
Ameliorating Effects of Coriander on Gastrocnemius Muscles Undergoing Precachexia in a Rat Model of Rheumatoid Arthritis: A Proteomics Analysis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiment
2.2. Measurement of Physiological Index and RA Biomarkers
2.3. Protein Preparation and Isobaric Tag for Relative and Absolute Quantitation (iTRAQ)-Based Quantitative Proteomics Analysis
2.4. Bioinformatics Analyses
2.5. Statistical Analyses
3. Results
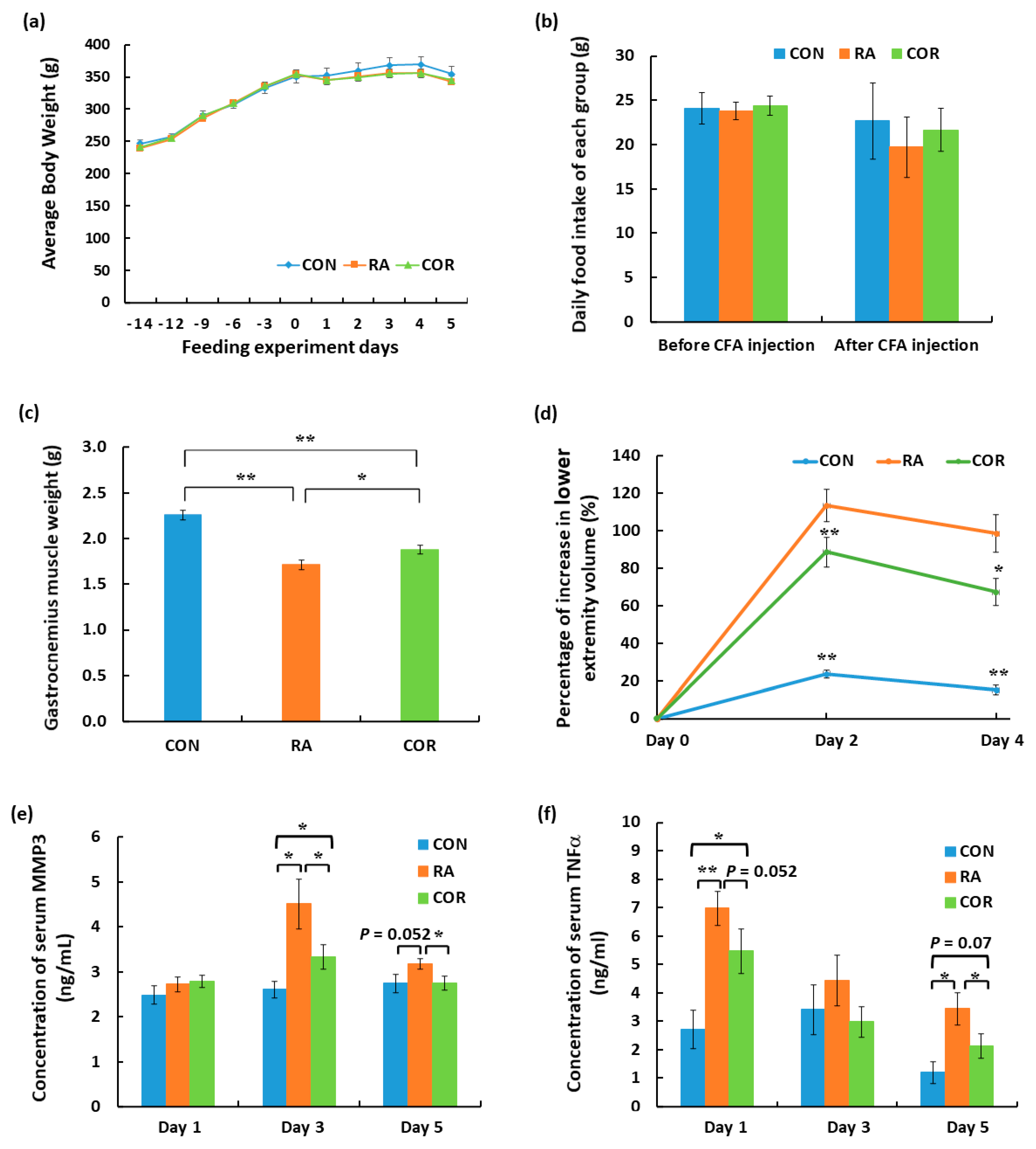
3.1. Assessment of the Physiological Index and Biomarkers of RA
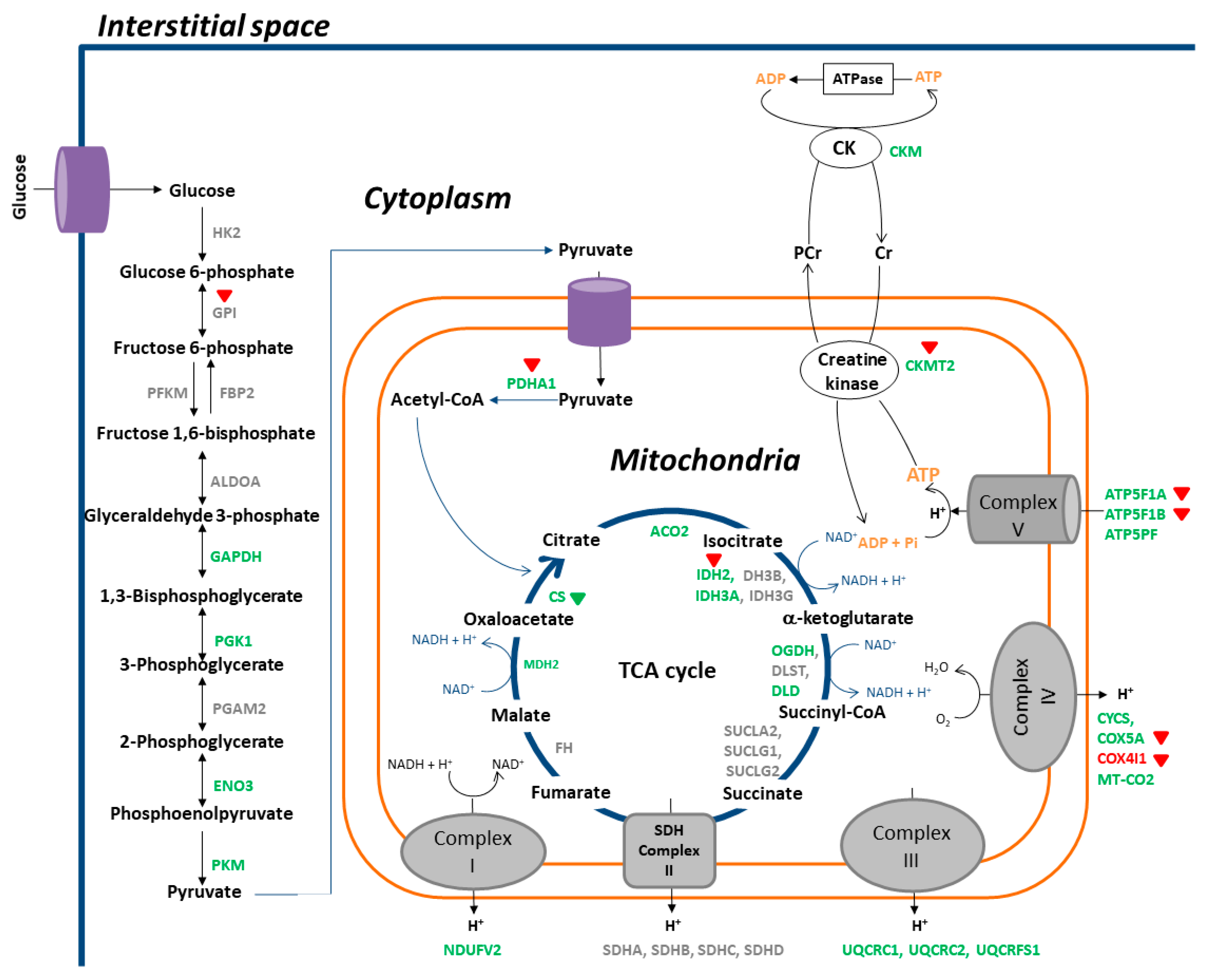
3.2. Identification of DEPs
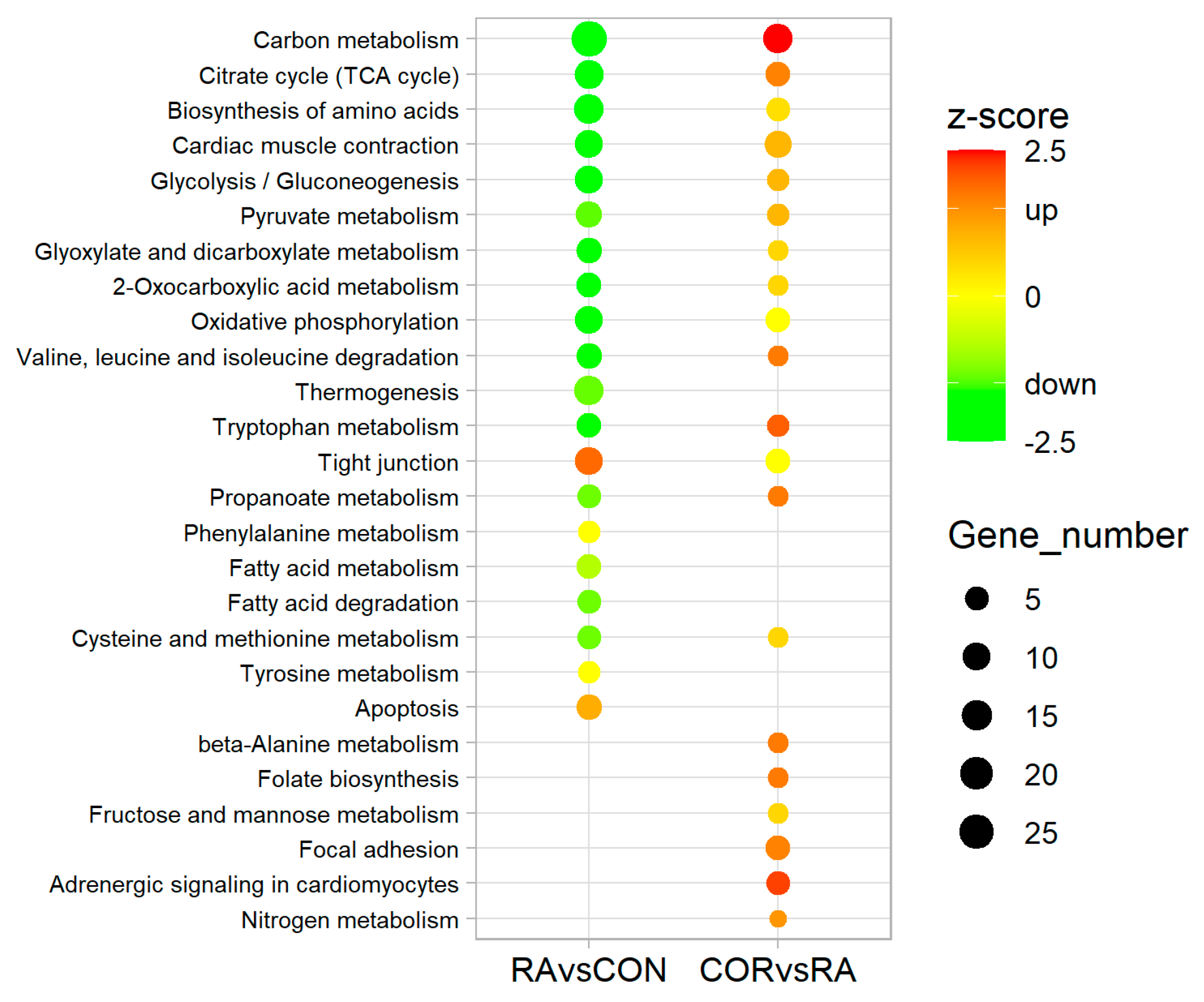
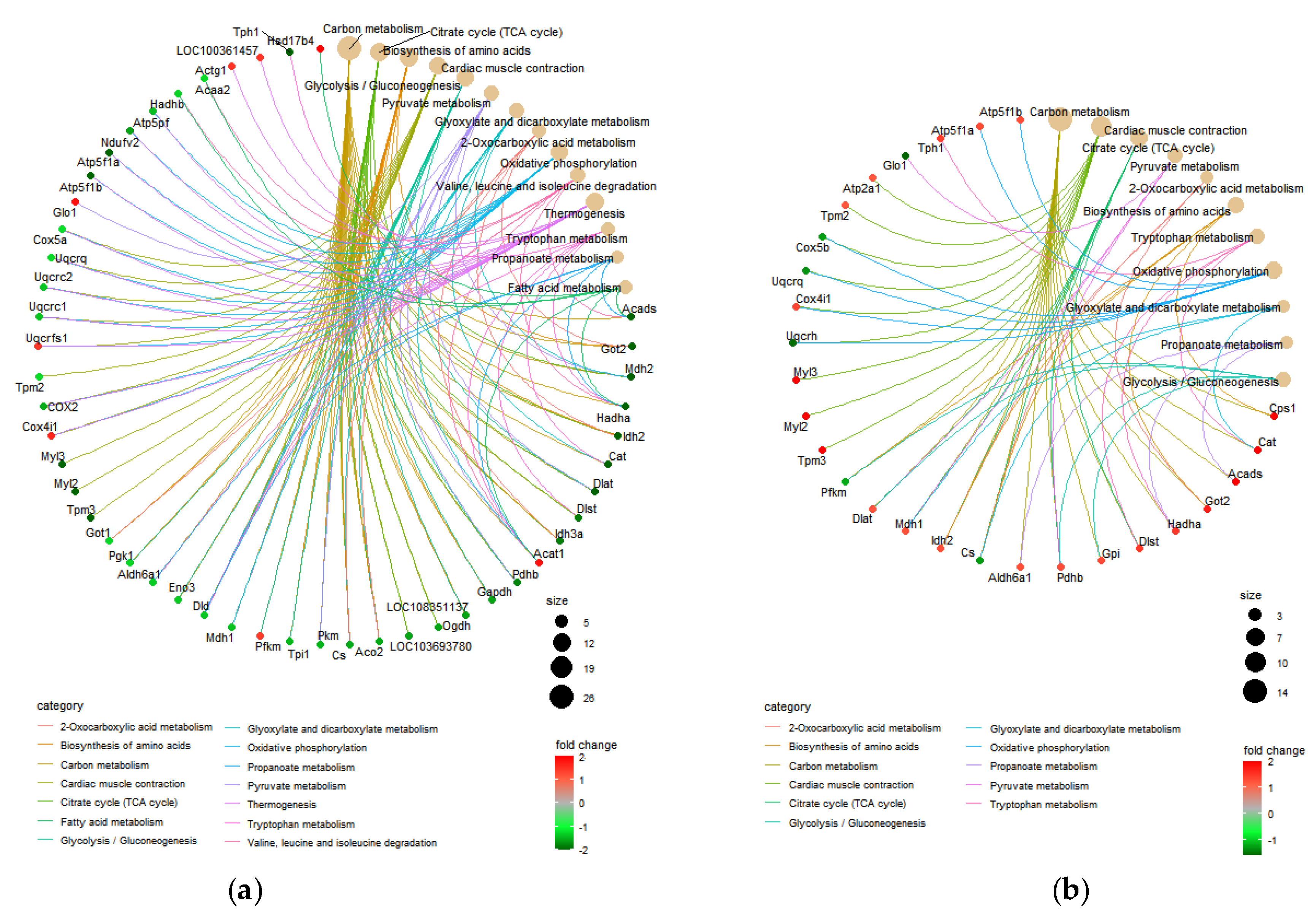
3.3. KEGG Pathway Analyses
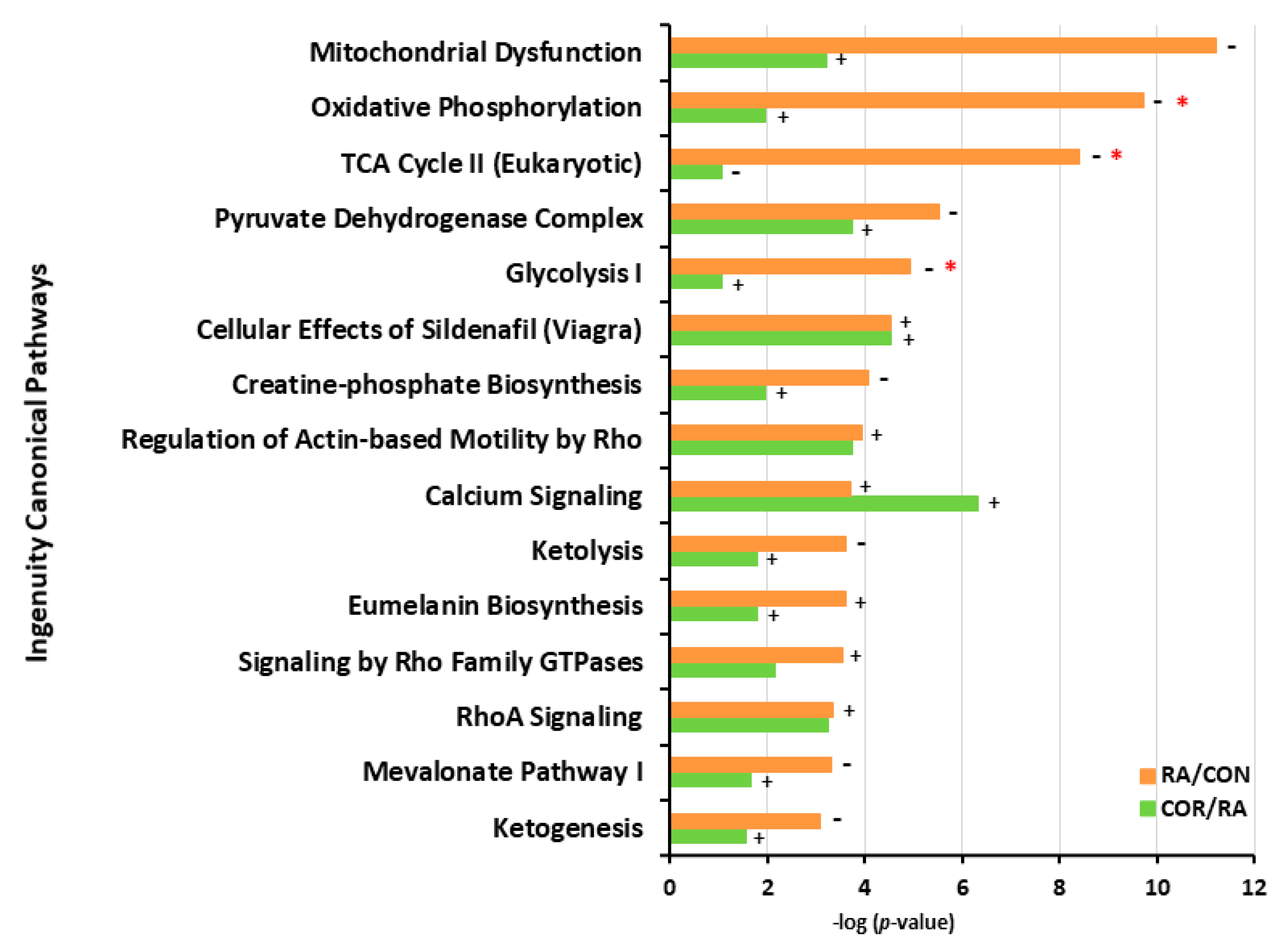
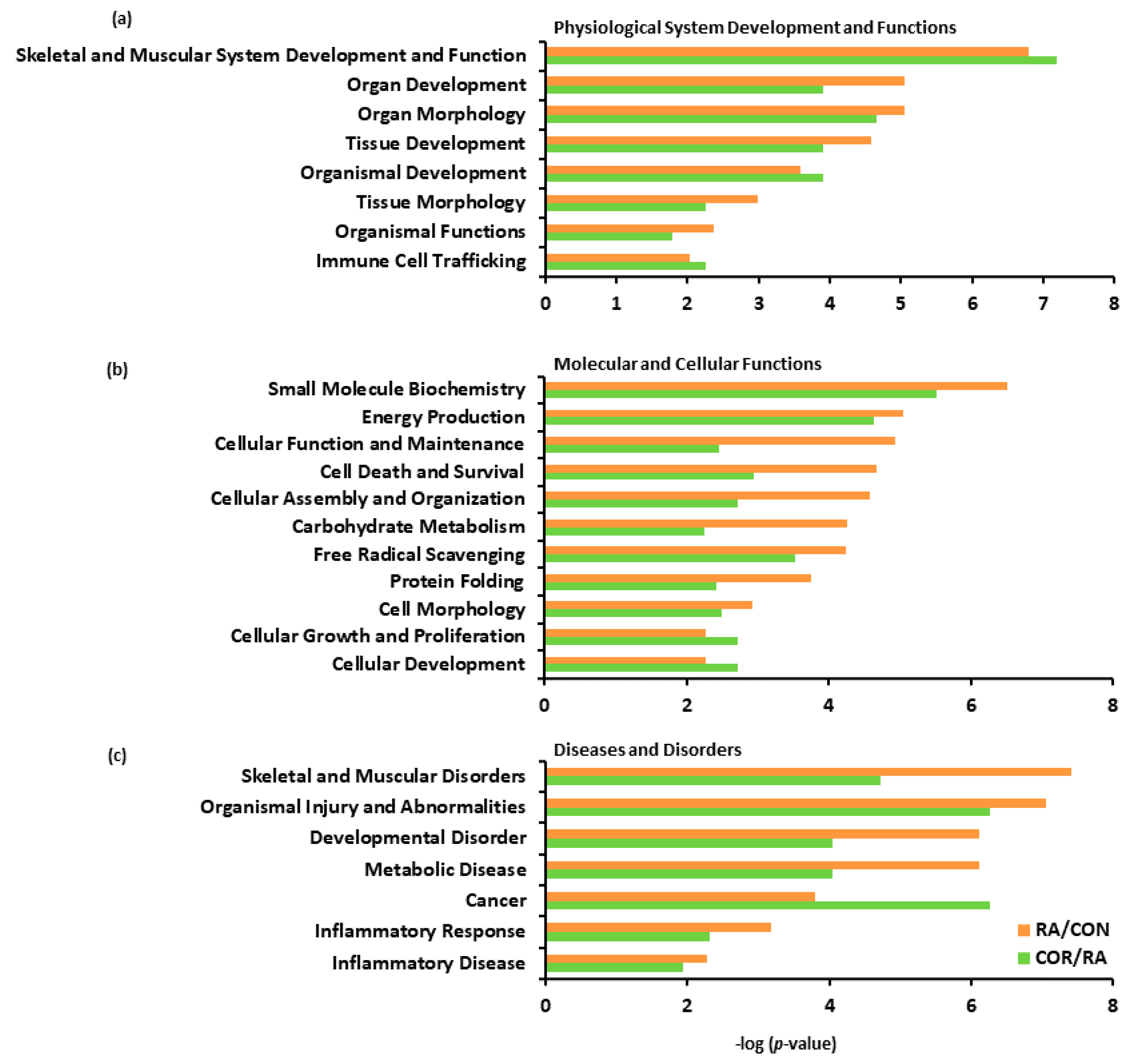
3.4. IPA
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Laribi, B.; Kouki, K.; M’Hamdi, M.; Bettaieb, T. Coriander (Coriandrum sativum L.) and its bioactive constituents. Fitoterapia 2015, 103, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.; Pandeya, S.; Yadav, S.K.; Singh, S.; Soni, P. A review on Coriandrum sativum (Linn.): An ayurvedic medicinal herb of happiness. J. Adv. Pharm. Healthc. Res. 2011, 1, 28–48. [Google Scholar]
- Khan, S.W.; Khatoon, S. Ethnobotanical studies on some useful herbs of Haramosh and Bugrote valleys in Gilgit, northern areas of Pakistan. Pak. J. Bot. 2008, 40, 43–58. [Google Scholar]
- Ugulu, I.; Baslar, S.; Yorek, N.; Dogan, Y. The investigation and quantitative ethnobotanical evaluation of medicinal plants used around Izmir province. Turk. J. Med. Plant Res. 2009, 3, 345–367. [Google Scholar]
- Walsmith, J.; Roubenoff, R. Cachexia in rheumatoid arthritis. Int. J. Cardiol. 2002, 85, 89–99. [Google Scholar] [CrossRef]
- Sugiyama, D.; Nishimura, K.; Tamaki, K.; Tsuji, G.; Nakazawa, T.; Morinobu, A.; Kumagai, S. Impact of smoking as a risk factor for developing rheumatoid arthritis: A meta-analysis of observational studies. Ann. Rheum. Dis. 2009, 69, 70–81. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.M.; Weinblatt, M.E. Rheumatoid arthritis. Lancet 2001, 358, 903–911. [Google Scholar] [CrossRef]
- Masuko, K. Rheumatoid Cachexia revisited: A metabolic co-morbidity in rheumatoid arthritis. Front. Nutr. 2014, 1, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Schueren, M.A.E.B.; Konijn, N.P.; Bultink, I.E.; Lems, W.F.; Earthman, C.P.; Van Tuyl, L.H. Relevance of the new pre-cachexia and cachexia definitions for patients with rheumatoid arthritis. Clin. Nutr. 2012, 31, 1008–1010. [Google Scholar] [CrossRef]
- Roubenoff, R.; Ward, L.M.; Holland, S.M.; Hellmann, D.B. Rheumatoid cachexia: Depletion of lean body mass in rheumatoid arthritis. Possible association with tumor necrosis factor. J. Rheumatol. 1992, 19, 1505–1510. [Google Scholar]
- Rajbhandary, R.; Khezri, A.; Panush, R.S. Rheumatoid Cachexia: What is it and why is it important? J. Rheumatol. 2011, 38, 406–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alabarse, P.V.; Lora, P.; Silva, J.M.; Santo, R.C.; Freitas, E.C.; de Oliveira, M.S.; Almeida, A.S.; Immig, M.; Teixeira, V.O.; Filippin, L.I.; et al. Collagen-induced arthritis as an animal model of rheumatoid cachexia. J. Cachex Sarcopenia Muscle 2018, 9, 603–612. [Google Scholar] [CrossRef]
- Muscaritoli, M.; Anker, S.; Argiles, J.M.; Aversa, Z.; Bauer, J.; Biolo, G.; Boirie, Y.; Bosaeus, I.; Cederholm, T.; Costelli, P.; et al. Consensus definition of sarcopenia, cachexia and pre-cachexia: Joint document elaborated by Special Interest Groups (SIG) “cachexia-anorexia in chronic wasting diseases” and “nutrition in geriatrics”. Clin. Nutr. 2010, 29, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Blauwhoff-Buskermolen, S.; de van der Schueren, M.A.; Verheul, H.M.; Langius, J.A. ‘Pre-cachexia’: A non-existing phenomenon in cancer? Ann. Oncol. 2014, 25, 1668–1669. [Google Scholar] [CrossRef] [PubMed]
- Fearon, K.; Strasser, F.; Anker, S.D.; Bosaeus, I.; Bruera, E.; Fainsinger, R.L.; Jatoi, A.; Loprinzi, C.; MacDonald, N.; Mantovani, G.; et al. Definition and classification of cancer cachexia: An international consensus. Lancet Oncol. 2011, 12, 489–495. [Google Scholar] [CrossRef]
- Kang, J.-S. Principles and applications of LC-MS/MS for the quantitative bioanalysis of analytes in various biological samples. In Tandem Mass Spectrometry—Applications and Principles; InTech: London, UK, 2012; pp. 459–510. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Pramanik, B.N. Application of LC/MS to proteomics studies: Current status and future prospects. Drug Discov. Today 2009, 14, 465–471. [Google Scholar] [CrossRef]
- van de Merbel, N.C. Advances in liquid chromatography–Tandem mass spectrometry (LC–MS–MS)-based quantitation of biopharmaceuticals in biological samples. LC-GC Europe 2015, 5, 38–42. [Google Scholar]
- Popa, C.; Netea, M.G.; van Riel, P.L.C.M.; van der Meer, J.W.M.; Stalenhoef, A.F.H. The role of TNF-α in chronic inflammatory conditions, intermediary metabolism, and cardiovascular risk. J. Lipid Res. 2007, 48, 751–762. [Google Scholar] [CrossRef] [Green Version]
- Uemura, Y.; Hayashi, H.; Takahashi, T.; Saitho, T.; Umeda, R.; Ichise, Y.; Sendo, S.; Tsuji, G.; Kumagai, S. MMP-3 as a biomarker of disease activity of rheumatoid arthritis. Rinsho Byori. Jpn. J. Clin. Pathol. 2015, 63, 1357–1364. [Google Scholar]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS A J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Walter, W.; Sánchez-Cabo, F.; Ricote, M. GOplot: An R package for visually combining expression data with functional analysis. Bioinformatics 2015, 31, 2912–2914. [Google Scholar] [CrossRef]
- Krämer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in ingenuity pathway analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]
- Casetti, F.; Bartelke, S.; Biehler, K.; Augustin, M.; Schempp, C.M.; Frank, U. Antimicrobial activity against bacteria with dermatological relevance and skin tolerance of the essential oil from Coriandrum sativum L. fruits. Phytother. Res. 2011, 26, 420–424. [Google Scholar] [CrossRef]
- Thompson, A.; Meah, D.; Ahmed, N.; Conniff-Jenkins, R.; Chileshe, E.; Phillips, C.O.; Claypole, T.C.; Forman, D.W.; Row, P.E. Comparison of the antibacterial activity of essential oils and extracts of medicinal and culinary herbs to investigate potential new treatments for irritable bowel syndrome. BMC Complement. Altern. Med. 2013, 13, 338. [Google Scholar] [CrossRef] [Green Version]
- Pawar, V.A.; Bhagat, T.B.; Toshniwal, M.R.; Mokashi, N.D.; Khandelwal, K.R. Formulation and evaluation of dental gel containing essential oil of Coriander against oral pathogens. Int. Res. J. Pharm. 2013, 4, 48–54. [Google Scholar] [CrossRef]
- Misharina, T.A.; Samusenko, A.L. Antioxidant properties of essential oils from lemon, grapefruit, coriander, clove, and their mixtures. Appl. Biochem. Microbiol. 2008, 44, 438–442. [Google Scholar] [CrossRef]
- Deepa, B.; Anuradha, C.V. Antioxidant potential of Coriandrum sativum L. seed extract. Indian J. Exp. Boil. 2011, 49, 30–38. [Google Scholar]
- Al-Juhaimi, F.; Ghafoor, K. Total phenols and antioxidant activities of leaf and stem extracts from coriander, mint and parsley grown in Saudi Arabia. Pak. J. Bot. 2011, 43, 2235–2237. [Google Scholar]
- Reuter, J.; Huyke, C.; Casetti, F.; Theek, C.; Frank, U.; Augustin, M.; Schempp, C. Anti-inflammatory potential of a lipolotion containing coriander oil in the ultraviolet erythema test. J. Dtsch. Dermatol. Ges. 2008, 6, 847–851. [Google Scholar] [CrossRef] [PubMed]
- Sonika, G.; Manubala, R.; Deepak, J. Comparative studies on anti-inflammatory activity of Coriandrum Sativum, Datura stramonium and Azadirachta Indica. Asian J. Exp. Biol. Sci. 2010, 1, 151–154. [Google Scholar]
- Gomez-Flores, R.; Hernández-Martínez, H.; Tamez-Guerra, P.; Tamez-Guerra, R.; Quintanilla-Licea, R.; Monreal-Cuevas, E.; Rodríguez-Padilla, C. Antitumor and immunomodulating potential of Coriandrum sativum. J. Nat. Prod. 2010, 3, 54–63. [Google Scholar]
- Tang, E.L.H.; Rajarajeswaran, J.; Fung, S.Y.; Kanthimathi, M.S. Antioxidant activity of Coriandrum sativum and protection against DNA damage and cancer cell migration. BMC Complement. Altern. Med. 2013, 13, 347. [Google Scholar] [CrossRef] [Green Version]
- Chithra, V.; Leelamma, S. Coriandrum sativum—Effect on lipid metabolism in 1,2-dimethyl hydrazine induced colon cancer. J. Ethnopharmacol. 2000, 71, 457–463. [Google Scholar] [CrossRef]
- Emam, G.M.; Heydari, H.G. Anticonvulsant effect of extract and essential oil of Coriandrum sativum seed in concious mice. Iran. J. Pharm. Res. 2004, 3, 71. [Google Scholar]
- Pathan, A.; Kothawade, K.; Logade, M.N. Anxiolytic and analgesic effect of seeds of Coriandrum sativum Linn. Int. J. Res. Pharm. Chem. 2011, 1, 1087–1099. [Google Scholar]
- Waheed, A.; Miana, G.; Ahmad, S.; Khan, M.A. Clinical investigation of hypoglycemic effect of Coriandrum sativum in type-2 (NIDDM) diabetic patients. Pak. J. Pharm. Sci. 2006, 23, 7–11. [Google Scholar]
- Naquvi, K.J.; Ali, M.; Ahmad, J. Antidiabetic activity of aqueous extract of Coriandrum sativum L. fruits in streptozotocin induced rats. Indian J. Exp. Biol. 2004, 42, 909–912. [Google Scholar]
- Pandeya, K.B.; Tripathi, I.P.; Mishra, M.K.; Dwivedi, N.; Pardhi, Y.; Kamal, A.; Gupta, P.; Dwivedi, N.; Mishra, C. A critical review on traditional herbal drugs: An emerging alternative drug for diabetes. Int. J. Org. Chem. 2013, 3, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Cioanca, O.; Hritcu, L.; Mihasan, M.; Hancianu, M. Cognitive-enhancing and antioxidant activities of inhaled coriander volatile oil in amyloid β (1–42) rat model of Alzheimer’s disease. Physiol. Behav. 2013, 120, 193–202. [Google Scholar] [CrossRef]
- Jang, Y.C.; Lustgarten, M.S.; Liu, Y.; Muller, F.L.; Bhattacharya, A.; Liang, H.; Salmon, A.B.; Brooks, S.V.; Larkin, L.; Hayworth, C.R.; et al. Increased superoxide in vivo accelerates age-associated muscle atrophy through mitochondrial dysfunction and neuromuscular junction degeneration. FASEB J. 2010, 24, 1376–1390. [Google Scholar] [CrossRef] [Green Version]
- Dupuis, L.; De Aguilar, J.-L.G.; Echaniz-Laguna, A.; Eschbach, J.; Rene, F.; Oudart, H.; Halter, B.; Huze, C.; Schaeffer, L.; Bouillaud, F.; et al. Muscle mitochondrial uncoupling dismantles neuromuscular junction and triggers distal degeneration of motor neurons. PLoS ONE 2009, 4, e5390. [Google Scholar] [CrossRef] [Green Version]
- Rygiel, K.A.; Picard, M.; Turnbull, D. The ageing neuromuscular system and sarcopenia: A mitochondrial perspective. J. Physiol. 2016, 594, 4499–4512. [Google Scholar] [CrossRef]
- Shum, A.M.Y.; Poljak, A.; Bentley, N.L.; Turner, N.; Tan, T.C.; Polly, P. Proteomic profiling of skeletal and cardiac muscle in cancer cachexia: Alterations in sarcomeric and mitochondrial protein expression. Oncotarget 2018, 9, 22001–22022. [Google Scholar] [CrossRef] [Green Version]
- Constantinou, C.; De Oliveira, C.C.F.; Mintzopoulos, D.; Busquets, S.; He, J.; Kesarwani, M.; Mindrinos, M.; Rahme, L.G.; Argiles, J.M.; Tzika, A.A. Nuclear magnetic resonance in conjunction with functional genomics suggests mitochondrial dysfunction in a murine model of cancer cachexia. Int. J. Mol. Med. 2010, 27, 15–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallimann, T.; Wyss, M.; Brdiczka, D.; Nicolay, K.; Eppenberger, H.M. Intracellular compartmentation, structure and function of creatine kinase isoenzymes in tissues with high and fluctuating energy demands: The ‘phosphocreatine circuit’ for cellular energy homeostasis. Biochem. J. 1992, 281, 21–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talmadge, R.J. Myosin heavy chain isoform expression following reduced neuromuscular activity: Potential regulatory mechanisms. Muscle Nerve 2000, 23, 661–679. [Google Scholar] [CrossRef]
- Bigard, A.-X.; Boehm, E.; Veksler, V.; Mateo, P.; Anflous, K.; Ventura-Clapier, R. Muscle unloading induces slow to fast transitions in myofibrillar but not mitochondrial properties. Relevance to skeletal muscle abnormalities in heart failure. J. Mol. Cell. Cardiol. 1998, 30, 2391–2401. [Google Scholar] [CrossRef]
- Ohira, Y.; Yoshinaga, T.; Nomura, T.; Kawano, F.; Ishihara, A.; Nonaka, I.; Roy, R.; Edgerton, V. Gravitational unloading effects on muscle fiber size, phenotype and myonuclear number. Adv. Space Res. 2002, 30, 777–781. [Google Scholar] [CrossRef]
- Canepari, M.; Cappelli, V.; Pellegrino, M.; Zanardi, M.; Reggiani, C. Thyroid hormone regulation of MHC Isoform composition and Myofibrillar ATPase activity in rat skeletal muscles. Arch. Physiol. Biochem. 1998, 106, 308–315. [Google Scholar] [CrossRef]
- Oberbach, A.; Bossenz, Y.; Lehmann, S.; Niebauer, J.; Adams, V.; Paschke, R.; Schön, M.R.; Blüher, M.; Punkt, K. Altered fiber distribution and fiber-specific Glycolytic and Oxidative enzyme activity in skeletal muscle of patients with type 2 diabetes. Diabetes Care 2006, 29, 895–900. [Google Scholar] [CrossRef] [Green Version]
- Penna, F.; Ballarò, R.; Beltrá, M.; De Lucia, S.; Costelli, P. Modulating metabolism to improve cancer-induced muscle wasting. Oxidative Med. Cell. Longev. 2018, 2018, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pin, F.; Busquets, S.; Toledo, M.; Camperi, A.; Lopez-Soriano, F.J.; Costelli, P.; Argiles, J.M.; Penna, F. Combination of exercise training and erythropoietin prevents cancer-induced muscle alterations. Oncotarget 2015, 6, 43202–43215. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Pessin, J.E. Mechanisms for fiber-type specificity of skeletal muscle atrophy. Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 243–250. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UniProt ID | Protein Name, Symbol (from IPA) | FC (RA/CON) | FC (COR/RA) |
|---|---|---|---|
| Characteristic proteins of fast-twitch fibers | |||
| P02625 | Parvalbumin alpha (PVALB) | 1.55 | −1.35 b |
| Q29RW1 | Myosin heavy chain 4 (MYH4) | 1.98 b | −1.15 b |
| P04466 | Myosin regulatory light chain 2 (fast skeletal muscle isoform) (MYLPF) | 1.24 | −1.22 b |
| P27768 | Troponin I (fast skeletal muscle) (TNNI2) | 1.21 | 1.01 |
| Characteristic proteins of slow-twitch fibers | |||
| Q9QZ76 | Myoglobin (MB) | −1.75 (0.07) | 1.27 |
| P08733 | Myosin regulatory light chain 2 (slow skeletal/cardiac muscle isoform) (MYL2) | -10 a | 4.32 (0.09) |
| P16409 | Myosin light chain 3 (slow-twitch muscle) (MYL3) | −8.33 b | 4.25 a |
| P58775 | Tropomyosin beta chain (slow skeletal muscle isoform) (TPM2) | −1.22 (0.08) | 1.21 |
| P13413 | Troponin I (slow skeletal muscle) (TNNI1) | -5 a | 3.11 (0.06) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, H.; Wen, Y.; Aw, W.; Saito, K.; Kato, H. Ameliorating Effects of Coriander on Gastrocnemius Muscles Undergoing Precachexia in a Rat Model of Rheumatoid Arthritis: A Proteomics Analysis. Nutrients 2021, 13, 4041. https://doi.org/10.3390/nu13114041
Jia H, Wen Y, Aw W, Saito K, Kato H. Ameliorating Effects of Coriander on Gastrocnemius Muscles Undergoing Precachexia in a Rat Model of Rheumatoid Arthritis: A Proteomics Analysis. Nutrients. 2021; 13(11):4041. https://doi.org/10.3390/nu13114041
Chicago/Turabian StyleJia, Huijuan, Ya Wen, Wanping Aw, Kenji Saito, and Hisanori Kato. 2021. "Ameliorating Effects of Coriander on Gastrocnemius Muscles Undergoing Precachexia in a Rat Model of Rheumatoid Arthritis: A Proteomics Analysis" Nutrients 13, no. 11: 4041. https://doi.org/10.3390/nu13114041
APA StyleJia, H., Wen, Y., Aw, W., Saito, K., & Kato, H. (2021). Ameliorating Effects of Coriander on Gastrocnemius Muscles Undergoing Precachexia in a Rat Model of Rheumatoid Arthritis: A Proteomics Analysis. Nutrients, 13(11), 4041. https://doi.org/10.3390/nu13114041