1. Introduction
Ulcerative colitis (UC) is an idiopathic inflammatory bowel disorder of the colon that causes dysregulation of gut microbiota and continuous mucosal inflammation. UC exhibits a higher prevalence worldwide, reaching 7.6–245 cases per 100,000 persons/year [
1]. Genetics, environmental factors, autoimmunity and gut microbiota were considered to increase the risk of UC [
2]. Given the complexity and appearing complications induced by immunosuppressive drugs, it is significant to developing novel therapeutic approaches and alternative drugs to treat UC.
Some probiotics, as nutritional supplements in functional foods, have been shown to promote intestinal health by enhancing epithelial function and reshaping gut microbial homeostasis. It was reported that
Bifidobacterium infantis 35624 could exert beneficial immunoregulatory effects in the mucosal immune system [
3].
Lactobacillus rhamnosus GG (
L. rhamnosus GG, ATCC 53103) could enhance intestinal functional maturation and IgA production and protect against colitis [
4].
Lactobacillus delbrueckii ssp
bulgaricus (LOT No. FK0201, LB-G040) attenuated the clinical signs of intestinal inflammation inducing a decrease in inflammatory cytokines [
5]. In addition, prebiotics might change the composition of gut microbiota, improve the function of the intestinal barrier, enhance intestinal immunity. It was shown that prebiotic fructans and resveratrol treatment could increase the amount of
Bifidobactrium and
Lactobacillus in DSS-induced colitis [
6]. Therefore, prebiotics and probiotics may represent a valid armamentarium to alleviate colitis, while the mechanism of the action is still unclear. In 2006, a clinical trial had been documented that
L. rhamnosus GG could be effective and safe for maintaining remission in patients with ulcerative colitis [
7]. However, the definite mechanism of
L. rhamnosus GG initiating anti-inflammatory activity remains incompletely illuminated.
The latest study showed that extracellular vesicles (EVs) played a crucial role in bacteria–bacteria and bacteria–host interactions [
8]. EVs derived from bacteria are spherical and nano-sized vesicles, and the diameter ranges from 20 to 400 nm. Like mammalian EVs, probiotic-derived EVs carry a large amount of microRNA, mRNA, proteins, and other active factors, which can mediate communications of cell–microbiota–host [
9]. It was speculated that EVs derived by bacteria played a key role in physiological and pathological functions by transferring cargoes to recipient cells. The beneficial immunomodulatory effects of probiotic-derived EVs were studied based on in vivo or in vitro assays [
10].
A. muciniphila (ATCC BAA-835) derived EV was reported as a beneficial factor that ameliorated the production of pro-inflammatory IL-6 from colon epithelial cells [
11]. EVs derived from
Lactobacillus kefirgranum PRCC-1301 might have an anti-inflammatory effect on colitis by inhibiting the NF-κB pathway and improving intestinal barrier function [
12]. However, many enigmatic puzzles regarding the underlying functions of Gram-positive EVs still remain unclear.
Given the efficacy of L. rhamnosus GG in patients with ulcerative colitis, whether the EVs released from L. rhamnosus GG are involved in anti-inflammatory response and the mechanism remain unknown. In this study, the aim was to investigate the effect of L. rhamnosus GG-EVs (LGG-EVs) on anti-inflammatory properties based on the TLRs-NF-κB-NLRP3 signaling pathway and gut microbiota in DSS-induced colitis mice.
2. Materials and Methods
2.1. Preparation of L. rhamnosus GG Derived EVs
L. rhamnosus GG was purchased from ATCC (53103) and cultured in MRS medium at 37 °C for 24 h. Then medium was centrifuged at 8000 × g for 30 min at 4 °C to remove cells and other debris. The supernatant was filtered by 0.22 μm PVDF filter and concentrated using Centricon™ Plus-70 Centrifugal Filter Units (MilliporeSigma™). The filtered suspension was ultracentrifuged at 100,000 g using HIMAC CP70ME Ultracentrifuge (Hitachi, Ltd., Tokyo, Japan) at 4 °C for 2 h. The pellets were resuspended with PBS, and washed by ultracentrifugation at 100,000 × g for 60 min at 4 °C. LGG-EVs were collected and stored at −80 °C. The size distribution of LGG-EVs was measured by Dynamic Light Scattering (DLS), and all samples were evaluated in three replicates. The morphology of LGG-EVs was visualized via Transmission Electron Microscope (TEM). The morphology was imaged using HITACHI H-7650 Transmission Electron Microscope (Hitachi, Ltd., Tokyo, Japan).
2.2. Animals and Treatments
Specific pathogen-free (SPF) C57BL/6J male mice (4–5 weeks of age) were purchased from the Laboratory Animal Breeding Center of Pengyue (Jinan, China). Mice were maintained at 22 °C with 12-h light/dark cycles and kept in individual cages. A standard diet (
Table S1) and water were provided ad libitum. The relative humidity was 30% to 70% in the experimental room. Bedding-change and cage-washing was carried out frequently and the preparation of recycled air was used to reduce the stress associated with the experiment. After adaptation for 1 week, mice were randomly divided three groups, including the control group, DSS group (3.5% DSS), and treatment group (3.5% DSS + 1.2 mg/kg LGG-EVs of body weight). Specifically, LGG-EVs were pre-administered to mice for 2 weeks by daily gavage of LGG-EVs (1.2 mg/kg of body weight) according to the latest study [
13]. Control and DSS group mice were fed an equal volume of PBS. Then 3.5% DSS (Yeasen Biotech, Shanghai, China) were added into the water for inducing colitis. LGG-EVs were administered until the end of the experiment. The body weight and presence of blood in the stool were used to monitor the development of colitis. All experimental processes were approved by the Committee on the Ethics of Animal Experiments of Ocean University of China (permission number: spxy20200820215).
2.3. Western Blot Analysis
Total proteins were extracted from colonic tissues and LGG-EVs using RIPA buffer containing protease and phosphatase inhibitor (Beyotime Biotechnology, Shanghai, China). The equal quantity of protein was loaded into gels, and then transferred onto PVDF membranes (Thermo Fisher Scientific). Proteins were quantified by Image J software (NIH, Bethesda, MD). Antibodies against TSG101 (ab225877), p65 (ab16502), p-p65 (ab76302), NLRP3 (ab210491), ASC (ab180799) were purchased from Abcam (Cambridge, UK). β-actin (GB12001) was purchased from Servicebio (Wuhan servicebio technology CO., LTD, Wuhan, China).
2.4. Quantitative Real-Time PCR
Colonic tissues were employed to evaluate the relative expression of inflammatory cytokines. Total RNA was extracted using Trizol Reagent (TIANGEN Biotech, Beijing, China), and 1 μg of total RNA was reverse transcribed using High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Waltham, MA, USA). The expression of IL-6, TNF-α, IL-1β, IL-2, TLR-4, Myd88, and GAPDH genes were quantitated using Power Up SYBR Green MasterMix on a StepOnePlus Real-Time PCR Instrument (Applied Biosystems). GAPDH was used as a housing gene. Primers were included in
Supplementary Table S2.
2.5. Enzyme-Linked Immunosorbent Assay
The concentrations of TNF-α, IL-6, and IL-1β were determined in serum using ELISA kits according to the manufacturer’s instructions (Nanjing Jiancheng Bio, Nanjing, China). In brief, serum was obtained by centrifugation at 5000× g for 30 min at 4 °C. The absorbance was detected at 450 nm by Multiskan FC (Thermo Scientific, Waltham, MA, USA).
2.6. Tissue Histology
The colonic segments were taken and fixed using 4% paraformaldehyde. Then the paraffin sections of each colon were stained with haematoxylin and eosin (H&E). The colonic thin sections were observed under a light microscope (OLYMPUS) at a magnification of 100×.
2.7. Microbiota 16S rRNA Gene Sequencing
Total genomic DNA was extracted from the fecal samples of three groups. The bacterial 16S rRNA genes V3–V4 region was amplified using the forward primer 338F (5′-ACTCCTACGGGAGGCAGCA-3′) and the reverse primer 806R (5′-TCGGACTACHVGGGTWTCTAAT-3′). Then the amplicons were pooled in equal amounts, and pair-end 2 × 300 bp sequencing was performed using the Illlumina MiSeq platform with MiSeq Reagent Kit V3 at Shanghai Personal Biotechnology Co., Ltd. (Shanghai, China). High-quality sequences were clustered into OTUs at 97% sequence identity by UCLUST. Microbial functions were predicted by PICRUSt according to high-quality sequences. LEfSe was used to investigate bacterial members that drive differences between groups.
2.8. Statistical Analysis
Data were presented as the mean ± standard deviation (SD) or the median, min and max values according to the normality of distribution. One-way analysis of variance (ANOVA), Duncan test and Normality and Lognormality Tests (D’Agostino and Pearson test) was performed using SPSS 22 (SPSS Inc., Chicago, IL, USA) and GraphPad Prime 8 (GraphPad Software, San Diego, CA, USA). All other statistical tests were performed using the GraphPad Prime 8. Statistical differences were considered significant at p < 0.05.
4. Discussion
In this study, the effect of
L. rhamnosus GG-derived extracellular vesicles (LGG-EVs) on the DSS-induced colitis was investigated in the mice model. To date,
L. rhamnosus GG was used for clinical study in many gastrointestinal diseases (NCT01790035, NCT01773967). However, there is no uniform understanding of the mechanism of its probiotic function. The role of Gram-positive EVs on health and disease has been drawing more attention recently. Accumulating evidence has indicated that the probiotic effects of some probiotic bacteria may not depend on the bacteria being alive but on the metabolites released by probiotic bacteria, such as EVs or carried effector molecules in EVs [
31,
32]. The latest study demonstrated that the function of
L. rhamnosus GG was mediated through its nanoparticles (EVs), which protected against alcohol-associated liver disease through intestinal aryl hydrocarbon receptor [
13]. To our knowledge, the immunomodulatory effect of LGG-EVs on experimental colitis remains unknown. Hereby, a mouse colitis model was induced by DSS. Consistent with the treatment of
L. rhamnosus GG in the previous report [
7], LGG-EVs seems to be effective and safe for maintaining remission in colitis mouse. In addition, a single dose of LGG-EVs was used in this study, more doses should be chosen for comprehensively clarifying the anti-inflammatory effect of LGG-EVs in subsequent research.
It is confirmed that TLR4 signaling is critical for colon carcinogenesis in colitis, and the inhibition of TLR4 signaling may be useful for protecting against colitis or colitis-associated cancer [
33]. NF-κB, the key regulator in the mucosal immune system, could be activated by upstream stimuli TLR4. Therefore, inhibiting TLR4 signaling can effectively regulate the expression levels of inflammatory cytokines induced by DSS. It had been reported that
L. rhamnosus GG or
L. rhamnosus GG-derived factors could attenuate the inflammatory response induced by activation of TLR4/NF-κB in the colitis [
34,
35]. In our current research, we found LGG-EVs had the same anti-inflammatory effect, which was accomplished by downregulating the expression of myd88, p65, and p-p65 via inhibiting TLR4 signaling. Accordingly, the expression of pro-inflammatory cytokines at the gene and protein levels were decreased in DSS-induced colitis mice treated with LGG-EVs. NLRP3, the member of the NLRs family, is rapidly emerging as a crucial regulator of the intestinal homeostasis, which could induce production of IL-1β and facilitate the activation of NF-κB signaling cascades, and then initiate the immune response [
36]. Interestingly, Toll-like receptors (TLRs) could induce the synthesis of mitochondrial DNA, which is crucial for NLRP3 signaling. Taken together, a closely TLRs-NF-κB-NLRP3 network was formed and might play a key role in the pathogenesis of colitis. Given the role of LGG-EVs in the TLRs-NF-κB-NLRP3 signaling pathway, it was considered that LGG-EVs might be involved in this network to improve colitis. However, whether LGG-EVs could target TLRs and induce the synthesis of mitochondrial DNA, and activate NLRP3 signaling still needs to be further studied.
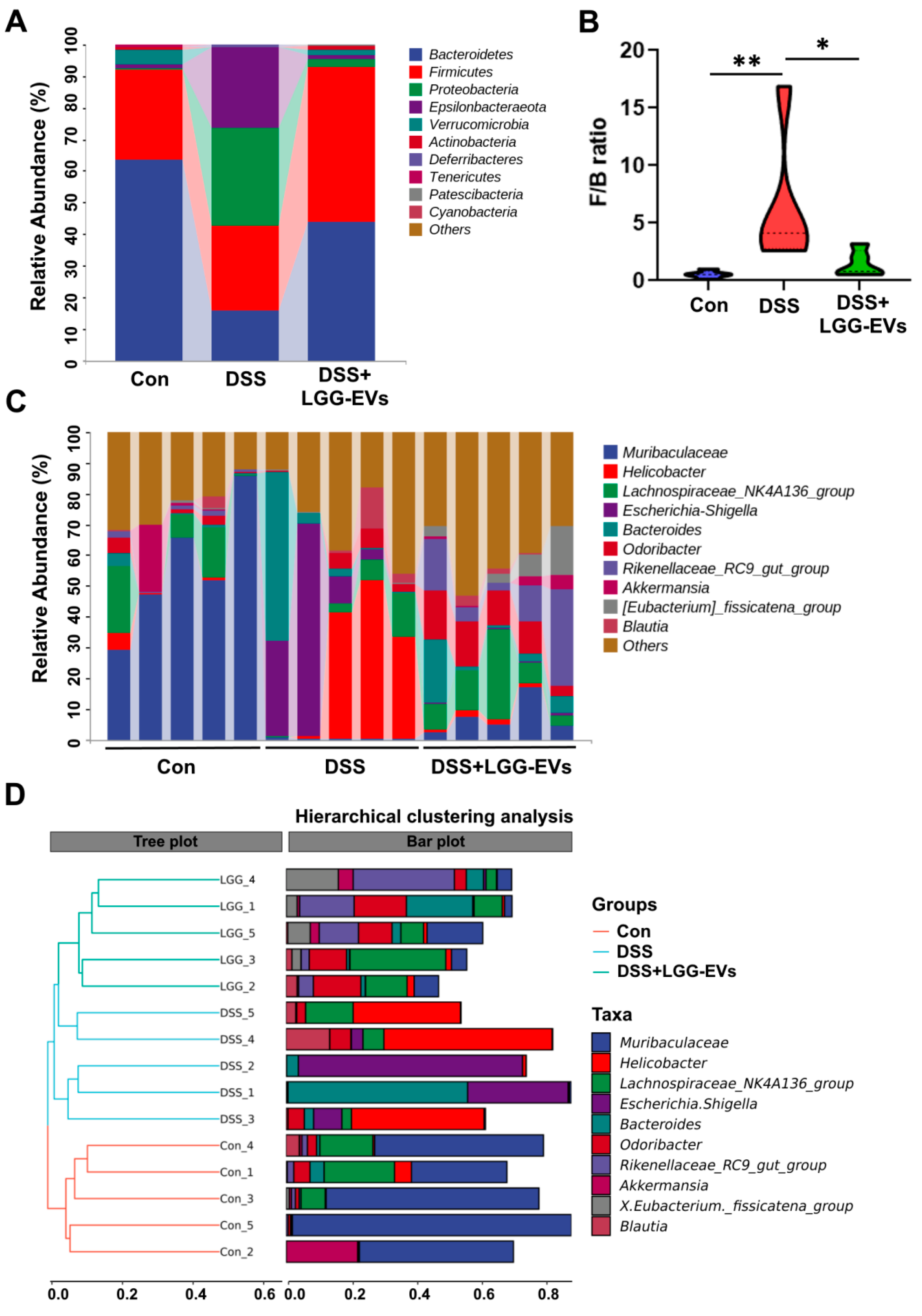
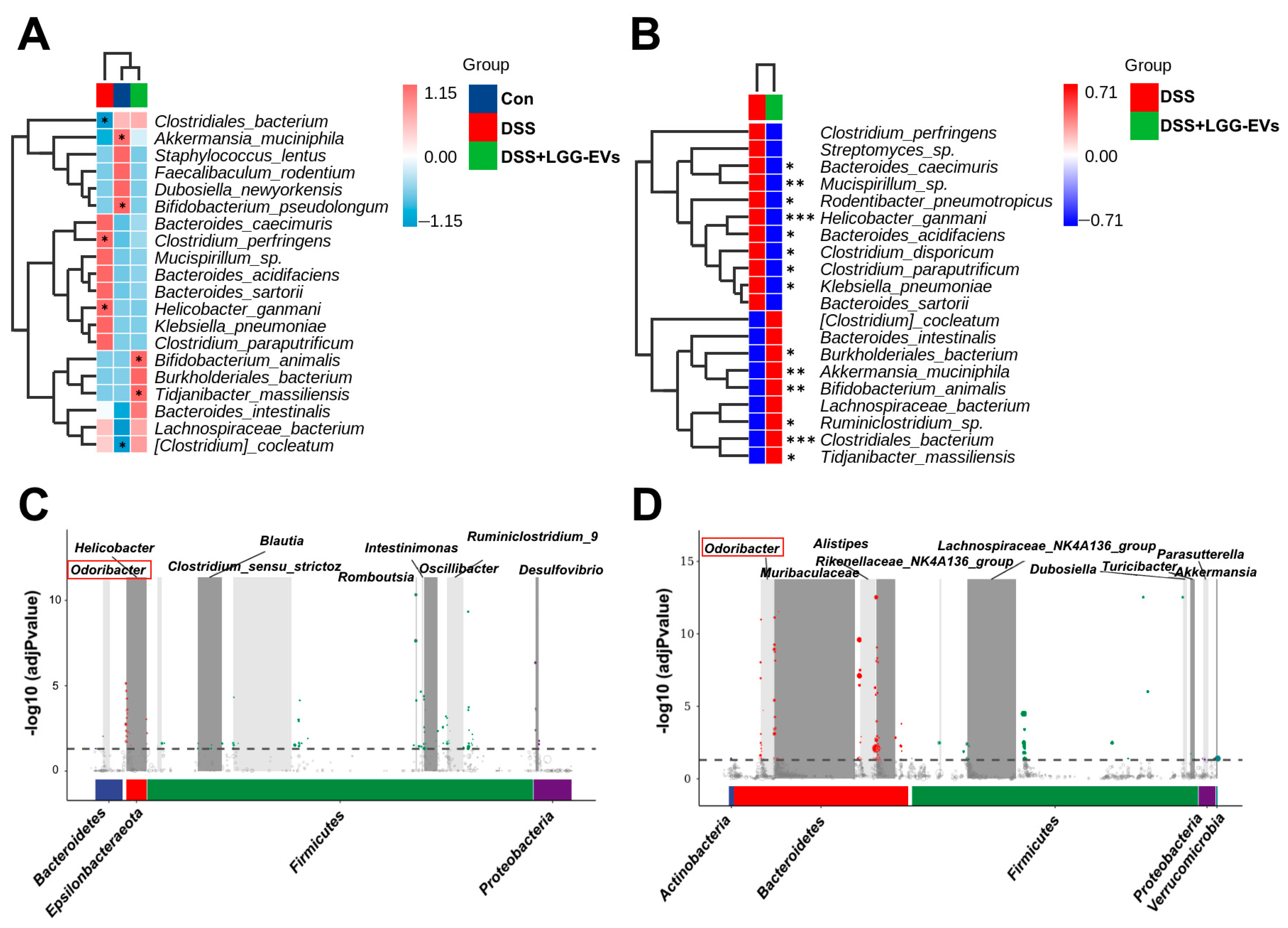
In general, the pathogenesis of colitis is closely related to the gut microbiota, although the bacteria contributing to colitis have not been determined [
37]. Probiotics are emerging as functional foods to manipulate the gut microbiota and suppress the growth of pathogens [
38]. However, the proposed mechanisms of probiotics on the intestinal microbiome are incomplete presently. Some studies documented that probiotics as living microorganisms belonged to the part of gut microbiota, and promoted beneficial effects when ingested in adequate amounts [
39]. Other researches indicated that probiotics produced metabolic compounds and inhibited the growth of other microorganisms [
40]. A recent study reported that the microbiota-derived EVs might play significant roles in host–helminth–microbiome crosstalk [
41]. Therefore, it is well recognized that the probiotic-derived EVs may be an overlooked mediator in host–microbiome–probiotic interactions.
Numerous studies were focused on the effects of EVs derived from various foods on gut microbiota [
42,
43]. Teng et al. showed that plant-derived EVs regulated gut microbiota composition and localization, and enhanced the gut barrier function to alleviate colitis [
44]. It was documented in our previous research that milk-derived EVs could alleviate colitis via modulating gut microbiota [
45]. Here, we found that LGG-EVs restored the disorders of α- and β-diversity induced by DSS. Indeed, colitis has been associated with dysbiosis, defined as a significant decrease in gut microbial diversity [
2]. The changes of microbial diversity were accompanied by the shift between commensal and pathogenic microorganisms, for example, the incensement of
Firmicutes:
Bacteroidetes (
F/B) ratio in the DSS group. Consistently, the members of
Proteobacteria phylum, including
Enterobacteriaceae, Escherichia_Shigella, were commonly increased in colitis compared to the healthy and LGG-EVs treated mice, which were consistent with the results of a previous report [
46]. Furthermore, LGG-EVs treatment significantly upregulated the relative abundance of
Akkermansia_muciniphila, Bifidobacterium_animalis at the species level. Emerging evidence has indicated a close negative correlation between
Akkermansia_muciniphila abundance and obesity, gut inflammation, or hypertension [
47,
48].
Bifidobacterium_animalis, as a generally recognized probiotic, plays a key role in gastrointestinal health and immune function [
49]. In a line with the previous reports, both
Akkermansia_muciniphila and
Bifidobacterium_animalis were closely correlated with the valine, leucine and isoleucine degradation pathway and fluorobenzoate degradation [
30,
50], which were regulated upon LGG-EVs at different level. The results demonstrated that the LGG-EVs could regulate the microbial metabolic pathway via affecting the special bacteria.
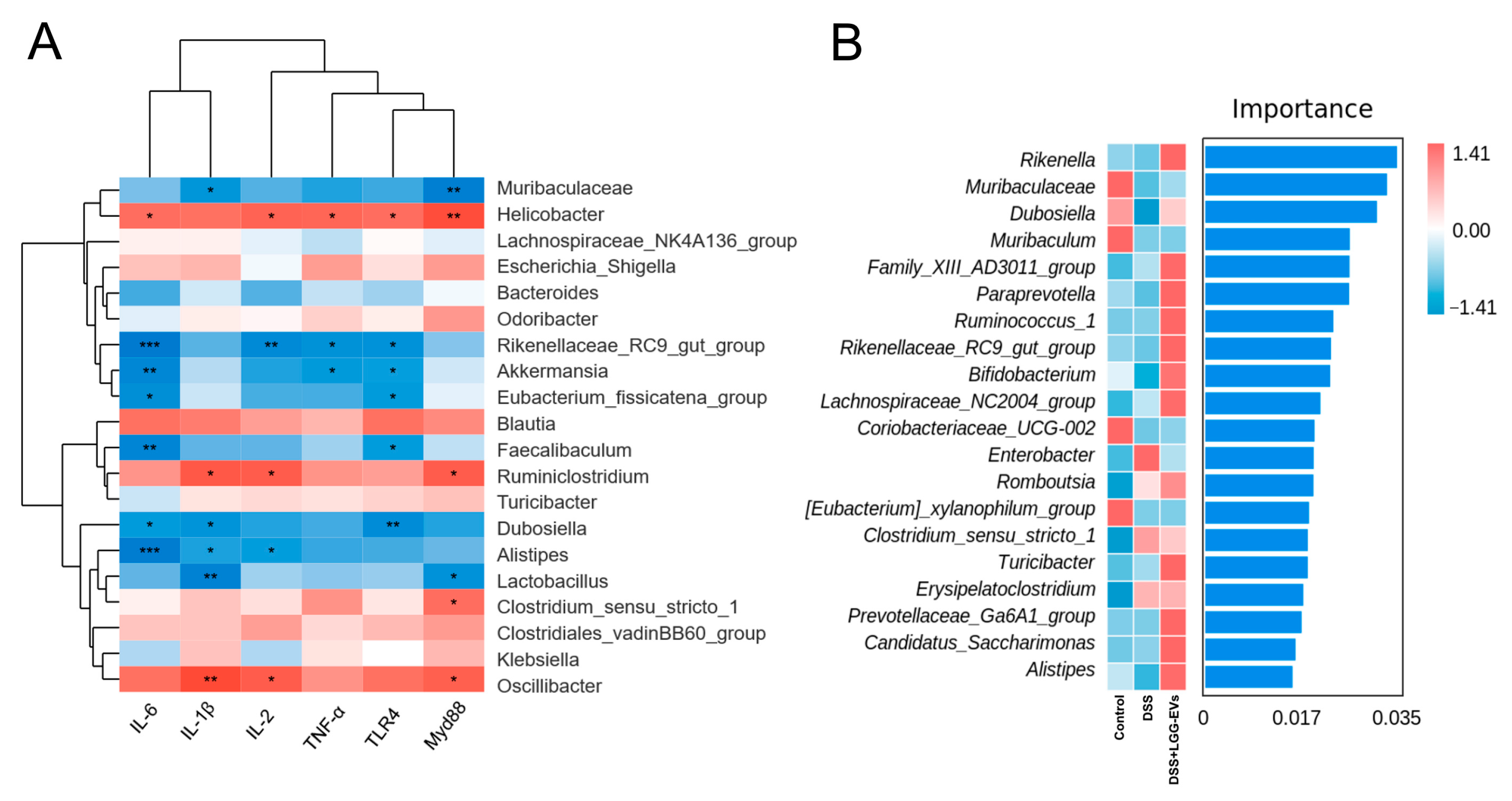
It had been suggested that TLRs and NLRs could contribute to shaping the composition of the gut microbiota. Dheer et al. indicated that the expression of epithelial TLR4 shaped the microbiota and affected the functional properties of the epithelium [
51]. The composition of gut microbiota and increased inflammation depends on the expression of TLR4 in colon tissues [
52]. Moreover, there was lower bacteria abundance in the NLRP3
−/− mice colon compared to that of the WT mice. NLRP3 inflammasome is emerging to be a crucial mediator of gut homeostasis by modulating immune responses to microbiota in the gut [
36]. In this study, we found that LGG-EVs could act not only as a key mediator of TLRs-NF-κB-NLRP3 but also reshape the construction and composition of gut microbiota. Namely, the abundance of
Muribaculaceae, Akkermansia, Faecalibaculum, Alistipes, Lactobcillus had a significant negative association with pro-inflammatory cytokines, such as IL-6, TNF-α, TLR4, etc. It was demonstrated that
Akkermansia, Faecalibaculum, Alistipes could inhibit the production of pro-inflammatory factors and play the potential beneficial effect on intestinal immune homeostasis [
48,
53]. However, whether LGG-EVs could mediate the TLRs-NF-κB-NLRP3 signaling pathway via regulating gut microbiota to alleviate colitis remains to be further explored.
To date, almost all research has focused on the characterization and function of EVs. In fact, EVs contain amounts of functional cargos, including RNA, proteins and etc. Further studies are required to explore which molecules play a crucial role in intestinal immune homeostasis and host health.


 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}