An Increase in Fat-Free Mass is Associated with Higher Appetite and Energy Intake in Older Adults: A Randomised Control Trial

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
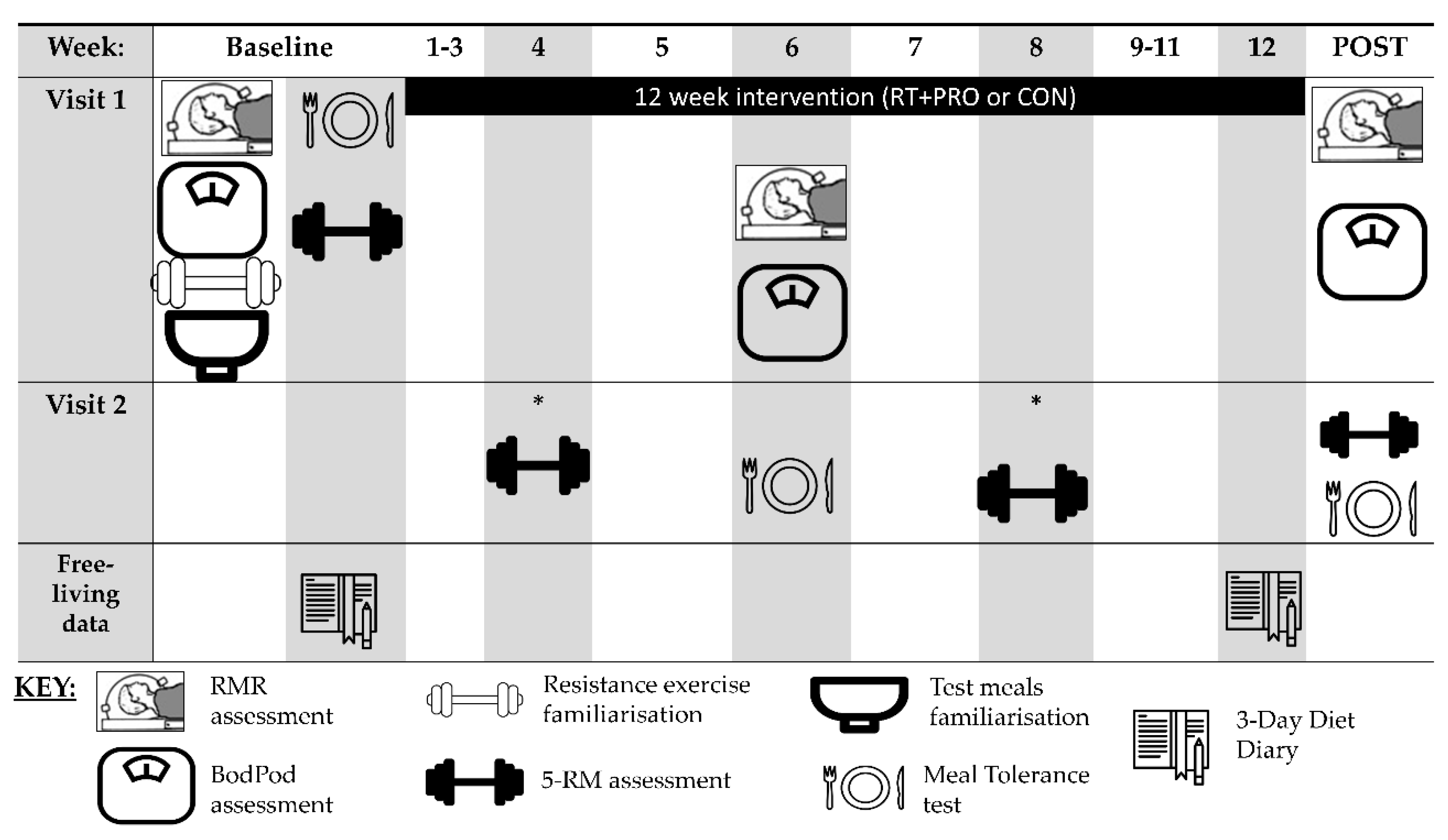
2.2. Experimental Design
2.3. Pre-Testing (Baseline)
2.4. Mid and Post-Testing (Week 6 and Week 12)
2.5. Experimental Protocol
2.6. Outcome Measures
2.6.1. Resting Metabolic Rate
2.6.2. Body Composition
Air Displacement Plethysmography (BOD POD)
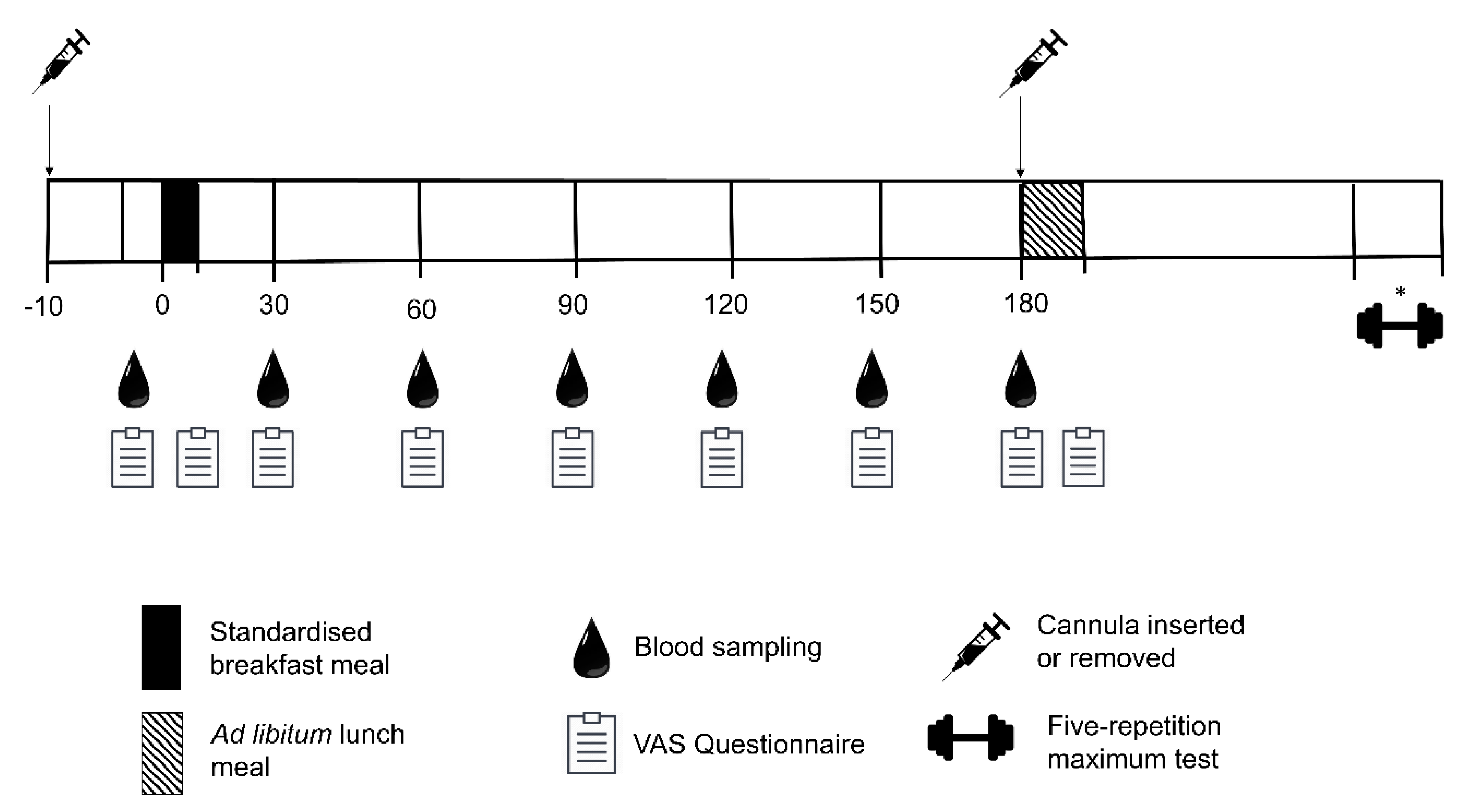
2.6.3. Meal Tolerance Test
Standardised Meals Familiarisation
2.7. Breakfast Test Meal
2.8. Subjective Appetite Sensations
2.9. Blood Sampling
2.10. Assessment of Energy Intake
2.10.1. Ad Libitum Energy Intake
2.10.2. Daily Energy Intake
2.11. Maximal Strength
2.12. Analysis of Blood Samples
2.13. Statistical Analysis
3. Results
3.1. Compliance
3.2. Baseline Paramenters
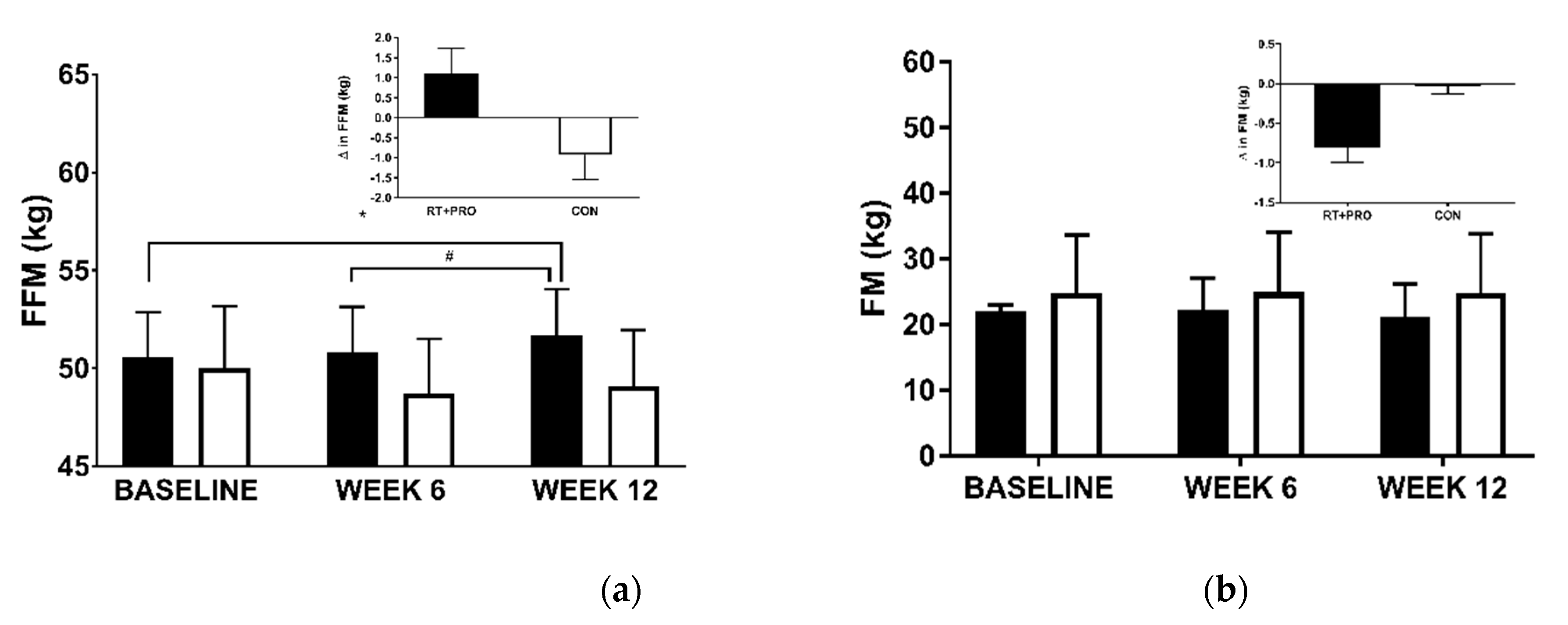
3.2.1. Fat-Free Mass (FFM)
3.2.2. Fat Mass (FM)
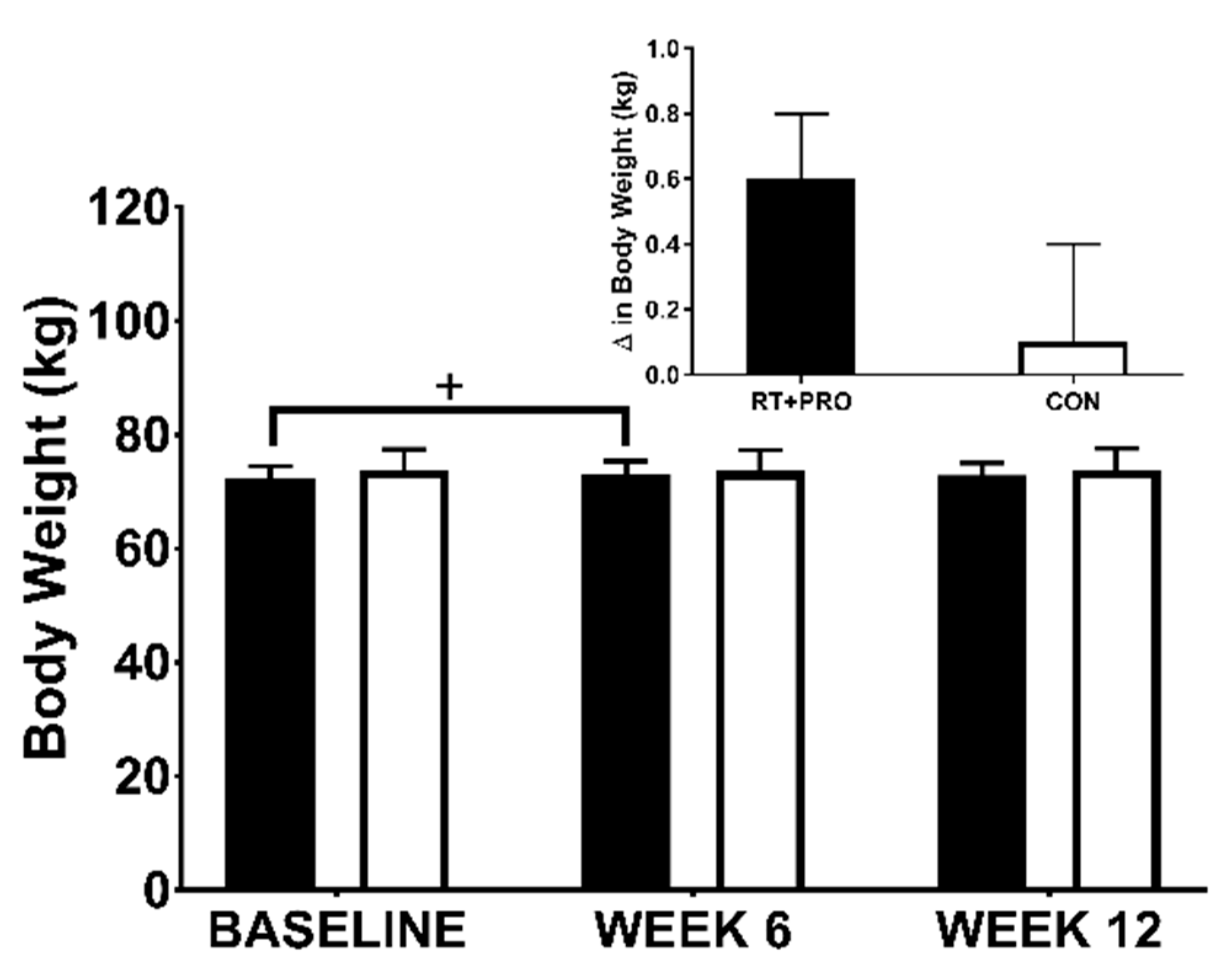
3.2.3. Body Weight
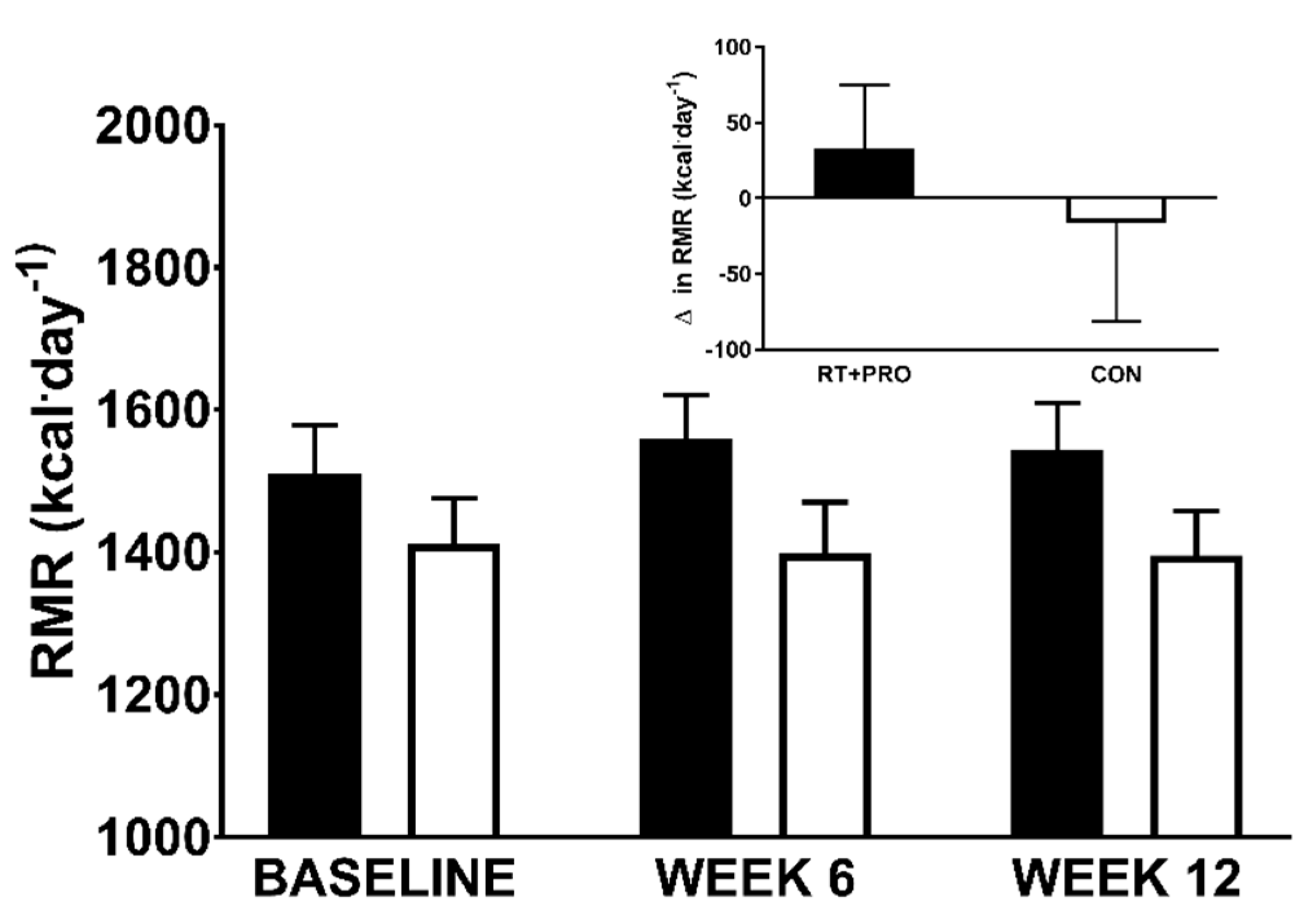
3.2.4. Resting Metabolic Rate
3.3. Subjective Appetite
3.4. Appetite-Relate Hormone Concentrations
3.5. Energy Intake (EI)
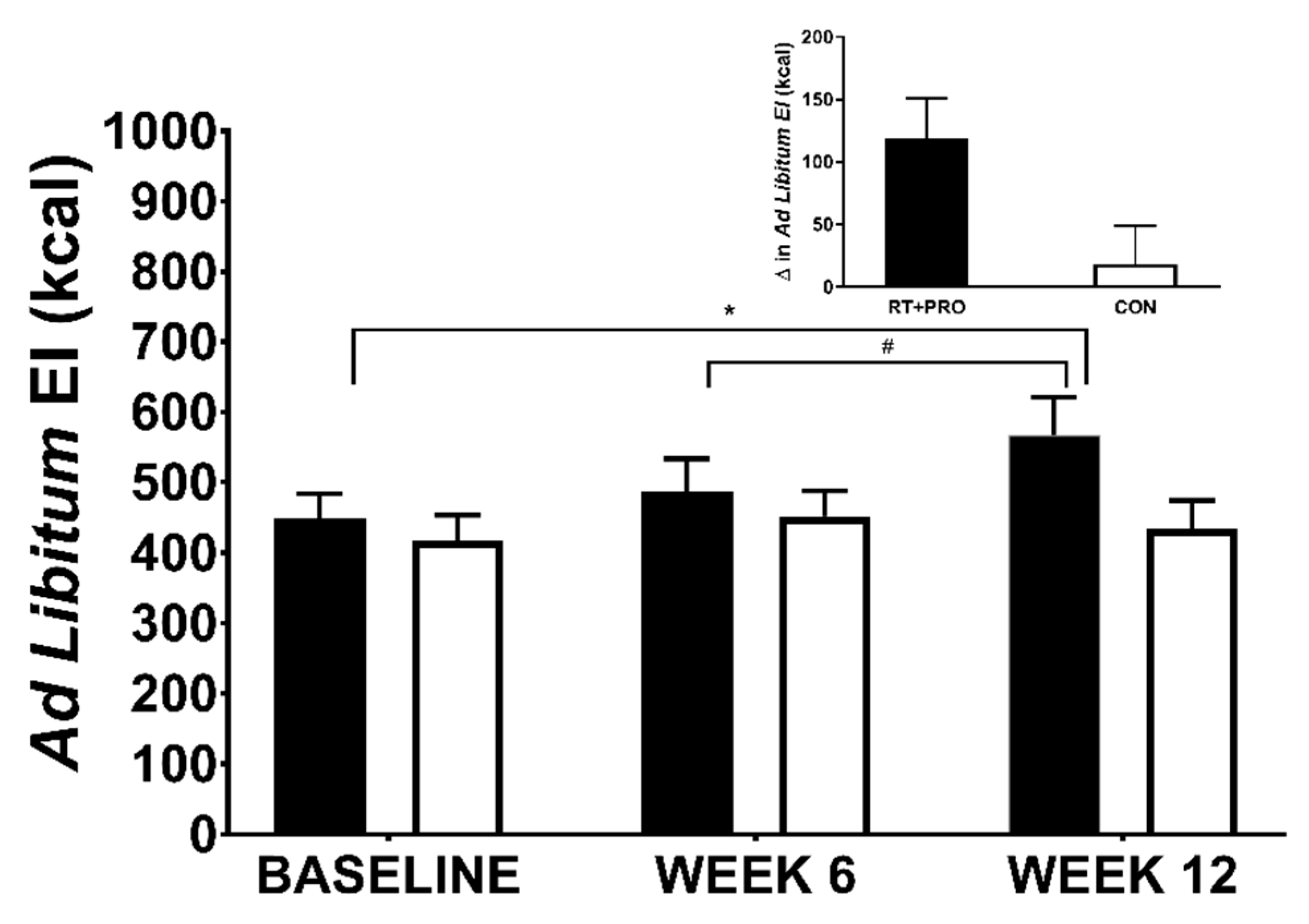
3.5.1. Ad Libitum EI
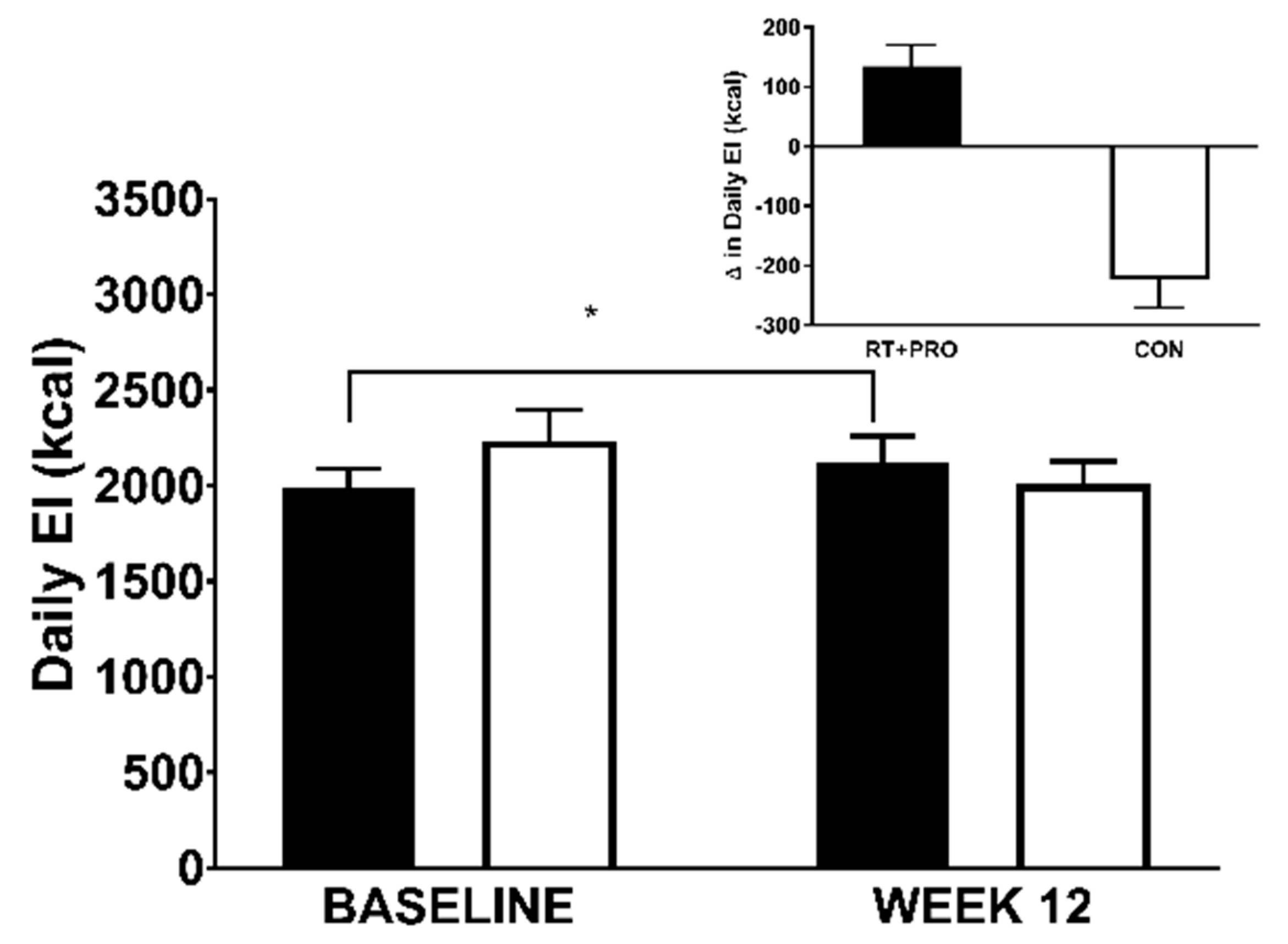
3.5.2. Daily EI
3.5.3. Explaining Group Difference in Ad Libitum EI
3.6. Maximum Strength
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morley, J.E.; Silver, A.J. Anorexia in the elderly. Neurobiol. Aging 1988, 9, 9–16. [Google Scholar] [CrossRef]
- Roberts, S.B.; Rosenberg, I. Nutrition and Aging: Changes in the Regulation of Energy Metabolism with Aging. Physiol. Rev. 2006, 86, 651–667. [Google Scholar] [CrossRef]
- Leij-Halfwerk, S.; Verwijs, M.H.; van Houdt, S.; Borkent, J.W.; Guaitoli, P.; Pelgrim, T.; Heymans, M.W.; Power, L.; Visser, M.; Corish, C.A. Prevalence of protein-energy malnutrition risk in European older adults in community, residential and hospital settings, according to 22 malnutrition screening tools validated for use in adults ≥65 years: A systematic review and meta-analysis. Maturitas 2019, 126, 80–89. [Google Scholar] [CrossRef]
- Rolland, Y.; Van Kan, G.A.; Gillette-Guyonnet, S.; Vellas, B. Cachexia versus sarcopenia. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 15–21. [Google Scholar] [CrossRef]
- Blundell, J.E.; Caudwell, P.; Gibbons, C.; Hopkins, M.; Naslund, E.; King, N.; Finlayson, G. Role of resting metabolic rate and energy expenditure in hunger and appetite control: A new formulation. Dis. Model. Mech. 2012, 5, 608–613. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.O.; Shannon, O.M.; Matu, J.; Holliday, A.; Ispoglou, T.; Deighton, K.; Johnson, K. Differences in circulating appetite-related hormone concentrations between younger and older adults: A systematic review and meta-analysis. Aging Clin. Exp. Res. 2020, 32, 1233–1244. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, A.M.; Murison, S.D.; Duncan, J.S.; Rance, K.A.; Speakman, J.R. Factors influencing variation in basal metabolic rate include fat-free mass, fat mass, age, and circulating thyroxine but not sex, circulating leptin, or triiodothyronine. Am. J. Clin. Nutr. 2005, 82, 941–948. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, A.M.; Rance, K.A.; Murison, S.D.; Duncan, J.S.; Speakman, J.R. Additional anthropometric measures may improve the predictability of basal metabolic rate in adult subjects. Eur. J. Clin. Nutr. 2006, 60, 1437–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravussin, E.; Burnand, B.; Schutz, Y.; Jéquier, E. Twenty-four-hour energy expenditure and resting metabolic rate in obese, moderately obese, and control subjects. Am. J. Clin. Nutr. 1982, 35, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Keys, A.; Taylor, H.L.; Grande, F. Basal metabolism and age of adult man. Metabolism 1973, 22, 579–587. [Google Scholar] [CrossRef]
- Caudwell, P.; Finlayson, G.; Gibbons, C.; Hopkins, M.J.D.; King, N.; Näslund, E.; Blundell, J.E. Resting metabolic rate is associated with hunger, self-determined meal size, and daily energy intake and may represent a marker for appetite. Am. J. Clin. Nutr. 2012, 97, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Westerterp-Plantenga, M.S.; Goris, A.H.C.; Meijer, E.P.; Westerterp, K.R. Habitual meal frequency in relation to resting and activity-induced energy expenditure in human subjects: The role of fat-free mass. Br. J. Nutr. 2003, 90, 643–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weise, C.M.; Hohenadel, M.G.; Krakoff, J.; Votruba, S.B. Body composition and energy expenditure predict ad-libitum food and macronutrient intake in humans. Int. J. Obes. 2013, 38, 243–251. [Google Scholar] [CrossRef] [Green Version]
- Blundell, J.E.; Caudwell, P.; Gibbons, C.; Hopkins, M.; Näslund, E.; King, N.A.; Finlayson, G. Body composition and appetite: Fat-free mass (but not fat mass or BMI) is positively associated with self-determined meal size and daily energy intake in humans. Br. J. Nutr. 2012, 107, 445–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cugini, P.; Salandri, A.; Cilli, M.; Ceccotti, P.; Di Marzo, A.; Rodio, A.; Fontana, S.; Pellegrino, A.M.; De Francesco, G.P.; Coda, S.; et al. Daily hunger sensation and body compartments: II. Their relationships in obese patients. Eat. Weight. Disord. Stud. Anorexia Bulim. Obes. 1999, 4, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Lissner, L.; Habicht, J.P.; Strupp, B.J.; Levitsky, D.A.; Haas, J.D.; Roe, D.A. Body composition and energy intake: Do overweight women overeat and underreport? Am. J. Clin. Nutr. 1989, 49, 320–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkins, M.; Finlayson, G.; Duarte, C.; Whybrow, S.; Ritz, P.; Horgan, G.W.; Blundell, J.E.; Stubbs, R.J. Modelling the associations between fat-free mass, resting metabolic rate and energy intake in the context of total energy balance. Int. J. Obes. 2015, 40, 312–318. [Google Scholar] [CrossRef] [Green Version]
- Stubbs, R.J.; Hopkins, M.; Finlayson, G.S.; Duarte, C.; Gibbons, C.; Blundell, J.E. Potential effects of fat mass and fat-free mass on energy intake in different states of energy balance. Eur. J. Clin. Nutr. 2018, 72, 698–709. [Google Scholar] [CrossRef]
- Blundell, J.E.; Gibbons, C.; Beaulieu, K.; Casanova, N.; Duarte, C.; Finlayson, G.; Stubbs, R.J.; Hopkins, M. The drive to eat in homo sapiens: Energy expenditure drives energy intake. Physiol. Behav. 2020, 219, 112846. [Google Scholar] [CrossRef]
- Blundell, J.E.; Gibbons, C.; Caudwell, P.; Finlayson, G.; Hopkins, M. Appetite control and energy balance: Impact of exercise. Obes. Rev. 2015, 16, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Dorling, J.L.; Broom, D.R.; Burns, S.F.; Clayton, D.J.; Deighton, K.; James, L.; King, J.A.; Miyashita, M.; Thackray, A.E.; Batterham, R.L.; et al. Acute and Chronic Effects of Exercise on Appetite, Energy Intake, and Appetite-Related Hormones: The Modulating Effect of Adiposity, Sex, and Habitual Physical Activity. Nutrients 2018, 10, 1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, N.A.; Burley, V.J.; Blundell, J.E. Exercise-induced suppression of appetite: Effects on food intake and implications for energy balance. Eur. J. Clin. Nutr. 1994, 48, 715–724. [Google Scholar] [PubMed]
- Deighton, K.; Stensel, D.J. Creating an acute energy deficit without stimulating compensatory increases in appetite: Is there an optimal exercise protocol? Proc. Nutr. Soc. 2014, 73, 352–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stubbs, R.J.; Hughes, D.A.; Johnstone, A.M.; Horgan, G.W.; King, N.; Blundell, J.E. A decrease in physical activity affects appetite, energy, and nutrient balance in lean men feeding ad libitum. Am. J. Clin. Nutr. 2004, 79, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Stubbs, R.J.; Hughes, D.A.; Johnstone, A.M.; Whybrow, S.; Horgan, G.W.; King, N.; Blundell, J. Rate and extent of compensatory changes in energy intake and expenditure in response to altered exercise and diet composition in humans. Am. J. Physiol. Integr. Comp. Physiol. 2004, 286, R350–R358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, C.; Kulseng, B.; King, N.A.; Holst, J.J.; Blundell, J.E. The Effects of Exercise-Induced Weight Loss on Appetite-Related Peptides and Motivation to Eat. J. Clin. Endocrinol. Metab. 2010, 95, 1609–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.E.; Phillips, S.M. Maximizing muscle protein anabolism: The role of protein quality. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 66–71. [Google Scholar] [CrossRef]
- Tieland, M.; Dirks, M.L.; Van Der Zwaluw, N.; Verdijk, L.B.; Van De Rest, O.; De Groot, L.C.; Van Loon, L.J. Protein Supplementation Increases Muscle Mass Gain During Prolonged Resistance-Type Exercise Training in Frail Elderly People: A Randomized, Double-Blind, Placebo-Controlled Trial. J. Am. Med Dir. Assoc. 2012, 13, 713–719. [Google Scholar] [CrossRef]
- Jeor, S.T.S.; Brunner, R.L.; Harrington, M.E.; Scott, B.J.; Daugherty, S.A.; Cutter, G.R.; Brownell, K.D.; Dyer, A.R.; Foreyt, J.P. A Classification System to Evaluate Weight Maintainers, Gainers, and Losers. J. Am. Diet. Assoc. 1997, 97, 481–488. [Google Scholar] [CrossRef]
- Goran, M.I. Energy Metabolism and Obesity. Med Clin. North Am. 2000, 84, 347–362. [Google Scholar] [CrossRef]
- Ravussin, E.; Lillioja, S.; Anderson, T.E.; Christin, L.; Bogardus, C. Determinants of 24-hour energy expenditure in man. Methods and results using a respiratory chamber. J. Clin. Investig. 1986, 78, 1568–1578. [Google Scholar] [CrossRef] [Green Version]
- Costanza, M.C. Matching. Prev. Med. 1995, 24, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Rothman, K.J.; Greenland, S.; Lash, T.L. Modern Epidemiology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2008. [Google Scholar]
- Willoughby, D.S. Resistance training in the older adult. In ACSM Current Comment; American College of Sports Medicine: Indiana, IN, USA, 2015. [Google Scholar]
- Mayhew, J.L.; Ball, T.E.; Arnold, M.D.; Bowen, J.C. Relative muscular endurance performance as a predictor of bench press strength in college men and women. J. Strength Cond. Res. 1992, 6, 200–206. [Google Scholar]
- Bauer, J.; Biolo, G.; Cederholm, T.; Cesari, M.; Cruz-Jentoft, A.J.; Morley, J.E.; Phillips, S.; Sieber, C.; Stehle, P.; Teta, D.; et al. Evidence-Based Recommendations for Optimal Dietary Protein Intake in Older People: A Position Paper From the PROT-AGE Study Group. J. Am. Med Dir. Assoc. 2013, 14, 542–559. [Google Scholar] [CrossRef] [PubMed]
- Symons, T.B.; Sheffield-Moore, M.; Wolfe, R.R.; Paddon-Jones, D. Moderating the portion size of a protein-rich meal improves anabolic efficiency in young and elderly. J. Am. Diet. Assoc. 2009, 109, 1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Compher, C.; Frankenfield, D.; Keim, N.; Roth-Yousey, L.; Evidence Analysis Working Group. Best Practice Methods to Apply to Measurement of Resting Metabolic Rate in Adults: A Systematic Review. J. Am. Diet. Assoc. 2006, 106, 881–903. [Google Scholar] [CrossRef]
- Branson, R.; Johannigman, J.A. The Measurement of Energy Expenditure. Nutr. Clin. Pr. 2004, 19, 622–636. [Google Scholar] [CrossRef]
- Henry, C.J.K. Basal metabolic rate studies in humans: Measurement and development of new equations. Public Heal. Nutr. 2005, 8, 1133–1152. [Google Scholar] [CrossRef]
- Weir, J.B.D.V. New methods for calculating metabolic rate with special reference to protein metabolism. J. Physiol. 1949, 109, 1–9. [Google Scholar] [CrossRef]
- Higgins, P.B.; Fields, D.A.; Hunter, G.R.; Gower, B.A. Effect of Scalp and Facial Hair on Air Displacement Plethysmography Estimates of Percentage of Body Fat. Obesity 2001, 9, 326–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCrory, M.A.; Gomez, T.D.; Bernauer, E.; Mole, P. Evaluation of a new air displacement plethysmograph for measuring human body composition. Med. Sci. Sports Exerc. 1995, 27, 1686–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crapo, R.O.; Morris, A.H.; Clayton, P.D.; Nixon, C.R. Lung volumes in healthy nonsmoking adults. Bull. Eur. Physiopathol. Respir. 1982, 18, 419–425. [Google Scholar] [PubMed]
- Dempster, P.; Aitkens, S. A new air displacement method for the determination of human body composition. Med. Sci. Sports Exerc. 1995, 27, 1692–1697. [Google Scholar] [CrossRef]
- Siri, W.E. Body composition from fluid spaces and density: Analysis of methods. 1961. Nutrition 1993, 9, 480–491. [Google Scholar]
- Gibney, M.J.; Lanham-New, S.A.; Cassidy, A.; Vorster, H.H. Introduction to Human Nutrition, 2nd ed.; John Wiley & Sons Ltd. Publication: Hoboken, NJ, USA, 2009. [Google Scholar]
- Gaal, S.; Kerr, M.A.; Ward, M.; McNulty, H.; Livingstone, M.B.E. Breakfast Consumption in the UK: Patterns, Nutrient Intake and Diet Quality. A Study from the International Breakfast Research Initiative Group. Nutrients 2018, 10, 999. [Google Scholar] [CrossRef] [Green Version]
- Blundell, J.; De Graaf, C.; Hulshof, T.; Jebb, S.; Livingstone, B.; Lluch, A.; Mela, D.; Salah, S.; Schuring, E.; Van Der Knaap, H.; et al. Appetite control: Methodological aspects of the evaluation of foods. Obes. Rev. 2010, 11, 251–270. [Google Scholar] [CrossRef] [Green Version]
- Stubbs, R.J.; Hughes, D.A.; Johnstone, A.M.; Rowley, E.; Reid, C.; Elia, M.; Stratton, R.; Delargy, H.; King, N.; Blundell, J.E. The use of visual analogue scales to assess motivation to eat in human subjects: A review of their reliability and validity with an evaluation of new hand-held computerized systems for temporal tracking of appetite ratings. Br. J. Nutr. 2000, 84, 405–415. [Google Scholar] [CrossRef] [Green Version]
- Hill, A.J.; Blundell, J.E. Nutrients and behaviour: Research strategies for the investigation of taste characteristics, food preferences, hunger sensations and eating patterns in man. J. Psychiatr. Res. 1982, 17, 203–212. [Google Scholar] [CrossRef]
- Date, Y. The vagus nerve and ghrelin function. In Metabotropic Glutamate Receptors; Springer Science and Business Media LLC: Berlin, Germany, 2014; Volume 25, pp. 53–61. [Google Scholar]
- Fawcett, J.K.; Wynn, V. Effects of Posture on Plasma Volume and Some Blood Constituents. J. Clin. Pathol. 1960, 13, 304–310. [Google Scholar] [CrossRef] [Green Version]
- Public Health England. Government Dietary Recommendations- Government Recommendations for Energy and Nutrients for Males and Females Aged 1–18 Years and 19+ Years; Public Health England: London, UK, 2016. [Google Scholar]
- Deighton, K.; Frampton, J.; Gonzalez, J.T. Test-meal palatability is associated with overconsumption but better represents preceding changes in appetite in non-obese males. Br. J. Nutr. 2016, 116, 935–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, J.D.; Fletcher, J.P. Reliability of the 8-repetition maximum test in men and wom en. J. Sci. Med. Sport 2012, 15, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Gail, S.; Rodefeld, S.; Künzell, S. Reproducibility of a 5-repetition maximum strength test in older adults. Isokinet. Exerc. Sci. 2015, 23, 291–295. [Google Scholar] [CrossRef]
- Haff, G.; Triplett, N. Essentials of Strength Training and Conditioning, 4th ed.; Human Kinetics: Champaign, IL, USA, 2016. [Google Scholar]
- Schwanbeck, S.; Chilibeck, P.D.; Binsted, G. A Comparison of Free Weight Squat to Smith Machine Squat Using Electromyography. J. Strength Cond. Res. 2009, 23, 2588–2591. [Google Scholar] [CrossRef]
- Wood, T.M.; Maddalozzo, G.F.; Harter, R.A. Accuracy of Seven Equations for Predicting 1-RM Performance of Apparently Healthy, Sedentary Older Adults. Meas. Phys. Educ. Exerc. Sci. 2002, 6, 67–94. [Google Scholar] [CrossRef]
- Torres, V.; Medina, Y.; Rodríguez, Y.; Sardiñas, Y.; Herrera, M.; Rodríguez, R. Application of the linear mixed and generalized mixed model as alternatives for analysis in experiments with repeated measures. Cuban. J. Agric. Sci. 2019, 53, 7–12. [Google Scholar]
- Cnaan, A.; Laird, N.M.; Slasor, P. Using the general linear mixed model to analyse unbalanced repeated measures and longitudinal data. Stat. Med. 1997, 16, 2349–2380. [Google Scholar] [CrossRef]
- Baayen, R.H.; Davidson, D.; Bates, D. Mixed-effects modeling with crossed random effects for subjects and items. J. Mem. Lang. 2008, 59, 390–412. [Google Scholar] [CrossRef] [Green Version]
- Harrison, X.A.; Donaldson, L.; Correa-Cano, M.E.; Evans, J.C.; Fisher, D.N.; Goodwin, C.E.; Robinson, B.S.; Hodgson, D.; Inger, R. A brief introduction to mixed effects modelling and multi-model inference in ecology. PeerJ 2018, 6, e4794. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, W. How to interpret changes in an athletic performance test (Online). Sport science 2004, 8, 1–7. [Google Scholar]
- Hill, R.J.; Davies, P.S.W. The validity of self-reported energy intake as determined using the doubly labelled water technique. Br. J. Nutr. 2001, 85, 415–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, J.; Cohen, P.; West, S.G.; Aiken, L.S. Applied Multiple Regression/Correlation Analysis for the Behavioral Sciences; Routledge: Abingdon, UK, 2013. [Google Scholar]
- Tabachnick, B.G.; Fidell, L.S.; Ullman, J.B. Using Multivariate Statistics; Pearson: Boston, MA, USA, 2007; Volume 5. [Google Scholar]
- Alajmi, N.; Deighton, K.; King, J.A.; Reischak-Oliveira, A.; Wasse, L.K.; Jones, J.; Batterham, R.L.; Stensel, D.J. Appetite and Energy Intake Responses to Acute Energy Deficits in Females versus Males. Med. Sci. Sports Exerc. 2016, 48, 412–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenkilde, M.; Reichkendler, M.H.; Auerbach, P.; Toräng, S.; Gram, A.S.; Ploug, T.; Holst, J.J.; Sjödin, A.; Stallknecht, B. Appetite regulation in overweight, sedentary men after different amounts of endurance exercise: A randomized controlled trial. J. Appl. Physiol. 2013, 115, 1599–1609. [Google Scholar] [CrossRef] [PubMed]
- Bryant, E.J.; Caudwell, P.; Hopkins, M.E.; King, N.A.; Blundell, J.E. Psycho-markers of weight loss. The roles of TFEQ Disinhibition and Restraint in exercise-induced weight management. Appetite 2012, 58, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.; Kulseng, B.; Rehfeld, J.F.; King, N.A.; Blundell, J.E. Effect of Chronic Exercise on Appetite Control in Overweight and Obese Individuals. Med. Sci. Sports Exerc. 2013, 45, 805–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornier, M.-A.; Melanson, E.L.; Salzberg, A.K.; Bechtell, J.L.; Tregellas, J.R. The effects of exercise on the neuronal response to food cues. Physiol. Behav. 2012, 105, 1028–1034. [Google Scholar] [CrossRef] [Green Version]
- Martins, C.; Truby, H.; Morgan, L.M. Short-term appetite control in response to a 6-week exercise programme in sedentary volunteers. Br. J. Nutr. 2007, 98, 834–842. [Google Scholar] [CrossRef] [Green Version]
- Weise, C.M.; Thiyyagura, P.; Reiman, E.M.; Chen, K.; Krakoff, J. A potential role for the midbrain in integrating fat-free mass determined energy needs: An H215O PET study. Hum. Brain Mapp. 2015, 36, 2406–2415. [Google Scholar] [CrossRef] [Green Version]
- Poppitt, S.; McCormack, D.; Buffenstein, R. Short-term effects of macronutrient preloads on appetite and energy intake in lean women. Physiol. Behav. 1998, 64, 279–285. [Google Scholar] [CrossRef]
- Campbell, W.W.; Crim, M.C.; Young, V.R.; Evans, W.J. Increased energy requirements and changes in body composition with resistance training in older adults. Am. J. Clin. Nutr. 1994, 60, 167–175. [Google Scholar] [CrossRef]
- Dulloo, A.G.; Jacquet, J.; Miles-Chan, J.L.; Schutz, Y. Passive and active roles of fat-free mass in the control of energy intake and body composition regulation. Eur. J. Clin. Nutr. 2016, 71, 353–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellinkoff, S.M.; Frankland, M.; Boyle, D.; Greipel, M. Relationship Between Serum Amino Acid Concentration and Fluctuations in Appetite. J. Appl. Physiol. 1956, 8, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Bray, G.A. Amino acids, protein, and body weight. Obes. Res. 1997, 5, 373–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millward, D.J. A Protein-Stat Mechanism for Regulation of Growth and Maintenance of the Lean Body Mass. Nutr. Res. Rev. 1995, 8, 93–120. [Google Scholar] [CrossRef] [Green Version]
- King, N.A.; Caudwell, P.P.; Hopkins, M.; Stubbs, J.R.; Naslund, E.; Blundell, J.E. Dual-process action of exercise on appetite control: Increase in orexigenic drive but improvement in meal-induced satiety. Am. J. Clin. Nutr. 2009, 90, 921–927. [Google Scholar] [CrossRef] [Green Version]
- Caudwell, P.; Gibbons, C.; Hopkins, M.; King, N.; Finlayson, G.; Blundell, J. No Sex Difference in Body Fat in Response to Supervised and Measured Exercise. Med. Sci. Sports Exerc. 2013, 45, 351–358. [Google Scholar] [CrossRef] [Green Version]
- Guelfi, K.J.; Donges, C.E.; Duffield, R. Beneficial effects of 12 weeks of aerobic compared with resistance exercise training on perceived appetite in previously sedentary overweight and obese men. Metabolism 2013, 62, 235–243. [Google Scholar] [CrossRef]
- Beaulieu, K.; Hopkins, M.; Blundell, J.; Finlayson, G. Homeostatic and non-homeostatic appetite control along the spectrum of physical activity levels: An updated perspective. Physiol. Behav. 2018, 192, 23–29. [Google Scholar] [CrossRef]
- Lambert, C.P.; Frank, L.L.; Evans, W.J. Macronutrient Considerations for the Sport of Bodybuilding. Sports Med. 2004, 34, 317–327. [Google Scholar] [CrossRef]
- Morishima, T.; Kurihara, T.; Hamaoka, T.; Goto, K. Whole body, regional fat accumulation, and appetite-related hormonal response after hypoxic training. Clin. Physiol. Funct. Imaging 2014, 34, 90–97. [Google Scholar] [CrossRef]
- O’Driscoll, R.; Turicchi, J.; Beaulieu, K.; Scott, S.; Matu, J.; Deighton, K.; Finlayson, G.; Stubbs, J. How well do activity monitors estimate energy expenditure? A systematic review and meta-analysis of the validity of current technologies. Br. J. Sports Med. 2018, 54, 332–340. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RT + PRO (n = 19) | CON (n = 20) | |
|---|---|---|
| Age (years) | 67 ± 4 | 65 ± 4 |
| Height (m) | 1.67 ± 5.3 | 1.68 ± 8.4 |
| Body mass (kg) | 72.4 ±9.6 | 73.8 ± 16.5 |
| BMI (kg·m−2) | 25.7 ± 3.6 | 26.2 ± 6.1 |
| RT + PRO | CON | p-Value | |
|---|---|---|---|
| Fat-free mass (kg) | 50.5 ± 2.2 | 49.2 ± 2.8 | 0.773 |
| Body Weight (kg) | 72.4 ± 9.3 | 73.8 ± 16.1 | 0.905 |
| Fat mass (kg) | 24.7 ± 9.1 | 21.9 ± 5.3 | 0.336 |
| Resting metabolic rate (kcal/day) | 1543 ± 67 | 1395 ± 65 | 0.297 |
| Ad libitum energy intake (kcal) | 448 ± 42 | 417 ± 41 | 0.543 |
| Daily energy intake (kcal/day) | 1987 ± 122 | 2234 ± 128 | 0.211 |
| Fasted subjective appetite (mm) Subjective appetite AUC (mm·min−1) | 67 ± 16 44 ± 15 | 66 ± 15 39 ± 14 | 0.620 0.205 |
| Fasted leptin (pg·mL) Leptin AUC (pg·mL·min−1) | 20.2 ± 22.4 16.8 ± 17.7 | 25.4 ± 30.2 21.4 ± 27.9 | 0.549 0.551 |
| Fasted insulin (μIU·mL−1) Insulin AUC (μIU·mL·min−1) | 43.0 ± 11.6 13.2 ± 5.2 | 50.6 ± 17.2 19.4 ± 45.0 | 0.039 * 0.117 |
| Baseline | Week 6 | Week 12 | Δ from Baseline to Week 12 | |
|---|---|---|---|---|
| Subjective appetite (mm) | ||||
| RT + PRO | 71 ± 16 | 65 ± 17 | 66 ± 15 | 5 |
| CON | 75 ± 15 | 67 ± 18 | 65 ± 14 | 10 |
| Leptin (pg·mL−1) RT + PRO | 20.2 ± 22.4 | 17.5 ± 17.8 | 27.6 ± 44.6 | 7.4 |
| CON | 25.4 ± 30.2 | 27.3 ± 33.9 | 22.6 ± 27.4 | −2.8 |
| Insulin (µIU·mL−1) RT + PRO | 13.2 ± 5.2 | 14.7 ± 6.6 | 16.4 ± 12.4 | 3.2 |
| CON | 19.4 ± 16.6 | 22.9 ± 22.0 | 18.2 ± 15.4 | −1.2 |
| Baseline | Week 6 | Week 12 | Δ from Baseline to Week 12 | |
|---|---|---|---|---|
| Subjective appetite (mm·min−1) | ||||
| RT + PRO | 44 ± 15 | 47 ± 12 | 52 ± 14 | 8 * |
| CON | 39 ± 14 | 39 ± 17 | 44 ± 17 | 5 |
| Leptin (pg·mL·min−1) | ||||
| RT + PRO | 21.4 ± 27.9 | 15.6 ± 15.3 | 19.4 ± 20.4 | −2.0 |
| CON | 16.8 ± 17.7 | 14.8 ± 12.3 | 15.3 ± 15.7 | −1.5 |
| Insulin (µIU·mL·min−1) | ||||
| RT + PRO | 43.0 ± 11.6 | 48.6 ± 17.2 | 42.9 ± 14.3 | −0.1 |
| CON | 50.6 ± 17.2 | 47.6 ± 15.2 | 45.0 ± 20.6 | −4.4 |
| Baseline | Week 12 | Δ from Baseline to Week 12 | |
|---|---|---|---|
| Energy Intake (kcal) RT + PRO (total) RT + PRO (-protein supplement) CON | 1987 ± 396 1987 ± 396 2234 ± 634 | 2120 ± 533 1953 ± 507 2010 ± 448 | +133 +34 −224 |
| Protein Intake (g) | |||
| RT + PRO (total) RT + PRO (-protein supplement) CON Carbohydrate Intake (g) RT + PRO (total) RT + PRO (-protein supplement) CON Fat Intake (g) RT + PRO (total) RT + PRO (-protein supplement) CON | 85 ± 21 85 ± 21 85 ± 18 228 ± 70 228 ± 70 235 ± 54 88 ± 17 88 ± 17 112 ± 34 | 103 ± 26 73 ± 20 77 ± 19 214 ± 78 210 ± 92 262 ± 65 101 ± 28 97 ± 19 80 ± 17 | +18 −12 −8 −14 −18 +27 +13 +9 −32 |
| Analysis | F | η2 | Δ η2 | p-Value |
|---|---|---|---|---|
| One-way ANOVA | ||||
| Group | 12.79 * | 0.257 | - | 0.01 |
| One-way ANCOVA, group effect adjusting for | ||||
| FFM | 4.85 * | 0.119 | 0.138 | 0.034 |
| Baseline | Week 12 | Δ from Baseline to Week 12 (kg) | Effect Size (d) | |
|---|---|---|---|---|
| Seated Row RT + PRO CON | 37.2 ± 12.3 33.5 ± 14.1 | 49.8 ± 17.0 33.4 ± 14.1 | 12.6 −0.1 | 1.05 |
| Leg Press RT + PRO CON | 97.8 ± 26.4 95.1 ± 38.8 | 151.9 ± 29.2 97.5 ± 30.8 | 54.1 0.0 | 0.78 |
| Chest Press RT + PRO CON | 33.5 ± 18.1 25.7 ± 17.6 | 55.6 ± 24.7 28.7 ± 17.9 | 22.1 0.0 | 0.98 |
| Leg Curl RT + PRO CON | 43.7 ± 16.7 43.8 ± 17.3 | 70.9 ± 19.7 40.8 ± 17.3 | 27.2 −3.0 | 1.30 |
| Lat Pull Down RT + PRO CON | 35.2 ± 12.8 35.7 ± 12.2 | 48.2 ± 15.3 32.7 ± 12.2 | 13.0 3.0 | 0.88 |
| Leg Extension RT + PRO CON | 45.0 ± 16.8 43.4 ± 18.3 | 76.7 ± 20.3 47.6 ± 20.8 | 31.7 4.2 | 1.16 |
| Shoulder Press RT + PRO CON | 21.4 ± 14.6 15.9 ± 10.1 | 39.5 ± 20.3 18.5 ± 23.6 | 18.1 2.6 | 0.80 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, K.O.; Holliday, A.; Mistry, N.; Cunniffe, A.; Howard, K.; Stanger, N.; O’Mahoney, L.L.; Matu, J.; Ispoglou, T. An Increase in Fat-Free Mass is Associated with Higher Appetite and Energy Intake in Older Adults: A Randomised Control Trial. Nutrients 2021, 13, 141. https://doi.org/10.3390/nu13010141
Johnson KO, Holliday A, Mistry N, Cunniffe A, Howard K, Stanger N, O’Mahoney LL, Matu J, Ispoglou T. An Increase in Fat-Free Mass is Associated with Higher Appetite and Energy Intake in Older Adults: A Randomised Control Trial. Nutrients. 2021; 13(1):141. https://doi.org/10.3390/nu13010141
Chicago/Turabian StyleJohnson, Kelsie Olivia, Adrian Holliday, Nathan Mistry, Andrew Cunniffe, Kieran Howard, Nicholas Stanger, Lauren L. O’Mahoney, Jamie Matu, and Theocharis Ispoglou. 2021. "An Increase in Fat-Free Mass is Associated with Higher Appetite and Energy Intake in Older Adults: A Randomised Control Trial" Nutrients 13, no. 1: 141. https://doi.org/10.3390/nu13010141
APA StyleJohnson, K. O., Holliday, A., Mistry, N., Cunniffe, A., Howard, K., Stanger, N., O’Mahoney, L. L., Matu, J., & Ispoglou, T. (2021). An Increase in Fat-Free Mass is Associated with Higher Appetite and Energy Intake in Older Adults: A Randomised Control Trial. Nutrients, 13(1), 141. https://doi.org/10.3390/nu13010141