1. Introduction
Green tea (GT) polyphenols have been widely investigated for their potential therapeutic health benefits from an antioxidant [
1], anti-inflammatory [
2,
3], chemoprotective [
4] and cardio-metabolic perspective [
5,
6,
7]. The catechin content of GT, of which (−)-epigallocatechin−3-gallate (EGCG) may account for up to 80% [
8], may be primarily related to adaptive mechanisms pertinent to health benefits. GT catechins may have specific thermogenic effects through the inhibition of catechol-o-methyl transferase (COMT) and subsequent catecholamine, cyclic adenosine monophosphate (cAMP) and lipolytic activity [
9]. Consequently, these effects may enhance whole-body fat oxidation [
10,
11], and GT catechins may have important metabolic effects pertinent to reduced adipose tissue [
12], improved body mass index (BMI) and/or body composition [
10], and lowered circulating metabolites (e.g., low-density lipoprotein cholesterol (LDL-c) [
13]). However, such mechanisms have also been challenged based on limited evidence from studies [
14]. Regular consumption of GT catechins has also been proposed to exert ‘calorie-restriction-mimetic’ effects [
15] over time, through modulated cell signalling (e.g., peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC1-α), sirtuin 1 (SIRT1) and AMP-activated protein kinase (AMPK) pathways), influencing gene expression associated with mitochondrial efficiency and fat oxidation [
14], and is supported through emerging experimental evidence [
16].
In terms of fat oxidation, studies with administration of GT extract (GTE) for <7 days, providing between 270 and 375 mg·d
−1 EGCG, have reported significant improvements in whole-body fat oxidation rates in untrained and overweight individuals both at rest [
17,
18], during exercise [
18] and post-exercise [
19], as well as increased 24 hour energy expenditure [
11]. Elevated catecholamine and glycerol concentrations in response to acute EGCG supplementation [
11,
19] support mechanisms associated with COMT inhibition. However, this may, in part, be explained through the inclusion of high-intensity exercise invoking elevated catecholamine levels [
19]. Surprisingly, the benefits observed in these studies occurred despite GTE consumption with food (which has been reported to reduce GTE bioavailability [
20]), inferring that observed improvements in fat oxidation likely occurred in response to pre-test fasted intake of GTE. Elsewhere, studies have reported no effect of short-term GTE supplementation on fat oxidation in trained individuals, with doses between 270 and 506 mg·d
−1 EGCG [
21,
22], suggesting that GTE may be beneficial only for untrained/overweight individuals.
In the longer term (>3 weeks), the effects of GTE supplementation on fat oxidation are equivocal. Low-dose GTE supplementation (68 mg·d
−1 EGCG) for 3 weeks did not improve fat oxidation during exercise in trained individuals [
23,
24], although improvements in cardio-metabolic and inflammatory metabolites (e.g., increased high-density lipoprotein cholesterol (HDL-c) [
23], and reduced C-reactive protein [
24]) were reported. A four-week supplementation period of moderate-dose decaffeinated GTE (dGTE; 400 mg·d
−1 EGCG) significantly improved total fat oxidation rates during exercise, and reduced body fat percentage in one study [
25], but not elsewhere, even when employing higher-dose dGTE (624 mg·d
−1 EGCG [
26]), with both studies involving healthy, lean males.
With regards to cardio-metabolic risk factors, studies lasting 8-12 weeks also provide equivocal findings. Low-dose GTE supplementation (99 mg·d
−1 GTE) for 8 weeks was not effective in improving body fat percentage, BMI, blood pressure, triglyceride and LDL-c levels over that obtained through introduction of aerobic training in overweight/obese, previously sedentary females [
27]. Likewise, high-dose caffeinated GTE (1206.9 mg·d
−1 GTE; 595.8 mg·d
−1 EGCG) for 12 weeks did not improve resting energy expenditure, body mass, BMI, waist to hip ratio (WHR) or fat mass over that explained through sustained inclusion of a low-energy diet in overweight woman [
28]. In contrast, an eight-week GTE supplementation period (500 mg·d
−1 GTE; ~225 mg·d
−1 EGCG) significantly improved body mass, BMI, body fat percentage, visceral fat area (VFA) and adiponectin levels above that obtained through introduction of aerobic training in previously sedentary overweight women [
29] and men [
30]. Therefore, it is unclear whether GTE supplementation positively impacts on fat oxidation, body composition and cardio-metabolic risk factors in overweight individuals, particularly when unaccustomed diet or exercise interventions are introduced.
In addition to the range of EGCG doses used, confounding factors such as GTE type and bioavailability may, in part, explain the varying findings observed. Only a handful of studies have employed a dGTE formula [
17,
18,
25,
26], whereas positive benefits found elsewhere could be explained through the thermogenic effects of caffeine [
11,
19,
29,
30]. Whilst positive findings have been observed with a moderate EGCG dose (>300 mg·d
−1), improving the bioavailability and effectiveness of GTE through standardised consumption away from food [
20] and/or inclusion of antioxidant nutrients (e.g., quercetin, α-lipoic acid (LA), curcumin [
31,
32,
33,
34]) may lead to enhanced fat oxidation, body composition, and/or cardio-metabolic risk factor improvement, particularly in overweight individuals. Indeed, precedent for the approach of combining GTE with quercetin or LA has previously been observed in animal models [
35,
36]. Elsewhere, use of a GT beverage with antioxidant supplementation has been shown to enhance exercise capacity, glucose tolerance and body composition in older individuals [
37]. However, there is a paucity of research confirming potential benefits of GTE compounds with human participants, despite novel combinations already being commercially available. Therefore, it is pertinent to investigate the efficacy such compounds compared with GTE only use.
Therefore, this study aimed to assess the longer-term impact of moderate dGTE supplementation (with or without antioxidant nutrients) on fat oxidation, body composition and cardio-metabolic risk factors in overweight, but healthy individuals engaged in regular exercise. It was hypothesised that fat oxidation would be improved with dGTE (particularly when additional antioxidant nutrients are included), resulting in improved body composition and cardio-metabolic risk factors compared with placebo.
4. Discussion
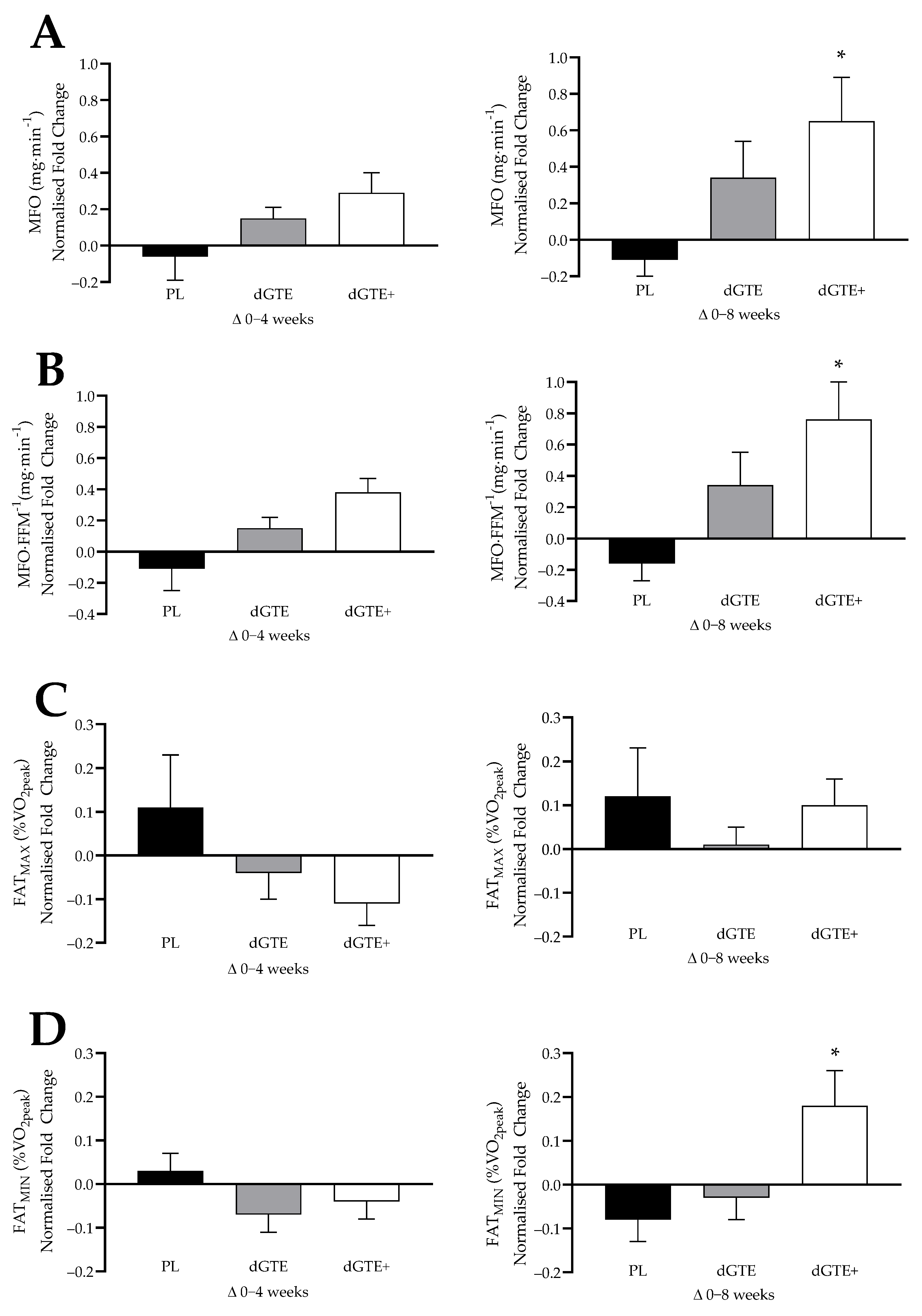
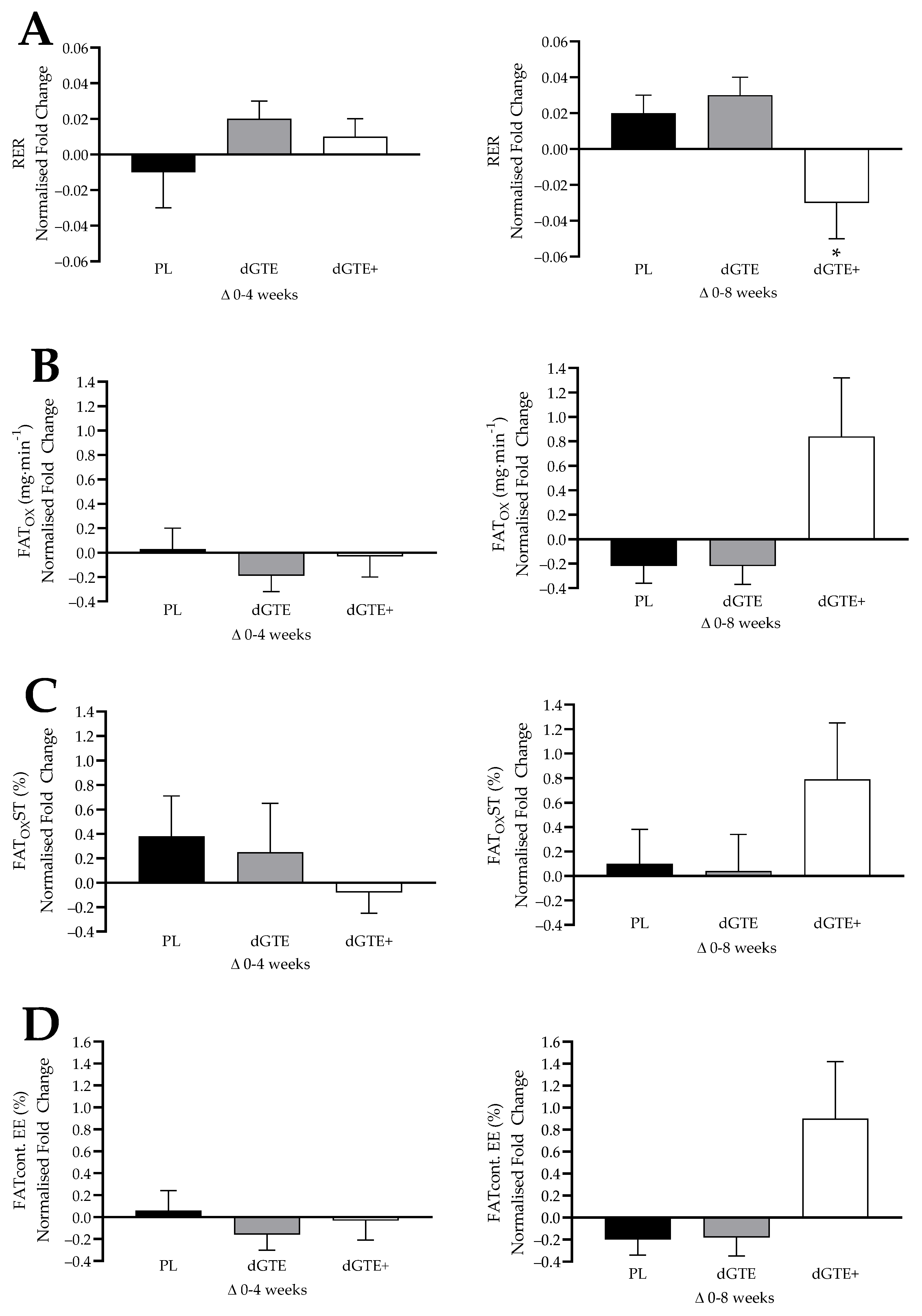
This study aimed to assess the longer-term impact of moderate dGTE supplementation (with or without antioxidant nutrients) on fat oxidation, body composition and cardio-metabolic risk factors in overweight, but healthy individuals engaged in regular exercise. The findings from the current study demonstrated that a moderate dose of dGTE consumed over 8 weeks did not result in a statistically significant change on fat oxidation rates, but did produce a non-significant increase in both MFO (+12.7%) and relative MFO (+13.9%). In contrast, the use of a novel dGTE+ formula, containing antioxidants in addition to dGTE, had a significant impact (moderate–large effect size) on MFO (+45.5%) and relative MFO (+49.4%) by week 8 of supplementation in recreationally active, overweight individuals. Additionally, a large effect size for FATMIN (+22.5%) was observed with dGTE+ over the last 4 weeks of this study. Metabolic changes were also observed by week 8 for dGTE+, with a large effect size reported for improvements in steady state substrate utilisation, evidenced by lower mean RER (−5.3%), improved relative FATOX (+55.2%) and increased the contribution of fat to total EE (+64.8%). Collectively, these findings suggest that a novel dGTE+ formula positively influenced fat oxidation in overweight individuals, particularly in the final 4 weeks of this study.
With regards to the methodological approaches to FAT
OX measures, concerns have been noted in the literature pertinent to reliability and variability [
53,
54,
55]. This was also observed in the current study with dGTE+ improving steady state FAT
OX by +28.4%, which was comparable to previous work undertaken [
25], despite being non-significant. Whilst mean MFO and steady state FAT
OX rates in the current study were comparable to previous research with untrained [
56], active [
57,
58,
59] or overweight individuals [
58,
60,
61,
62], a large inter-individual range was noted (MFO range: 26–353 mg·min
−1). As such, normalised data were also assessed to confirm or support findings. When data were normalised, taking into consideration individual responses to the intervention, MFO, relative MFO and FAT
MIN were all significantly enhanced with dGTE+ overall compared with PL, but not dGTE. However, for steady state exercise, normalised data only confirmed improved substrate utilisation for dGTE+ based on a reduced RER and the contribution of carbohydrate to total EE compared with dGTE. Whilst this further supports the contention that dGTE+ improved fat oxidation dynamics and substrate utilisation efficiency, data should be interpreted with caution based on individual ‘responders’ to the protocol. Overall FAT
MAX was not improved with dGTE+, despite a +24% increase in the latter 4 weeks of this study. This may be explained with the wide range of FAT
MAX values observed across this study (range: 24.1 to 52.4%
O
2peak).
Previous studies have reported mixed findings on the effect of GTE on fat oxidation, with several observing improvements in whole-body fat oxidation rates (+17–35% improvement employing a dose range of 270–400 mg·d
−1 EGCG) [
11,
17,
18,
19,
25] and no or small effects elsewhere (<+5–7% improvement with a dose range of 180–624 mg·d
−1 EGCG) [
21,
22,
23,
24,
26,
63]. Indeed, a recent meta-analysis [
64] highlighted a non-significant effect of EGCG supplementation on fat oxidation when compared with placebo (mean difference: 210 mg·min
−1), although these findings were based on pooled data from only two randomised controlled trials and should be interpreted with caution. Interestingly, beneficial effects of GTE have been mainly observed with healthy, active, untrained or sedentary individuals using acute (<2 days) moderate EGCG dose [
11,
17,
18,
19]. However, within these studies, it is unclear whether enhanced fat oxidation was due to accumulated GTE intake or dose consumed prior to exercise testing considering plasma EGCG concentrations peak at ~90 mins, with a reported half-life of ~10 hours [
65,
66]. It is further unclear whether these short-term studies have meaningful impact in the longer term. Chronic intake of dGTE, with comparable EGCG dose, but not consumed on the day of testing, has been shown to enhance fat oxidation and exercise performance in healthy, recreationally active males [
25]. However, the use of dGTE has been disputed elsewhere [
26] with physically active males, and appears to offer little benefit to trained individuals in terms of substrate utilisation and/or endurance exercise performance [
21,
22,
23,
24].
Collectively, these studies indicate that EGCG (both caffeinated and decaffeinated) may be effective in recreationally active or less trained individuals, potentially via thermogenic mechanisms pertinent to COMT inhibition and increased lipolytic activity leading to enhanced FAT
OX during exercise [
9,
14], although supporting evidence to confirm this is currently lacking. In the current study, it is noteworthy that normalised FFAs were greater for both dGTE groups compared with PL, albeit non-significantly. In the longer term, the effectiveness of this mechanism may be related to COMT genotype [
67] and/or specific adaptations in mitochondrial efficiency with regular EGCG consumption (particularly when coupled with exercise training). It has been proposed that EGCG consumption may act in a ‘calorie-restriction-mimetic’ manner through AMPK and sirtuin pathways. This may further modulate PGC1α, peroxisome proliferator-activated receptors (PPARs) and Forkhead box O (FOXO) gene expression, particularly in liver, adipose and skeletal muscle tissue [
14,
15,
68]. However, this has been disputed elsewhere in animal studies, with inference that adaptations in fatty acid translocase/CD36mRNA are more likely associated with improvements in fat oxidation [
69]. Future studies should aim to explore underlying mechanisms pertinent to fat metabolism gene expression, β-oxidation enzymes and fatty acid transporters following regular EGCG consumption. Further attention towards EGCG dose, as well as the effects of other potential calorie-restriction-mimetic nutrients (e.g., resveratrol, LA, curcumin, and other plant polyphenols [i.e., gallic acid]) on mitochondrial efficiency are also warranted.
In the current study, the specific use of dGTE did not significantly enhance fat oxidation measures in agreement with previous research [
21,
22,
26,
63]. However, the use of a novel dGTE+ formula does support findings particularly pertinent to less trained and/or overweight individuals [
11,
17,
19]. The inclusion of quercetin within the dGTE+ may have reduced degradation or oxidation of EGCG within the small intestine (especially considering dietary quercetin intakes were consistent between groups across the intervention). It has been suggested that this could enhance EGCG bioavailability either directly, or through increased EGCG delivery to the large intestine, microbiota conversion to EGCG ring-fission metabolites and phenolic compounds, and subsequent elevated plasma free and conjugated forms of EGCG [
31,
32,
65,
70]. Additionally, the increased bioavailability of quercetin, even at a low dose may have supported antioxidant mechanisms of EGCG pertinent to longer-term mitochondria efficiency [
70].
The additional inclusion of LA in the dGTE+ formula may have also enhanced EGCG stability through local antioxidant mechanisms, leading to increased EGCG availability. However, as EGCG bioavailability was not ascertained in the current study, this cannot be confirmed. As the use of dGTE only did not significantly influence FAT
OX, it is feasible the observed metabolic benefits associated with dGTE+ may be specific to LA. Whilst numerous mechanisms have been proposed, LA supports mitochondrial efficiency as a cofactor for α-ketoacid dehydrogenases [
71]. Reduced LA (DHLA) has also been implicated as a powerful natural antioxidant, highlighting that LA may support endogenous antioxidant status and mitochondrial protection [
71,
72,
73]. Additionally, LA has been proposed to influence AMPK pathways which may modulate muscle glucose uptake via GLUT4 translocation, and insulin sensitivity [
71,
72]. Therefore, in the longer term, it is feasible that, coupled with regular exercise training, LA could support enhanced mitochondrial efficiency, FAT
OX and subsequent weight loss. However, whilst a recent meta-analysis supports the contention that LA may elicit small, meaningful weight reduction benefits, studies ranging from 8–52 weeks have employed higher LA doses between 300 and 1800 mg·d
−1 [
74], which may explain such findings. As a lower dose of LA was employed in the current study, it is proposed that the synergistic inclusion of LA and quercetin may have acted to enhance EGCG bioavailability leading to up-regulated fat oxidation pathways in addition to regular exercise. Further research is warranted to confirm EGCG bioavailability with synergistic inclusion of quercetin and/or LA, along with cellular mechanistic pathways, particularly considering higher doses commonly used in commercial formulas.
Whilst the use of dGTE+ impacted on fat oxidation, by the end of the intervention, neither dGTE supplementation influenced overall body composition measures. This was despite relative, albeit small reductions in overall mean body mass (−0.50 kg), body fat percentage (−0.50%), fat mass (−0.37 kg), abdominal circumference (−0.77 cm) or SAD (−0.61 cm), particularly for dGTE+ compared with PL. Furthermore, the relative reduction in estimated VFA with dGTE+ was ~3-fold greater than for both dGTE and PL, albeit non-significant. Maintenance of habitual exercise only marginally impacted on body composition indices as evidenced by a significant reduction in waist and abdomen circumference for PL and dGTE at week 4 within group only. Collectively, the results indicate that when habitual exercise and dietary intake are controlled for, dGTE supplementation did not favour improved body composition indices. As dGTE+ only enhanced FAT
OX by week 8, any changes in body composition attributed to EGCG mechanisms may require longer interventions based on overall relative patterns in body composition indices with dGTE+. Indeed, a previous meta-analysis [
9] highlighted that GT consumption across studies exceeding 12 weeks resulted in a small, positive effect on body mass loss (−1.31 kg), with greater effects (−1.60 kg) observed for low habitual caffeine intake users.
However, the lack of efficacy of dGTE to modify body composition measures does concur with other longer-term studies (EGCG dose: 199–843 mg·d
−1) [
28,
67,
75], although physical activity was only partially monitored in these studies, and one study [
28] also introduced a low energy diet as part of the intervention. Elsewhere, significant improvements in body composition have been reported with GTE consumption (EGCG dose: 100–225 mg·d
−1) [
12,
13,
29], particularly when combined with an exercise intervention in previously sedentary individuals [
12,
29]. Interestingly, in one study [
29] exercise prescription began at 40% HR reserve (HRR) and progressed up to 50-59% HRR. Other studies utilising training at FAT
MAX (range:~34–52%
O
2max [
62,
76,
77]) in overweight, obese or diabetic women demonstrated significant improvements in body mass, BMI, fat mass and visceral trunk fat. In the current study, physical activity guidelines (>150 mins·wk
−1 at a moderate intensity ~60–65%
O
2peak in line with other studies [
18,
78]) were maintained. As this intensity was above participants’ FAT
MAX, this likely favoured carbohydrate metabolism and, in part, may explain the small decrease in MFO observed with PL. Combining dGTE+ with individualised FAT
MAX training in overweight individuals may therefore favourably improve body composition, warranting further research.
Cardio-metabolic variables were largely unaffected by either dGTE supplementation, with the exception of a medium effect size reported for LDL-c which was ~11% lower with dGTE+ by week 8, and further supported via normalised values. The relative reduction in total LDL-c across the intervention with dGTE+ (−0.38 ± 0.22 mmol·L
−1) was comparable to previous studies [
5,
79], which may be important as the mean values were considered elevated. This concurs with a recent meta-analysis indicating that LDL-c is likely reduced with regular green tea consumption (all sources) by −0.12 mmol·L
−1 [
80], and likely further reduced (−0.14 mmol·L
−1) when capsulated or decaffeinated green tea products are used [
80]. Interestingly, the observed effects of green tea consumption (all sources) appear greater over time, with noted reductions for total cholesterol (−0.14 mmol·L
−1) and triglycerides (−0.23 mmol·L
−1) in longer-term clinical trials (>12 weeks) [
80]. In the current study, as cholesterol biomarkers were largely unaffected with dGTE alone, this infers that the observed reductions in LDL-c with dGTE+ may have been attributed to either increased bioavailability via inclusion of low-dose quercetin, or separately via the inclusion of LA acting independently. Indeed, other reviews [
81] have reported significant reductions in LDL-c of −0.33 mmol·L
−1 with LA alone, which is comparable to that observed in the current study.
Both leptin and adiponectin levels were unaffected across the intervention with dGTE or dGTE+. This agrees with a recent meta-analysis [
82] demonstrating that leptin concentration is likely unaffected with GT (from all sources) when consumed for less than 12 weeks. In contrast, longer-term use of GT (>12 weeks) likely increases leptin concentrations (effect size: 2.90 [
82]). As appetite hormones and body composition changes were largely unaffected by either dGTE in the current study, this suggests that observed benefits of dGTE+ on fat oxidation by week 8 may have been attributed to gene expression associated with mitochondrial efficiency [
14]. It is noteworthy that adiponectin levels were significantly higher with dGTE+ compared to PL at baseline, week 4 and 8. Adiponectin has been associated with synergistic activation of AMPK, p38 mitogen-activated protein kinase (MAPK) and PPARα in skeletal muscle, and hence fatty acid oxidation [
83]. Higher adiponectin levels, combined with dGTE+, may have favourably altered AMPK kinetics, resulting in enhanced mitochondrial fatty acid oxidation across the intervention. In accordance with previous reviews [
82], it is feasible that longer-term dGTE consumption further influences appetite regulation favouring improved body composition characteristics, especially when combined with enhanced fat metabolism.
It is also noteworthy, when considering normalised values, that mean insulin, HOMA-IR and leptin changes were elevated with PL across the intervention (albeit non-significantly), with negligible changes observed for dGTE or dGTE+. It is therefore unclear whether the small changes observed for PL were related to dietary composition differences, or whether dGTE acted in a ‘protective’ manner. However, collectively, these results support previous findings that GTE was ineffective in improving cardio-metabolic function [
6,
27,
28], but differ from other studies where positive effects of GTE have been observed with [
30] or without exercise [
5,
13]. Interestingly, one other study [
84] employed a theaflavin-enriched GTE strategy, resulting in significant reductions in LDL-c and total cholesterol. This combination was proposed to reduce intestinal absorption and/or increased excretion of cholesterol, as well as LDL-c liver receptor up-regulation. The combined use of quercetin and LA with dGTE+ in the current study may have similarly enhanced dGTE effectiveness.
Whilst dGTE+ also resulted in a significant reduction in resting HR by week 4 compared to dGTE, but not by week 8, this did not impact on blood pressure measurements. This is somewhat surprising as a recent meta-analysis on green tea (from all sources) highlighted studies (≤12 weeks) resulting in a reduction of −1.32 mmHg for SBP (non-significant) and -1.78 mmHg for DBP (deemed significant), with lower observed effects for capsulated green tea, and green tea (all sources) consumed over longer periods (> 12 weeks) [
85]. In the current study, dGTE+ resulted in a reduction of −3.33 mmHg (for SBP) and -1.78 mmHg (for DBP), which exceeded the observed reduction expected for exercise/placebo (−2.44 mmHg for SBP and −0.78 mmHg for DBP), but was deemed non-significant. Whilst this may be partly explained by inter-individual variance in blood pressure responses, our results suggest that decaffeinated green tea extract did not impact on blood pressure response above that expected through maintenance of regular exercise.
An important aspect of the current study was the inclusion of liver enzyme and bilirubin measures. This was undertaken based on current concerns with the potential for GTE-induced hepatotoxicity [
86]. However, no significant elevations were observed for any measures, indicating relative safety of dGTE at a moderate dose across the intervention period. Whilst it has been noted that fasted intake may be a contributing factor to GTE-induced heptatoxicity [
87], other studies have highlighted the potential for reduced bioavailability when GTE is consumed with food [
20]. In the current study, participants were requested to consume dGTE away from food in a split-dose manner. As such, it is possible that consumption may have occurred in an absorptive state. Coupled with a lower single dose per consumption (i.e., 200 mg EGCG per capsule), this may, in part, explain normal liver function observed in the current study. Whilst this supports previous research [
6], and is in line with current recommendations [
87,
88], the efficacy of lower doses is warranted, as well as awareness of product composition. As example, unpublished data from our laboratory indicate that exposure to the specific dGTE used in this study impacts AMPK activity and mitochondrial oxidative stress to improve survival of hepatocytes in vitro. Therefore, due consideration of the effect of dGTE on oxidative stress pathways at a cellular level is also needed to understand the molecular mechanisms impacting MFO, as other products may differ in terms of biological effects.
It is important to note several limitations of the current study. As previously mentioned, EGCG bioavailability was not undertaken, and as no LA-specific intervention was included, findings should be interpreted with caution. Further studies investigating the specific effects of LA with or without dGTE are therefore warranted. In the current study a standardised dGTE (70% EGCG) product was sourced based on our previous research [
25]. Whilst this was independently verified, the content of other catechins (e.g., (−)-epicatechin-3-gallate (ECG)), flavonols (e.g., isoquercitin), flavonol glycosides and their aglycones, were not quantified. Whilst EGCG is the most abundant catechin within dGTE, the remaining catechin/flavonol compounds may have contributed to findings. Consideration to the presence, and biological activity, of other catechins/flavonols within dGTE supplementation is therefore warranted in future research.
In a similar context, consideration to dietary consistency and total polyphenol intake is also warranted. In the present study, relative dietary intake consistency was established in the 7 day period leading into each testing session. Furthermore, estimated quercetin intake (overall mean: 20.6 ± 8.5 mg·d
−1) was comparable between groups, and similar to previously reported research [
89,
90]. Therefore, any influence of the dGTE+ compound may have related to the enhanced bioavailability of EGCG with additional quercetin. However, it is feasible that individual dietary variance and total polyphenol intake outside of collection periods may have limited findings. Future research should therefore consider the impact of dGTE strategies in conjunction with low or high dietary polyphenol intake when combined with exercise strategies. Similarly, whilst exercise was monitored across the full intervention period, we did not investigate other components of physical activity such as non-exercise activity thermogenesis or sedentary periods which could affect overall metabolic flexibility and/or fat oxidation, as previously reported [
91,
92], even when physical activity guidelines are met. Finally, although testing sessions were standardised, as the majority of the participants were females, it is feasible that hormonal variance pertinent to menstrual cycle regulation may have influenced metabolic changes. However, this has been disputed elsewhere [
93].


 ,
, 





{kind=link}
{kind=link}