Glycemic Control and Metabolic Adaptation in Response to High-Fat versus High-Carbohydrate Diets—Data from a Randomized Cross-Over Study in Healthy Subjects

 , , ,
, , , 
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Setting, Participants and Study Design
2.2. Enrolment and Randomization
2.3. Diets and Study Instructions
2.4. Mixed Meal Test
2.5. Biochemistry
2.6. Glucose, Insulin, GLP-1 and GIP Quantifications
2.7. Metabolomics Sample Preparation and NMR Data Acquisition and Processing
2.8. Statistics
3. Results
3.1. The Study Group and General Responses
3.2. Appetite Scoring during the Mixed Meal Tests
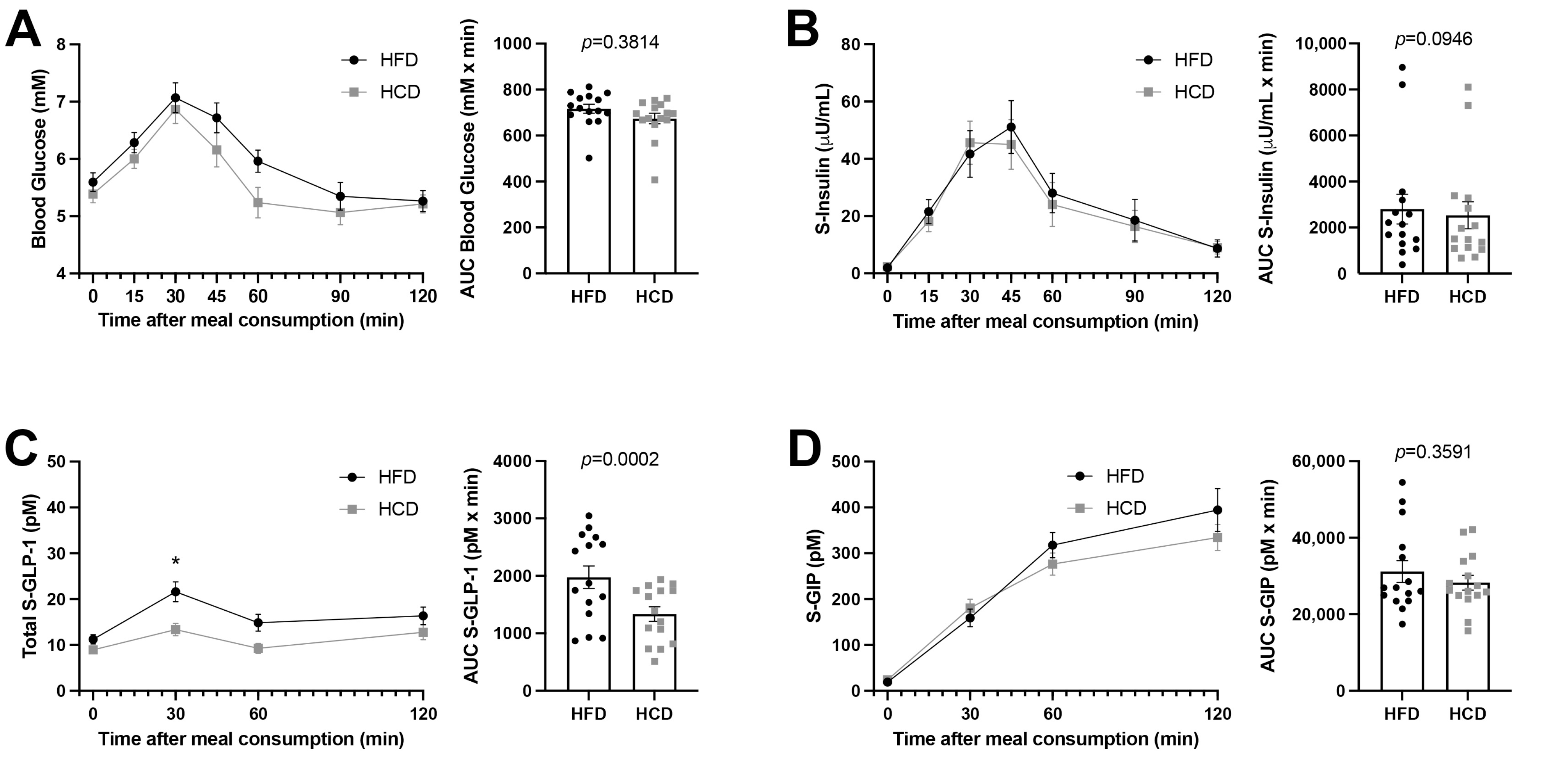
3.3. Glucose and Insulin Levels as Well as Incretins in Blood following the Mixed Meal Tests
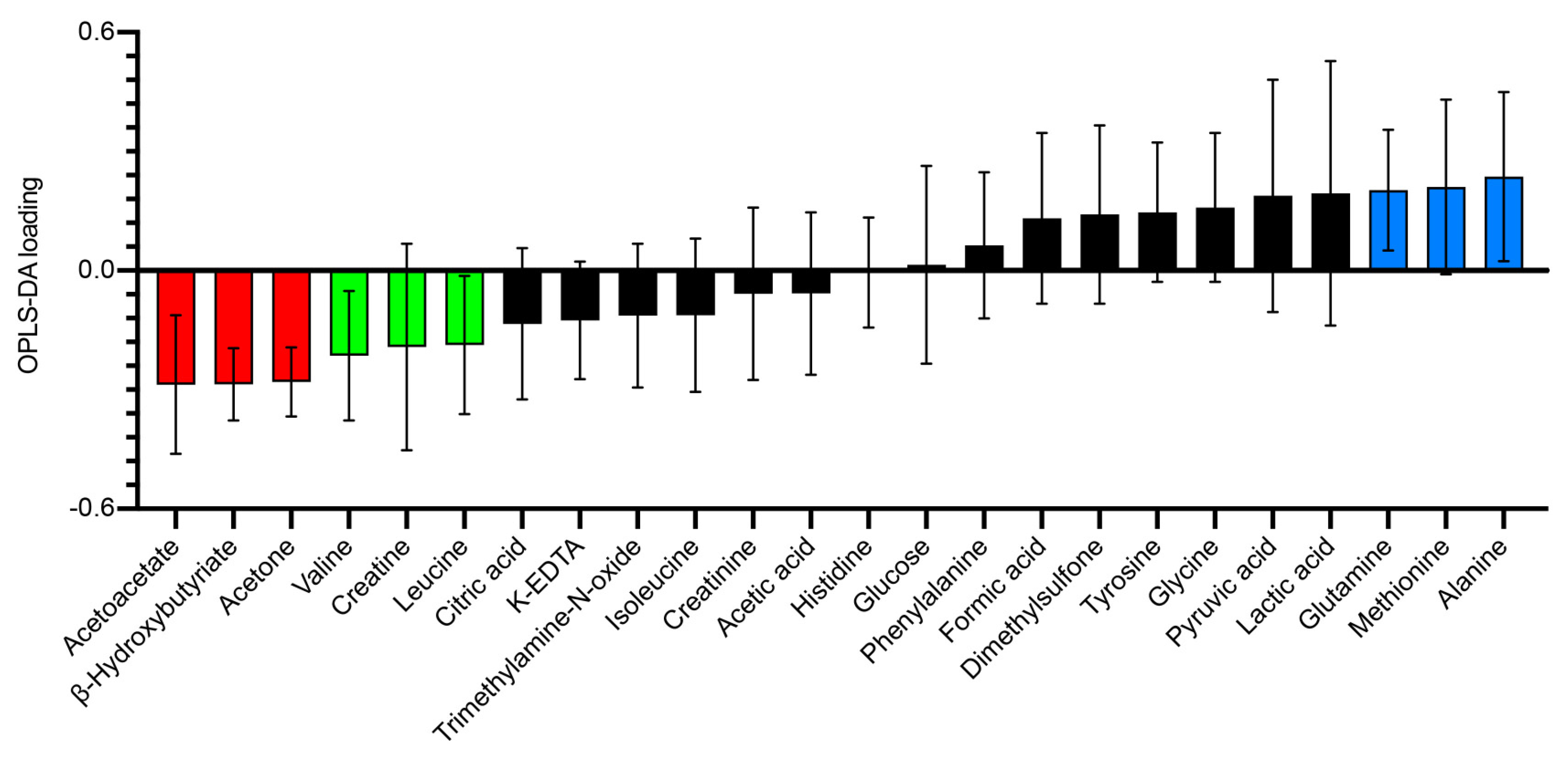
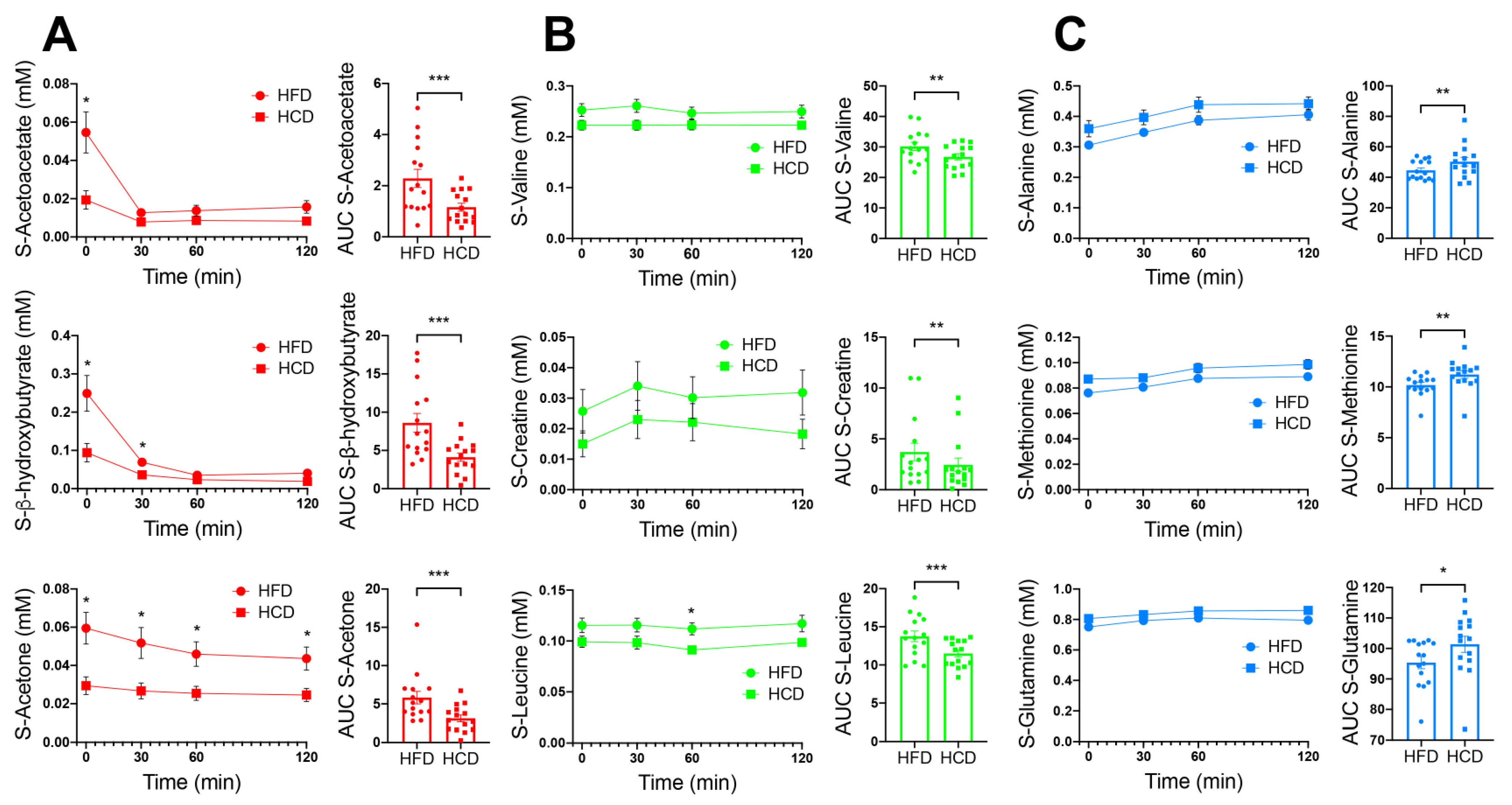
3.4. Other Systemic Metabolites Indicating Risk for a Diabetogenic Effect of High-Fat Diet
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Astrup, A.; Hjorth, M.F. Low-Fat or Low Carb for Weight Loss? It Depends on Your Glucose Metabolism. EBioMedicine 2017, 22, 20–21. [Google Scholar] [CrossRef] [Green Version]
- van Wyk, H.J.; Davis, R.E.; Davies, J.S. A critical review of low-carbohydrate diets in people with Type 2 diabetes. Diabet. Med. 2016, 33, 148–157. [Google Scholar] [CrossRef]
- Wan, Y.; Wang, F.; Yuan, J.; Li, J.; Jiang, D.; Zhang, J.; Huang, T.; Zheng, J.; Mann, J.; Li, D. Effects of Macronutrient Distribution on Weight and Related Cardiometabolic Profile in Healthy Non-Obese Chinese: A 6-month, Randomized Controlled-Feeding Trial. EBioMedicine 2017, 22, 200–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helander, H.F.; Fandriks, L. Surface area of the digestive tract—Revisited. Scand. J. Gastroenterol. 2014, 49, 681–689. [Google Scholar] [CrossRef]
- Lynch, C.J.; Adams, S.H. Branched-chain amino acids in metabolic signalling and insulin resistance. Nat. Rev. Endocrinol. 2014, 10, 723–736. [Google Scholar] [CrossRef] [Green Version]
- Felig, P.; Marliss, E.; Cahill, G.F., Jr. Plasma amino acid levels and insulin secretion in obesity. N. Engl. J. Med. 1969, 281, 811–816. [Google Scholar] [CrossRef]
- Lotta, L.A.; Scott, R.A.; Sharp, S.J.; Burgess, S.; Luan, J.; Tillin, T.; Schmidt, A.F.; Imamura, F.; Stewart, I.D.; Perry, J.R.B.; et al. Genetic Predisposition to an Impaired Metabolism of the Branched-Chain Amino Acids and Risk of Type 2 Diabetes: A Mendelian Randomisation Analysis. PLoS Med. 2016, 13, e1002179. [Google Scholar] [CrossRef]
- Neinast, M.; Jang, C.; Hui, S.; Murashige, D.S.; Chu, Q.; Morscher, R.J.; Li, X.; Zhan, L.; White, E.; Anthony, T.G.; et al. Quantitative Analysis of the Whole-Body Metabolic Fate of Branched-Chain Amino Acids. Cell Metab. 2019, 29, 417–429.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vangipurapu, J.; Stancakova, A.; Smith, U.; Kuusisto, J.; Laakso, M. Nine Amino Acids Are Associated With Decreased Insulin Secretion and Elevated Glucose Levels in a 7.4-Year Follow-up Study of 5181 Finnish Men. Diabetes 2019, 68, 1353–1358. [Google Scholar] [CrossRef]
- Wang, T.J.; Larson, M.; Vasan, R.S.; Cheng, S.; Rhee, E.P.; McCabe, E.; Lewis, G.D.; Fox, C.S.; Jacques, P.F.; Fernandez, C.; et al. Metabolite profiles and the risk of developing diabetes. Nat. Med. 2011, 17, 448–453. [Google Scholar] [CrossRef]
- Mifflin, M.D.; St Jeor, S.T.; Hill, L.A.; Scott, B.J.; Daugherty, S.A.; Koh, Y.O. A new predictive equation for resting energy expenditure in healthy individuals. Am. J. Clin. Nutr. 1990, 51, 241–247. [Google Scholar] [CrossRef]
- Beirnaert, C.; Meysman, P.; Vu, T.N.; Hermans, N.; Apers, S.; Pieters, L.; Covaci, A.; Laukens, K. speaq 2.0: A complete workflow for high-throughput 1D NMR spectra processing and quantification. PLoS Comput. Biol. 2018, 14, e1006018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thevenot, E.A.; Roux, A.; Xu, Y.; Ezan, E.; Junot, C. Analysis of the Human Adult Urinary Metabolome Variations with Age, Body Mass Index, and Gender by Implementing a Comprehensive Workflow for Univariate and OPLS Statistical Analyses. J. Proteome Res. 2015, 14, 3322–3335. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Westerhuis, J.A.; Rosen, J.; Landberg, R.; Brunius, C. Variable selection and validation in multivariate modelling. Bioinformatics 2019, 35, 972–980. [Google Scholar] [CrossRef] [PubMed]
- Holst, J.J. The incretin system in healthy humans: The role of GIP and GLP-1. Metabolism 2019, 96, 46–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herzberg-Schafer, S.; Heni, M.; Stefan, N.; Haring, H.U.; Fritsche, A. Impairment of GLP1-induced insulin secretion: Role of genetic background, insulin resistance and hyperglycaemia. Diabetes Obes. Metab. 2012, 14, 85–90. [Google Scholar] [CrossRef]
- Wallenius, V.; Elias, E.; Elebring, E.; Haisma, B.; Casselbrant, A.; Larraufie, P.; Spak, E.; Reimann, F.; Le Roux, C.W.; Docherty, N.; et al. Suppression of enteroendocrine cell glucagon-like peptide (GLP)-1 release by fat-induced small intestinal ketogenesis: A mechanism targeted by Roux-en-Y gastric bypass surgery but not by preoperative very-low-calorie diet. Gut 2020, 69, 1423–1431. [Google Scholar] [CrossRef] [Green Version]
- Wallenius, V.; Dirinck, E.; Fandriks, L.; Maleckas, A.; le Roux, C.W.; Thorell, A. Glycemic Control after Sleeve Gastrectomy and Roux-En-Y Gastric Bypass in Obese Subjects with Type 2 Diabetes Mellitus. Obes. Surg. 2018, 28, 1461–1472. [Google Scholar] [CrossRef] [Green Version]
- Neinast, M.; Murashige, D.; Arany, Z. Branched Chain Amino Acids. Annu. Rev. Physiol. 2019, 81, 139–164. [Google Scholar] [CrossRef]
- Sun, Y.; Gao, H.Y.; Fan, Z.Y.; He, Y.; Yan, Y.X. Metabolomics Signatures in Type 2 Diabetes: A Systematic Review and Integrative Analysis. J. Clin. Endocrinol. Metab. 2020, 105, 1000–1008. [Google Scholar] [CrossRef]
- Post, A.; Groothof, D.; Schutten, J.C.; Flores-Guerrero, J.L.; Swarte, J.C.; Douwes, R.M.; Kema, I.P.; de Boer, R.A.; Garcia, E.; Connelly, M.A.; et al. Plasma creatine and incident type 2 diabetes in a general population-based cohort: The PREVEND study. Clin. Endocrinol. 2021, 94, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Vogelzangs, N.; Van Der Kallen, C.J.H.; Van Greevenbroek, M.M.J.; Van Der Kolk, B.W.; Jocken, J.W.E.; Goossens, G.; Schaper, N.C.; Henry, R.M.A.; Eussen, S.J.P.M.; Valsesia, A.; et al. Metabolic profiling of tissue-specific insulin resistance in human obesity: Results from the Diogenes study and the Maastricht Study. Int. J. Obes. 2020, 44, 1376–1386. [Google Scholar] [CrossRef] [PubMed]
- Cobb, J.; Eckhart, A.; Motsinger-Reif, A.; Carr, B.; Groop, L.; Ferrannini, E. alpha-Hydroxybutyric Acid Is a Selective Metabolite Biomarker of Impaired Glucose Tolerance. Diabetes Care 2016, 39, 988–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrannini, E.; Natali, A.; Camastra, S.; Nannipieri, M.; Mari, A.; Adam, K.-P.; Milburn, M.V.; Kastenmüller, G.; Adamski, J.; Tuomi, T.; et al. Early Metabolic Markers of the Development of Dysglycemia and Type 2 Diabetes and Their Physiological Significance. Diabetes 2012, 62, 1730–1737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gall, W.E.; Beebe, K.; Lawton, K.A.; Adam, K.-P.; Mitchell, M.; Nakhle, P.J.; Ryals, J.A.; Milburn, M.V.; Nannipieri, M.; Camastra, S.; et al. alpha-Hydroxybutyrate Is an Early Biomarker of Insulin Resistance and Glucose Intolerance in a Nondiabetic Population. PLoS ONE 2010, 5, e10883. [Google Scholar] [CrossRef] [Green Version]
- Goodman, R.P.; Markhard, A.L.; Shah, H.; Sharma, R.; Skinner, O.S.; Clish, C.B.; Deik, A.; Patgiri, A.; Hsu, Y.-H.H.; Masia, R.; et al. Hepatic NADH reductive stress underlies common variation in metabolic traits. Nature 2020, 583, 122–126. [Google Scholar] [CrossRef]
- DeFronzo, R.A.; Ferrannini, E. Insulin resistance. A multifaceted syndrome responsible for NIDDM, obesity, hypertension, dyslipidemia, and atherosclerotic cardiovascular disease. Diabetes Care 1991, 14, 173–194. [Google Scholar] [CrossRef]
- Shantavasinkul, P.C.; Muehlbauer, M.J.; Bain, J.R.; Ilkayeva, O.R.; Craig, D.M.; Newgard, C.B.; Svetkey, L.P.; Shah, S.H.; Torquati, A. Improvement in insulin resistance after gastric bypass surgery is correlated with a decline in plasma 2-hydroxybutyric acid. Surg. Obes. Relat. Dis. 2018, 14, 1126–1132. [Google Scholar] [CrossRef]
- Brown-Borg, H.M.; Buffenstein, R. Cutting back on the essentials: Can manipulating intake of specific amino acids modulate health and lifespan? Ageing Res. Rev. 2017, 39, 87–95. [Google Scholar] [CrossRef]
- Liu, X.; Gao, X.; Zhang, R.; Liu, Z.; Shen, N.; Di, Y.; Fang, T.; Li, H.; Tian, F. Discovery and comparison of serum biomarkers for diabetes mellitus and metabolic syndrome based on UPLC-Q-TOF/MS. Clin. Biochem. 2020, 82, 40–50. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scoring before Meal | Change from Baseline | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 30 min after Start of Meal | 120 min after Start of Meal | ||||||||
| Sensation | HCD | HFD | p | HCD | HFD | p | HCD | HFD | p |
| Craving | 48 (6.9) | 67 (5.8) | 0.021 | 21 (4.7) | 19 (4.0) | 0.44 | 39 (7.8) | 22 (6.0) | 0.48 |
| Hunger | 54 (6.6) | 64 (6.4) | 0.12 | 26 (5.7) | 17 (3.8) | 0.036 | 41 (7.3) | 33 (6.0) | 0.091 |
| Satiation | 16 (4.3) | 13 (4.1) | 0.37 | 60 (5.2) | 68 (4.7) | 0.14 | 45 (4.8) | 56 (5.3) | 0.092 |
| Baseline Data | Post-Treatment Data | |||
|---|---|---|---|---|
| HCD | HFD | p | ||
| No. of randomized participants | 16 | 15 | 15 | |
| Gender (f/m) | 8/8 | 7/8 | 7/8 | |
| Age (years) | 26.1 (0.8) | |||
| Body weight (kg) | 72.3 (3.0) | 72.5 (3.0) | 71.7 (2.9) | 0.009 |
| B-hemoglobin (g/L) | 137 (2.9) | 135 (2.9) | 139 (2.7) | 0.045 |
| White cell count (109/L) | 5.0 (0.3) | 5.3 (0.5) | 4.8 (0.2) | ns |
| CRP (mg/L) | 0.13 (0.09) | 0 (0) | 0.07 (0.07) | ns |
| Triacylglycerol (mmol/L) | 0.93 (0.06) | 0.96 (0.11) | 0.70 (0.07) | <0.001 |
| HDL (mmol/L) | 1.72 (0.12) | 1.47 (0.11) | 1.82 (0.13) | <0.001 |
| LDL (mmol/L) | 2.94 (0.22) | 2.42 (0.18) | 2.79 (0.20) | 0.003 |
| LDL/HDL | 1.75 (0.18) | 1.77 (0.18) | 1.65 (0.17) | ns |
| Total cholesterol | 4.69 (0.88) | 4.07 (0.72) | 4.74 (0.88) | <0.001 |
| ASAT (µkat/L) | 0.44 (0.02) | 0.41 (0.04) | 0.45 (0.07) | ns |
| ALAT (µkat/L) | 0.33 (0.02) | 0.31 (0.03) | 0.33 (0.03) | ns |
| ALP (µkat/L) | 1.03 (0.07) | 0.95 (0.07) | 0.94 (0.07) | ns |
| Bilirubin (µmol/L) | 10.6 (0.95) | 10.9 (1.66) | 10.2 (1.20) | ns |
| Na+(mmol/L) | 140 (0.63) | 139 (0.36) | 140 (0.34) | ns |
| K+ (mmol/L) | 4.2 (0.07) | 4.1 (0.05) | 4.0 (0.04) | ns |
| Creatinine (µmol/L) | 80 (2.3) | 84 (2.60) | 85 (3.20) | ns |
| Metabolite | Diet | Correlation with | r | p |
|---|---|---|---|---|
| Fasting value | ||||
| Acetone | HFD | HOMA-IR | −0.59 | 0.0227 |
| Acetone | HCD | HOMA-IR | −0.59 | 0.0230 |
| β-hydroxybutyric acid | HCD | HOMA-IR | −0.53 | 0.0471 |
| AUC | ||||
| Acetoacetate | HFD | Insulin | −0.56 | 0.0335 |
| Alanine | HFD | GLP-1 | 0.74 | 0.0022 |
| α-hydroxybutyric acid | HFD | Insulin | −0.65 | 0.0107 |
| β-hydroxybutyric acid | HCD | GLP-1 | −0.62 | 0.0155 |
| Methionine | HCD | GIP | 0.62 | 0.0162 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wallenius, V.; Elebring, E.; Casselbrant, A.; Laurenius, A.; le Roux, C.W.; Docherty, N.G.; Biörserud, C.; Björnfot, N.; Engström, M.; Marschall, H.-U.; et al. Glycemic Control and Metabolic Adaptation in Response to High-Fat versus High-Carbohydrate Diets—Data from a Randomized Cross-Over Study in Healthy Subjects. Nutrients 2021, 13, 3322. https://doi.org/10.3390/nu13103322
Wallenius V, Elebring E, Casselbrant A, Laurenius A, le Roux CW, Docherty NG, Biörserud C, Björnfot N, Engström M, Marschall H-U, et al. Glycemic Control and Metabolic Adaptation in Response to High-Fat versus High-Carbohydrate Diets—Data from a Randomized Cross-Over Study in Healthy Subjects. Nutrients. 2021; 13(10):3322. https://doi.org/10.3390/nu13103322
Chicago/Turabian StyleWallenius, Ville, Erik Elebring, Anna Casselbrant, Anna Laurenius, Carel W. le Roux, Neil G. Docherty, Christina Biörserud, Niclas Björnfot, My Engström, Hanns-Ulrich Marschall, and et al. 2021. "Glycemic Control and Metabolic Adaptation in Response to High-Fat versus High-Carbohydrate Diets—Data from a Randomized Cross-Over Study in Healthy Subjects" Nutrients 13, no. 10: 3322. https://doi.org/10.3390/nu13103322
APA StyleWallenius, V., Elebring, E., Casselbrant, A., Laurenius, A., le Roux, C. W., Docherty, N. G., Biörserud, C., Björnfot, N., Engström, M., Marschall, H. -U., & Fändriks, L. (2021). Glycemic Control and Metabolic Adaptation in Response to High-Fat versus High-Carbohydrate Diets—Data from a Randomized Cross-Over Study in Healthy Subjects. Nutrients, 13(10), 3322. https://doi.org/10.3390/nu13103322