapfel C. Associations between Genotype–Diet Interactions and Weight Loss—A Systematic Review. Nutrients. 2020; 12(9):2891. https://doi.org/10.3390/nu12092891
Chicago/Turabian StyleBayer, Sandra, Vincent Winkler, Hans Hauner, and Christina Holzapfel. 2020. "Associations between Genotype–Diet Interactions and Weight Loss—A Systematic Review" Nutrients 12, no. 9: 2891. https://doi.org/10.3390/nu12092891
APA StyleBayer, S., Winkler, V., Hauner, H., & Holzapfel, C. (2020). Associations between Genotype–Diet Interactions and Weight Loss—A Systematic Review. Nutrients, 12(9), 2891. https://doi.org/10.3390/nu12092891