
Combination of Trans-Resveratrol and ε-Viniferin Induces a Hepatoprotective Effect in Rats with Severe Acute Liver Failure via Reduction of Oxidative Stress and MMP-9 Expression

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experimental Procedures
2.2. Spectrophotometric Analysis of Biochemical Parameters
2.3. Multiplex Analysis
2.4. Gene Expression Analyses
2.5. Alkaline Comet Assay
2.6. Statistical Analysis
3. Results
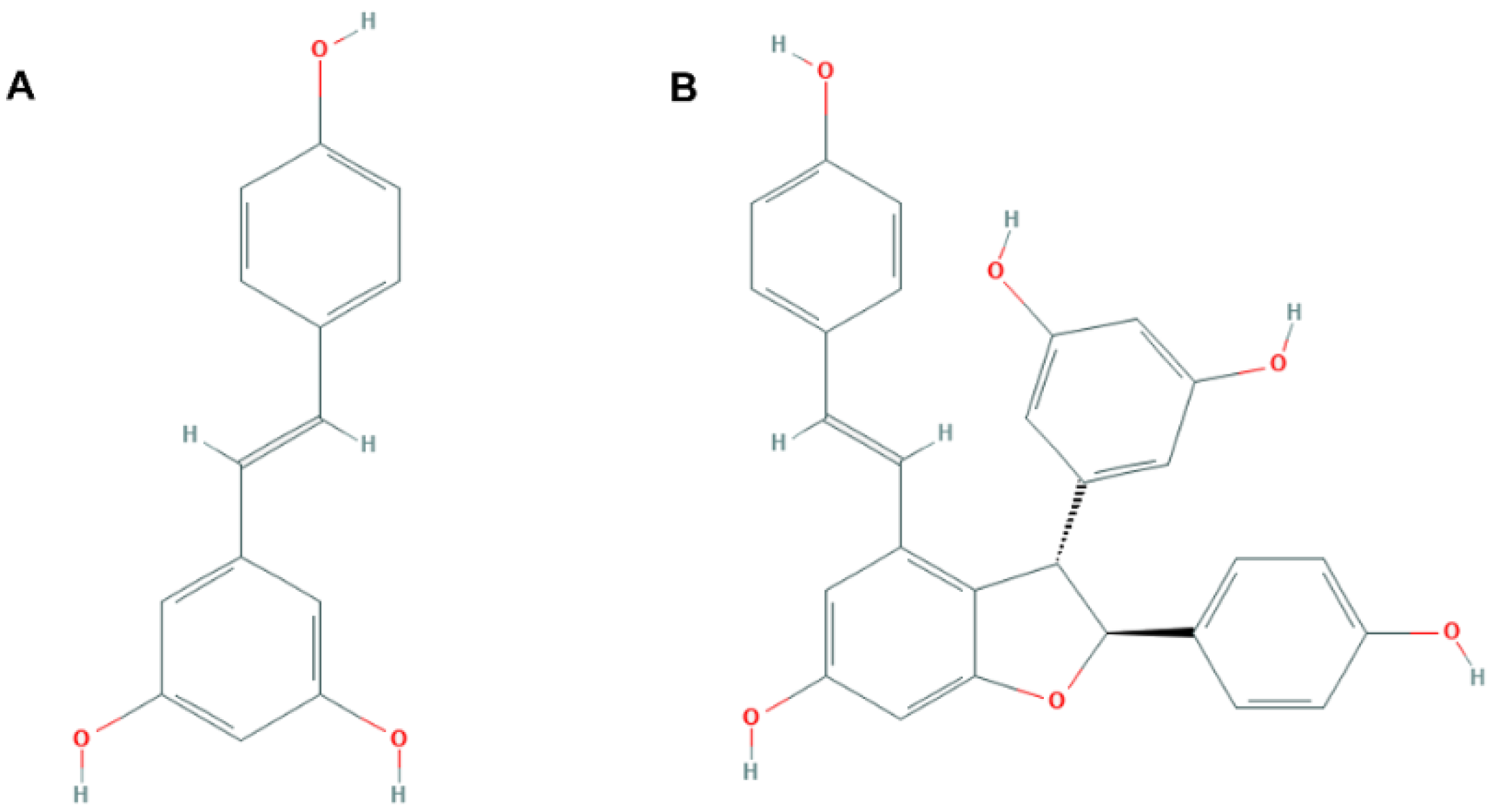
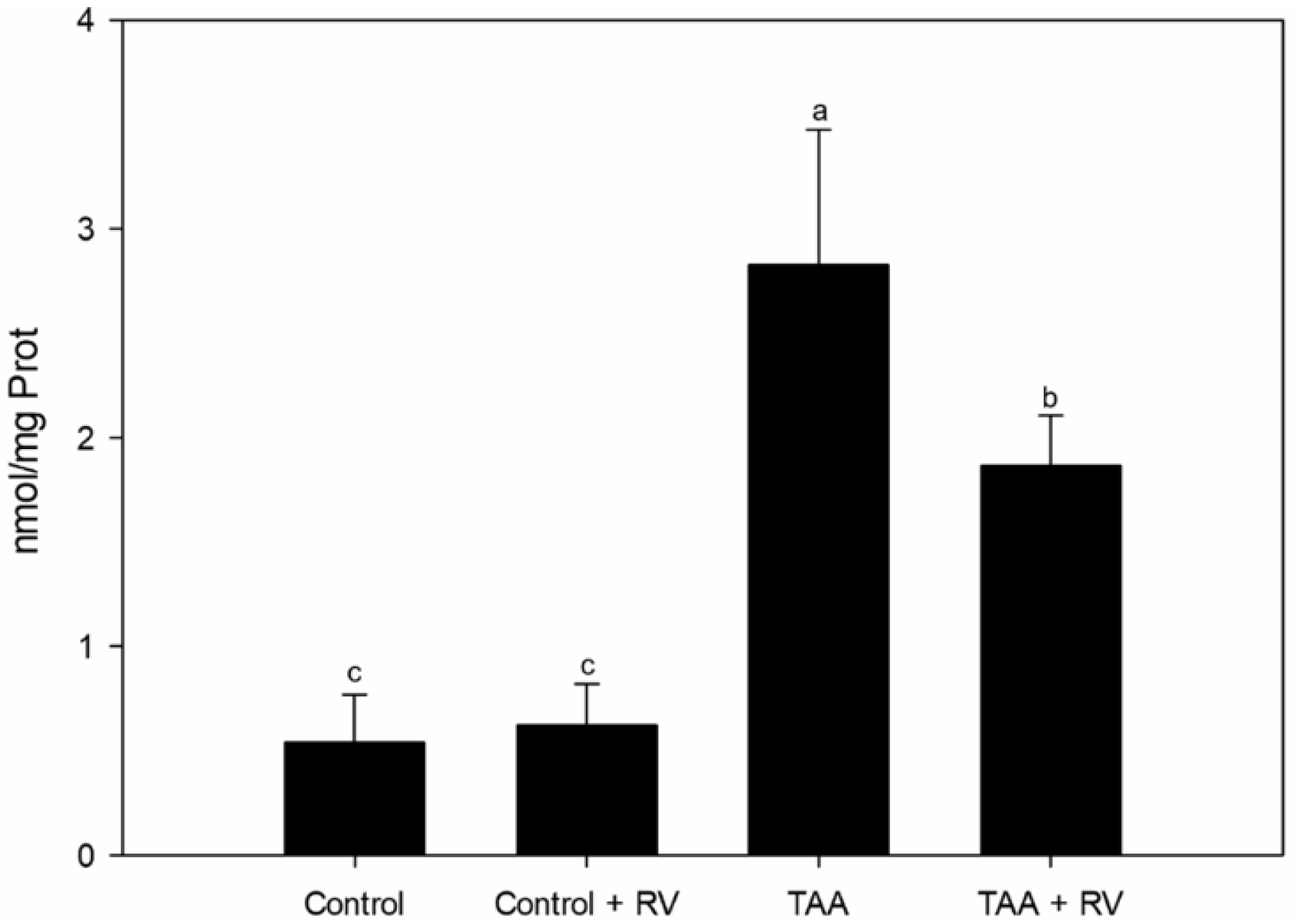
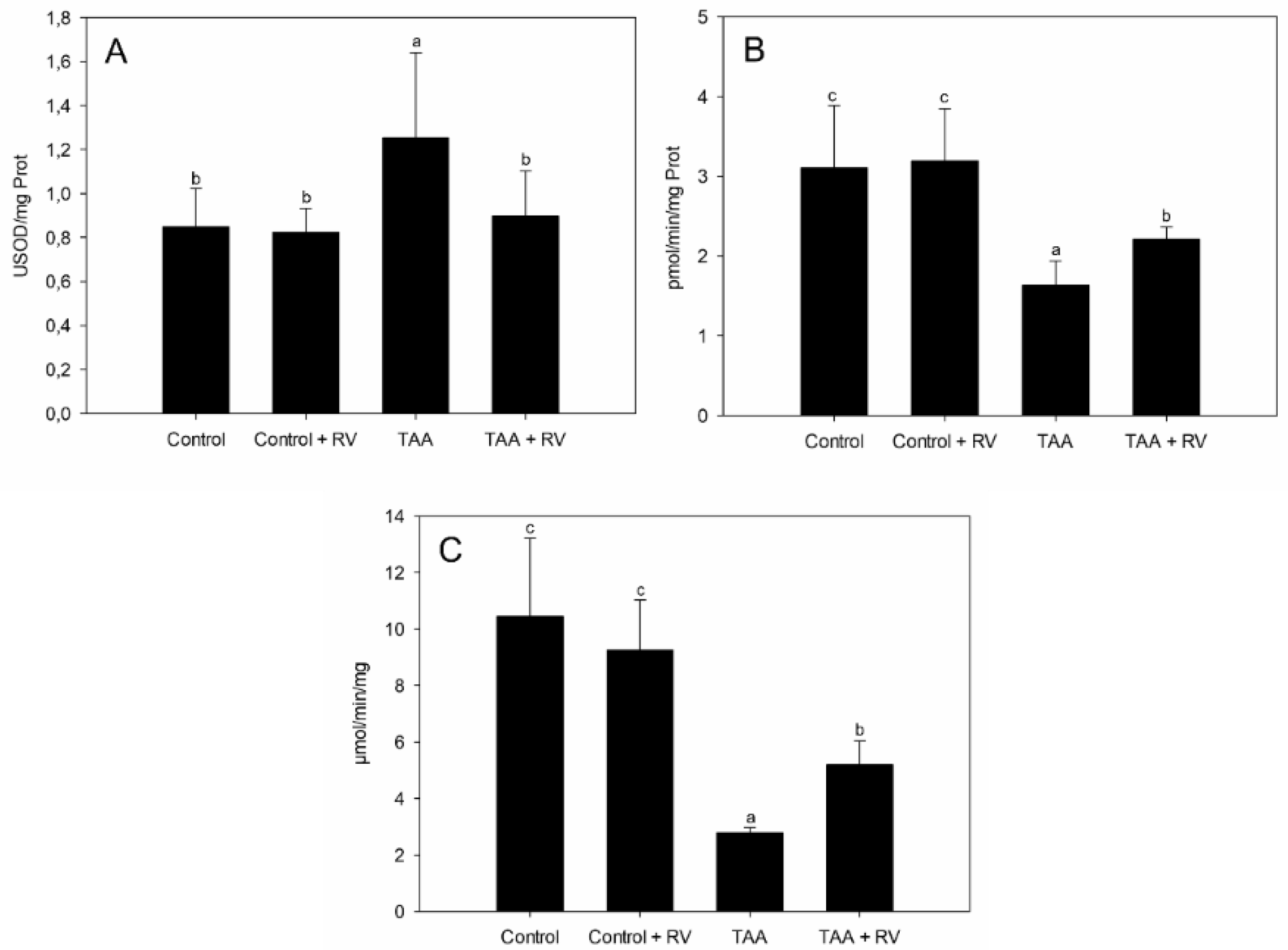
3.1. Assessment of Oxidative Stress
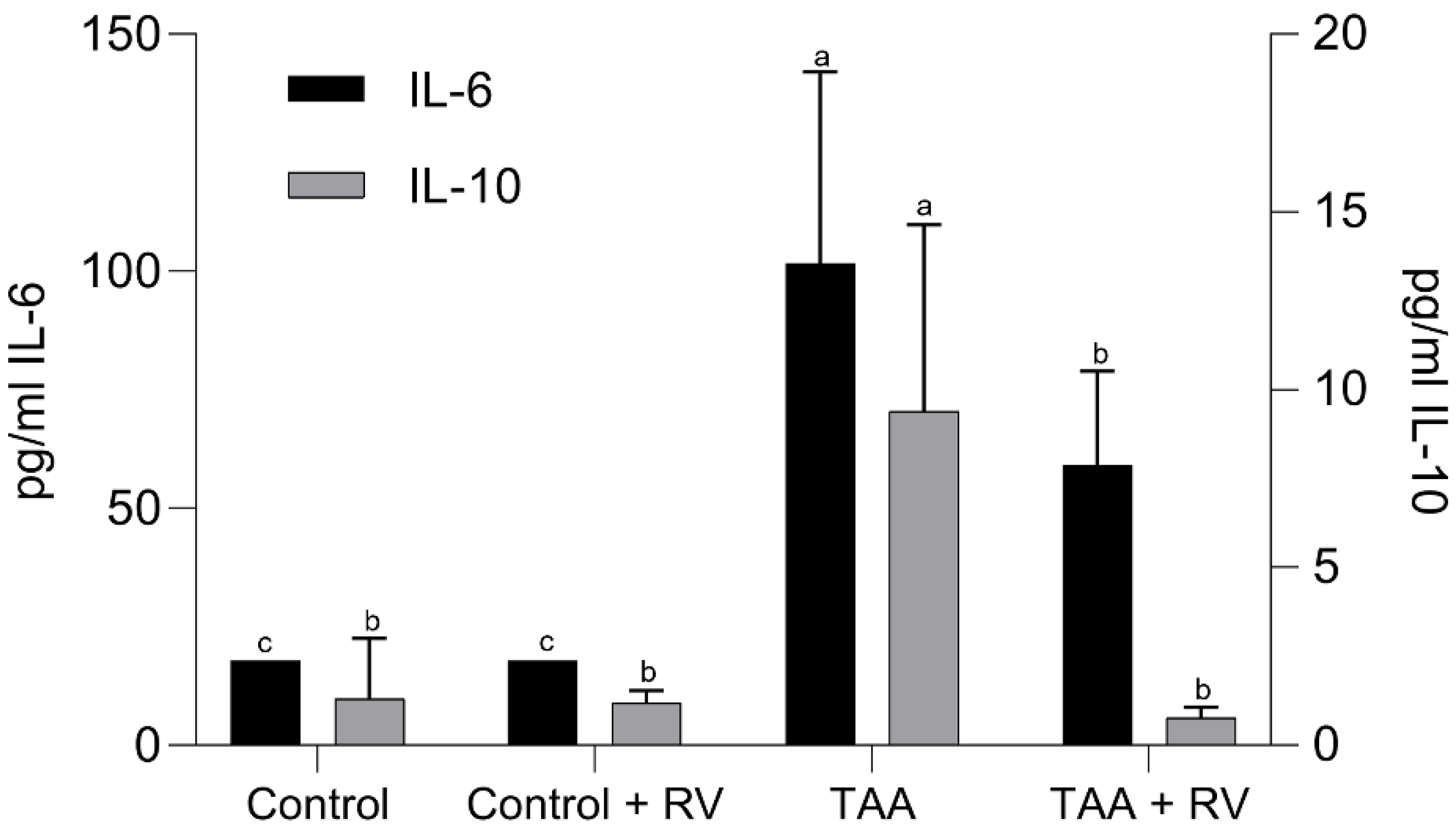
3.2. Assessment of the Inflammatory Process
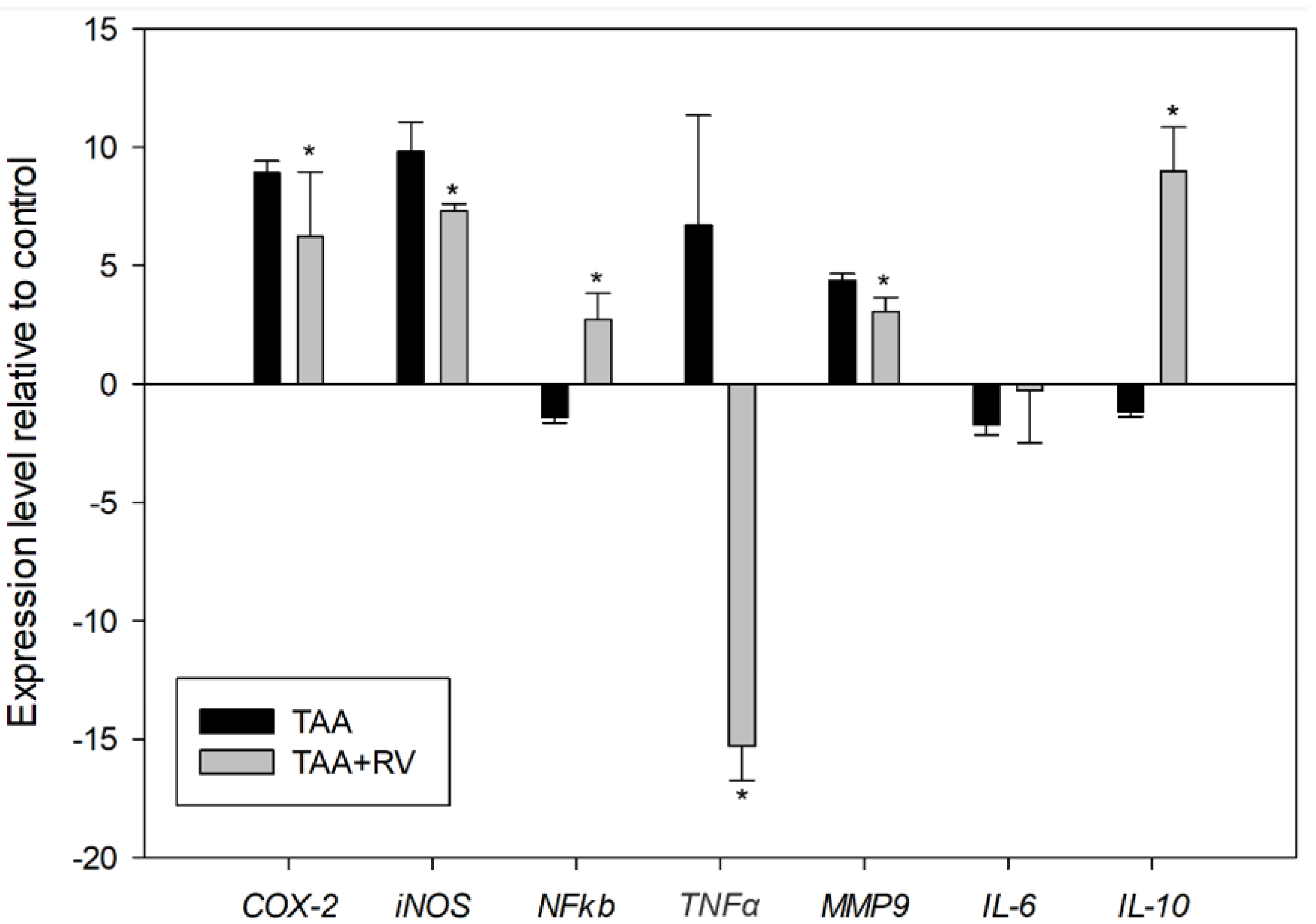
3.3. Assessment of mRNA Expression by Quantitative Real-Time PCR (RT-qPCR)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sachdeva, V.; Roy, A.; Bharadvaja, N. Current prospects of nutraceuticals: A review. Curr. Pharm. Biotechnol. 2020, 21, 884–896. [Google Scholar] [CrossRef]
- Kambiranda, D.M.; Basha, S.M.; Stringer, S.J.; Obuya, J.O.; Snowden, J.J. Multi-year Quantitative Evaluation of Stilbenoids Levels Among Selected Muscadine Grape Cultivars. Molecules 2019, 11, 981. [Google Scholar] [CrossRef] [Green Version]
- Sirerol, J.A.; Rodríguez, M.L.; Mena, S.; Asensi, M.A.; Estrela, J.M.; Ortega, A.L. Role of Natural Stilbenes in the Prevention of Cancer. Oxidative Med. Cell. Longev. 2016, 2016, 3128951. [Google Scholar] [CrossRef] [Green Version]
- Privat, C.; Telo, J.P.; Bernardes-Genisson, V.; Vieira, A.; Souchard, J.-P.; Nepveu, F. Antioxidant properties of trans-ε-Viniferin as compared to stilbene derivatives in aqueous and nonaqueous media. J. Agric. Food Chem. 2002, 50, 1213–1217. [Google Scholar] [CrossRef]
- Zghonda, N.; Yoshida, S.; Araki, M.; Kusunoki, M.; Mliki, A.; Ghorbel, A.; Miyazaki, H. Greater effectiveness of ε-viniferin in red wine than its monomer resveratrol for inhibiting vascular smooth muscle cell proliferation and migration. Biosci. Biotechnol. Biochem. 2011, 75, 1259–1267. [Google Scholar] [CrossRef] [Green Version]
- Stravitz, R.T.; Lee, W.M. Acute liver failure. Lancet 2019, 394, 869–881. [Google Scholar] [CrossRef]
- Schemitt, E.G.; Hartmann, R.M.; Colares, J.R.; Licks, F.; Salvi, J.O.; Marroni, C.A.; Marroni, N.P. Protective action of glutamine in rats with severe acute liver failure. World J. Hepatol. 2019, 11, 273–286. [Google Scholar] [CrossRef]
- Yin, X.; Gong, X.; Jiang, R.; Kuang, G.; Wang, B.; Zhang, L.; Xu, G.; Wan, J. Emodin ameliorated lipopolysaccharide-induced fulminant hepatic failure by blockade of TLR4/MD2 complex expression in Dgalactosamine-sensitized mice. Int. Immunopharmacol. 2014, 23, 66–72. [Google Scholar] [CrossRef]
- Hartmann, R.M.; Licks, F.; Schemitt, E.G.; Colares, J.R.; do Couto Soares, M.; Zabot, G.P.; Fillmann, H.S.; Marroni, N.P. Protective effect of glutamine on the main and adjacent organs damaged by ischemia-reperfusion in rats. Protoplasma 2017, 254, 2155–2168. [Google Scholar] [CrossRef]
- Xia, X.; Su, C.; Fu, J.; Zhang, P.; Jiang, X.; Xu, D.; Hu, L.; Song, E.; Song, Y. Role of α-lipoic acid in LPS/d-GalN induced fulminant hepatic failure in mice: Studies on oxidative stress, inflammation and apoptosis. Int. Immunopharmacol. 2014, 22, 293–302. [Google Scholar] [CrossRef]
- Naim, A.; Pan, Q.; Baig, M.S. Matrix Metalloproteinases (MMPs) in Liver Diseases. J. Clin. Exp. Hepatol. 2017, 7, 367–372. [Google Scholar] [CrossRef]
- Scheau, C.; Badarau, I.A.; Costache, R.; Caruntu, C.; Mihai, G.L.; Didilescu, A.C.; Constantin, C.; Neagu, M. The Role of Matrix Metalloproteinases in the Epithelial-Mesenchymal Transition of Hepatocellular Carcinoma. Anal. Cell. Pathol. 2019, 26, 9423907. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.J.; Wang, N.; Sun, Z.W.; Chen, J.; Cui, H.W. MiR-5692a promotes the invasion and metastasis of hepatocellular carcinoma via MMP9. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 4869–4878. [Google Scholar] [CrossRef]
- Roderfeld, M. Matrix metalloproteinase functions in hepatic injury and fibrosis. Matrix Biol. 2018, 68–69, 452–462. [Google Scholar] [CrossRef]
- Wu, C.; Zhu, Y.; Yu, M. Serum Metabonomics Analysis of Liver Failure Treated by Nonbioartificial Liver Support Systems. Can. J. Gastroenterol. Hepatol. 2018, 2018, 2586052. [Google Scholar] [CrossRef] [Green Version]
- Mattio, L.M.; Marengo, M.; Parravicini, C.; Eberini, I.; Dallavalle, S.; Bonomi, F.; Iametti, S.; Pinto, A. Inhibition of Pancreatic α-amylase by Resveratrol Derivatives: Biological Activity and Molecular Modelling Evidence for Cooperativity between Viniferin Enantiomers. Molecules 2019, 24, 3225. [Google Scholar] [CrossRef] [Green Version]
- Schemitt, E.G.; Colares, J.R.; Hartmann, R.M.; Morgan-Martins, M.; Marroni, C.A.; Tunon, M.J.; Marroni, N.P. Effect of glutamine on oxidative stress and infammation in a rat model of fulminant hepatic failure. Nutr. Hosp. 2016, 33, 92. [Google Scholar] [CrossRef]
- Brasilia, Law no. 11794, 8 of October of 2008. Available online: http://www.planalto.gov.br/ccivil_03/_ato2007-2010/2008/lei/l11794.htm (accessed on 18 September 2021).
- Brazil, Ministério da Ciência, Tecnologia e Inovação. Resolução normativa nº 30, de 2 de fevereiro de 2016. Diretriz brasileira para o cuidado e a utilização de animais em atividades de ensino ou de pesquisa científica—DBCA. Available online: http://www.icb.usp.br (accessed on 28 September 2019).
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzym. 1978, 52, 302–310. [Google Scholar]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]
- Boveris, A.; Chance, B. The mitochondrial generation of hydrogen peroxide. General properties and effect of hyperbaric oxygen. Biochem. J. 1973, 134, 707–716. [Google Scholar] [CrossRef]
- Flohé, L.; Günzler, W.A. Assays of glutathione peroxidase. Methods Enzymol. 1984, 105, 114–121. [Google Scholar]
- Tice, R.R.; Agurell, E.; Anderson, D.; Burlinson, B.; Hartmann, A.; Kobayashi, H.; Miyamae, Y.; Rojas, E.; Ryu, J.C.; Sasaki, Y.F. Single cell gel/comet assay: Guidelines for in vitro and in vivo genetic toxicology testing. Env. Mol. Mutagen. 2000, 35, 206–221. [Google Scholar] [CrossRef]
- Picada, J.N.; Flores, D.G.; Zettler, C.G.; Marroni, N.P.; Roesler, R.; Henriques, J.Á. DNA damage in brain cells of mice treated with an oxidized form of apomorphine. Mol. Brain Res. 2003, 114, 80–85. [Google Scholar] [CrossRef]
- Miguel, F.M.; Schemitt, E.G.; Colares, J.R.; Hartmann, R.M.; Morgan-Martins, M.I.; Marroni, N.P. Action of Vitamin E on Experimental Severe Acute Liver Failure. Arq. Gastroenterol. 2017, 54, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Vitaglione, P.; Ottanelli, B.; Milani, S.; Morisco, F.; Caporaso, N.; Fogliano, V. Dietary trans-resveratrol bioavailability and effect on CCl4-induced liver lipid peroxidation. J. Gastroenterol. Hepatol. 2009, 24, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Eissa, L.A.; Kenawy, H.I.; El-Karef, A.; Elsherbiny, N.M.; El-Mihi, K.A. Antioxidant and anti-inflammatory activities of berberine attenuate hepatic fibrosis induced by thioacetamide injection in rats. Chem.-Biol. Interact. 2018, 294, 91–100. [Google Scholar] [CrossRef]
- Urrutia-Hernández, T.A.; Santos-López, J.A.; Benedí, J.; Sánchez-Muniz, F.J.; Velázquez-González, C.; De la O.-Arciniega, M.; Jaramillo-Morales, O.A.; Bautista, M. Antioxidant and Hepatoprotective Effects of Croton hypoleucus Extract in an Induced-Necrosis Model in Rats. Molecules 2019, 24, 2533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukalingam, K.; Ganesan, K.; Xu, B. Protective Effect of Aqueous Extract from the Leaves of Justicia tranquebariesis against Thioacetamide-Induced Oxidative Stress and Hepatic Fibrosis in Rats. Antioxidants 2018, 7, 78. [Google Scholar] [CrossRef] [Green Version]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwa, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [Green Version]
- Pham, C.G.; Bubici, C.; Zazzeroni, F.; Papa, S.; Jones, J.; Alvarez, K.; Jayawardena, S.; De Smaele, E.; Cong, R.; Beaumont, C.; et al. Ferritin heavy chain upregulation by NF-kappaB inhibits TNFalpha-induced apoptosis by suppressing reactive oxygen species. Cell 2004, 119, 529–542. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, J.; Li, S.; He, B.; Mi, Y.; Cao, H.; Zhang, C.; Li, L. Ameliorative effect of grape seed proanthocyanidin extract on thioacetamide-induced mouse hepatic fibrosis. Toxicol. Lett. 2012, 213, 353–360. [Google Scholar] [CrossRef]
- Demirel, U.; Yalniz, M.; Aygün, C.; Orhan, C.; Tuzcu, M.; Sahin, K.; Ozercan, I.H.; Bahçecioğlu, I.H. Allopurinol ameliorates thioacetamide-induced acute liver failure by regulating cellular redox-sensitive transcription factors in rats. Inflammation 2012, 35, 1549–1557. [Google Scholar] [CrossRef]
- Yang, B.Y.; Zhang, X.Y.; Guan, S.W.; Hua, Z.C. Protective Effect of Procyanidin B2 against CCl4-Induced Acute Liver Injury in Mice. Molecules 2015, 20, 12250–12265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, B.; Qin, S.Y.; Hu, B.L.; Qin, Q.Y.; Jiang, H.X.; Luo, W. Resveratrol improves CCL4-induced liver fibrosis in mouse by upregulating endogenous IL-10 to reprogramme macrophages phenotype from M(LPS) to M(IL-4). Biomed. Pharmacother. 2019, 117, 109–110. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.M.; Taha, A.M.; Eldahshan, O.A.; Sayed, A.A.; Salem, A.M. Modulatory effect of Prosopis juliflora leaves on hepatic fibrogenic and fibrolytic alterations induced in rats by thioacetamide. Biomed. Pharmacother. 2019, 115, 108788. [Google Scholar] [CrossRef] [PubMed]
- Robert, S.; Gicquel, T.; Bodin, A.; Lagente, V.; Boichot, E. Characterization of the MMP/TIMP Imbalance and Collagen Production Induced by IL-1β or TNF-α Release from Human Hepatic Stellate Cells. PLoS ONE 2016, 11, e0153118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Pan, C.; Zhao, S.; Wang, Z.; Zhang, H.; Wu, W. Resveratrol inhibits tumor necrosis factor-alpha-mediated matrix metalloproteinase-9 expression and invasion of human hepatocellular carcinoma cells. Biomed. Pharmacother. 2008, 62, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Rašić, D.; Želježić, D.; Kopjar, N.; Kifer, D.; Klarić, M.Š.; Peraica, M. DNA damage in rat kidneys and liver upon subchronic exposure to single and combined ochratoxin A and citrinin. World Mycotoxin J. 2019, 12, 163–172. [Google Scholar] [CrossRef]
- Walle, T.; Hsieh, F.; DeLegge, M.H.; Oatis, J.E., Jr.; Walle, U.K. High absorption but very low bioavailability of oral resveratrol in humans. Drug Metab. Dispos. 2004, 32, 1377–1382. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Liao, X.; Xu, J.; Zhao, J.; Luo, J.; Kong, L. Development and validation of a sensitive and selective LC-MS/MS method for the determination of trans δ-veniferin, a resveratrol dehydrodimer, in rat plasma and its application to pharmacokinetics and bioavailability studies. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2014, 958, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Damage Index (0–400) | Damage Frequency (%) |
|---|---|---|
| CO | 69.57 ± 9.79 c | 38.57 ± 4.03 c |
| CO + RV | 81.57 ± 22.58 c | 42.14 ± 9.90 c |
| TAA | 297 ± 15.16 a | 91.75 ± 3.40 a |
| TAA + RV | 176 ± 32.04 b | 71.66 ± 13.57 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, J.C.; Schemitt, E.G.; Da Silva, J.; Marroni, N.P.; Lima, A.; Ferreira, R.B. Combination of Trans-Resveratrol and ε-Viniferin Induces a Hepatoprotective Effect in Rats with Severe Acute Liver Failure via Reduction of Oxidative Stress and MMP-9 Expression. Nutrients 2021, 13, 3677. https://doi.org/10.3390/nu13113677
Fernandes JC, Schemitt EG, Da Silva J, Marroni NP, Lima A, Ferreira RB. Combination of Trans-Resveratrol and ε-Viniferin Induces a Hepatoprotective Effect in Rats with Severe Acute Liver Failure via Reduction of Oxidative Stress and MMP-9 Expression. Nutrients. 2021; 13(11):3677. https://doi.org/10.3390/nu13113677
Chicago/Turabian StyleFernandes, João C., Elizângela G. Schemitt, Juliana Da Silva, Norma P. Marroni, Ana Lima, and Ricardo B. Ferreira. 2021. "Combination of Trans-Resveratrol and ε-Viniferin Induces a Hepatoprotective Effect in Rats with Severe Acute Liver Failure via Reduction of Oxidative Stress and MMP-9 Expression" Nutrients 13, no. 11: 3677. https://doi.org/10.3390/nu13113677
APA StyleFernandes, J. C., Schemitt, E. G., Da Silva, J., Marroni, N. P., Lima, A., & Ferreira, R. B. (2021). Combination of Trans-Resveratrol and ε-Viniferin Induces a Hepatoprotective Effect in Rats with Severe Acute Liver Failure via Reduction of Oxidative Stress and MMP-9 Expression. Nutrients, 13(11), 3677. https://doi.org/10.3390/nu13113677