
A Mechanistic Study of the Antiaging Effect of Raw-Milk Cheese Extracts

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Milk and Cheese Samples
2.2. Reagents and Solvents
2.3. Obtaining of Milk Extracts
2.4. Obtaining of Cheese Extracts
2.5. Microbial Strains, Growth Conditions and Heat-Killed Preparation
2.6. C. elegans Maintenance
2.7. Synchronisation of Wild-Type C. elegans and Mutant Strains
2.8. Caenorhabditis elegans Longevity Assay Incubated with Dried Milk Extracts
2.9. Longevity Assay of DAF-16 Loss of Function Mutant (GR1307 Strain) Incubated with Dried Cheese Extracts
2.10. Cellular Localisation of DAF-16::GFP
2.11. Survival of the Worms on the Oxidative Medium
2.12. Determination of the Expression of Gene of Interest
2.12.1. Incubation of the Worms
2.12.2. RNA Isolation and RT-Quantitative PCR
2.13. Leukocyte Viability
2.14. Kinetics of ROS Production by Leukocytes
2.15. Statistical Analysis
3. Results
3.1. Implication of DAF-16 in the Capacity of the Extracts to Induce an Increase in Longevity
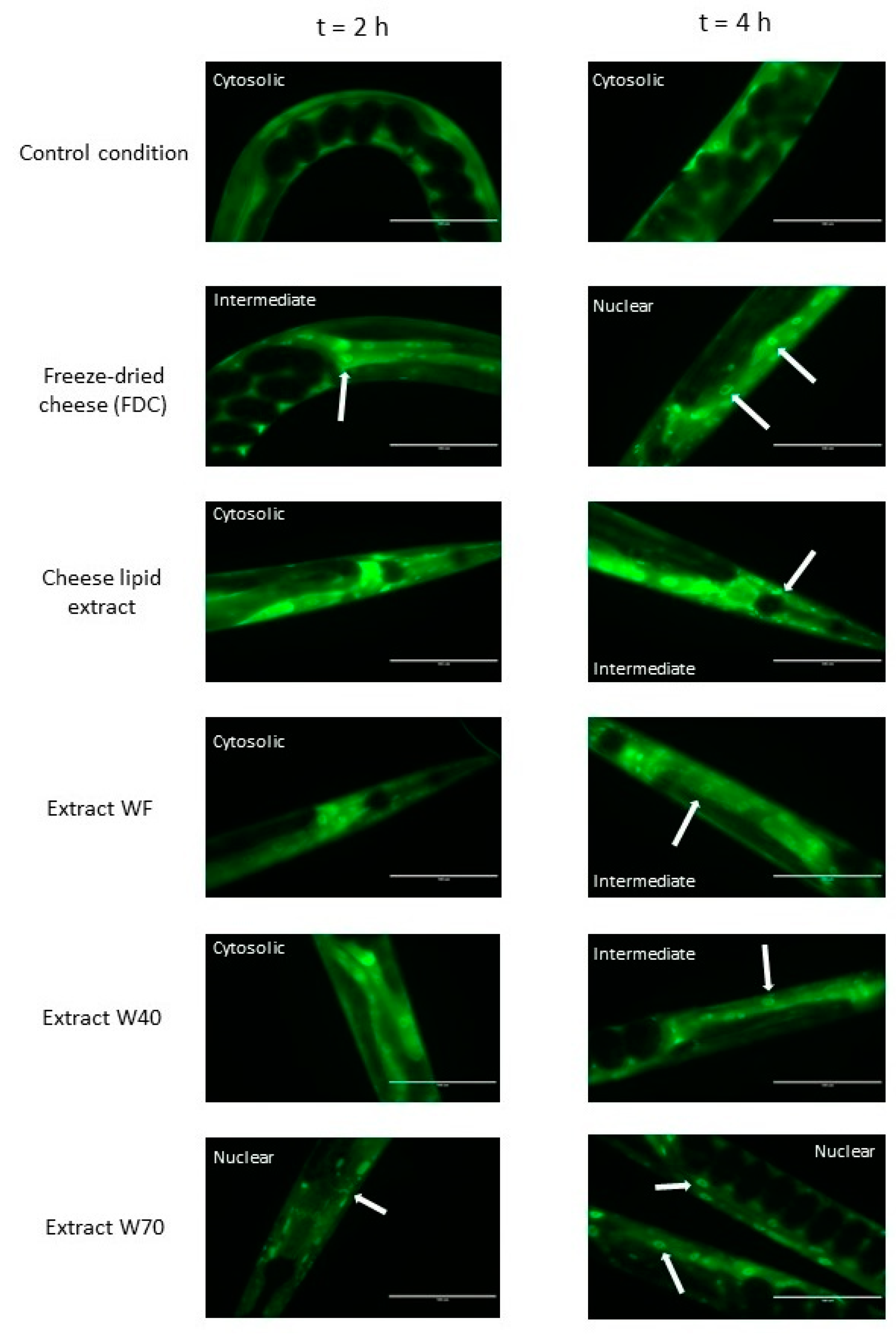
3.2. Cellular Localisation of DAF-16::GFP
3.3. Effect of the Dried Cheese Extracts on the Survival Rate of the Wild-Type C. elegans N2 Strain on the Oxidative Medium
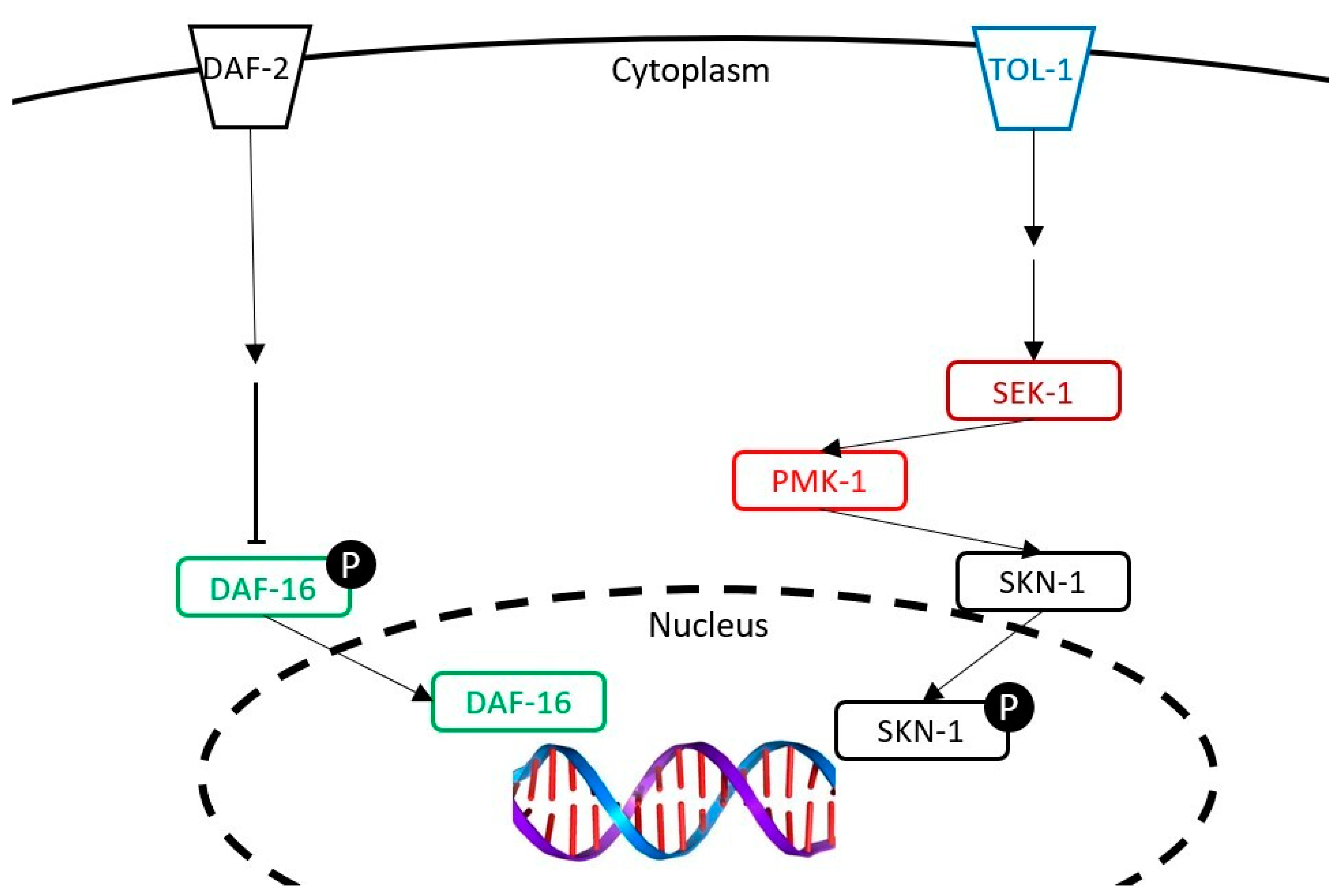
3.4. Implication of the Signaling Pathways in the Survival of the C. elegans on the Oxidative Medium
3.5. Evaluation of the Expression of the Genes of Interest (GOI) daf-16, sek-1 and pmk-1
3.6. Production of ROS in Human Blood Leukocytes Triggered by PMA
3.7. Effect of the Dried Milk Extracts on the Longevity of Wild-Type C. elegans N2 Strain and Its Survival Rate on the Oxidative Medium
4. Discussion
4.1. The Transcription Factor DAF-16 Is Implicated in the Mechanism by Which the Dried Cheese Extracts Increase Longevity
4.2. The Dried Cheese Extracts Influenced the Survival of the Worms on the Oxidative Medium
4.3. Influence of the Dried Cheese Extracts on ROS Production in Human Leukocytes
4.4. The Milk and Its Extracts Exert a Lower Beneficial Effect on the Wild-Type C. elegans N2 Strain Compared with the Cheese and Its Extracts
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Phelan, M.; Kerins, D. The potential role of milk-derived peptides in cardiovascular disease. Food Funct. 2011, 2, 153–167. [Google Scholar] [CrossRef]
- Muro Urista, C.; Álvarez Fernández, R.; Riera Rodriguez, F.; Arana Cuenca, A.; Téllez Jurado, A. Review: Production and functionality of active peptides from milk. Food Sci. Technol. Int. 2011, 17, 293–317. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, S.; Kuwata, H.; Kawamoto, K.; Shirakawa, J.; Atobe, S.; Hoshi, Y.; Yamasaki, M.; Nishiyama, K.; Tachibana, H.; Yamada, K.; et al. Effect of highly lipolyzed goat cheese on HL-60 human leukemia cells: Antiproliferative activity and induction of apoptotic DNA damage. J. Dairy Sci. 2012, 95, 2248–2260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardin, G.; Ripoche, I.; Poupet, C.; Bonnet, M.; Veisseire, P.; Chalard, P.; Chauder, A.; Saunier, E.; Priam, J.; Bornes, S.; et al. Development of an innovative methodology combining chemical fractionation and in vivo analysis to investigate the biological properties of cheese. PLoS ONE 2020, 15, e0242370. [Google Scholar] [CrossRef]
- Wilson, M.A.; Shukitt-Hale, B.; Kalt, W.; Ingram, D.K.; Joseph, J.A.; Wolkow, C.A. Blueberry polyphenols increase lifespan and thermotolerance in Caenorhabditis elegans. Aging Cell 2006, 5, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Clark, L.C.; Hodgkin, J. Commensals, probiotics and pathogens in the Caenorhabditis elegans model. Cell. Microbiol. 2014, 16, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Park, M.R.; Yun, H.S.; Son, S.J.; Oh, S.; Kim, Y. Short communication: Development of a direct in vivo screening model to identify potential probiotic bacteria using Caenorhabditis elegans. J. Dairy Sci. 2014, 97, 6828–6834. [Google Scholar] [CrossRef] [Green Version]
- Poupet, C.; Chassard, C.; Nivoliez, A.; Bornes, S. Caenorhabditis elegans, a host to reveal and investigate the probiotic properties of beneficial microorganisms. Front. Nutr. 2020, 7, 135. [Google Scholar] [CrossRef]
- Roselli, M.; Schifano, E.; Guantario, B.; Zinno, P.; Uccelletti, D.; Devirgiliis, C. Caenorhabditis elegans and Probiotics Interactions from a Prolongevity Perspective. Int. J. Mol. Sci. 2019, 20, 5020. [Google Scholar] [CrossRef] [Green Version]
- Murphy, C.T.; McCarroll, S.A.; Bargmann, C.I.; Fraser, A.; Kamath, R.S.; Ahringer, J.; Li, H.; Kenyon, C. Genes that act downstream of DAF-16 to influence the lifespan of Caenorhabditis elegans. Nature 2003, 424, 277–284. [Google Scholar] [CrossRef]
- Lee, S.S.; Kennedy, S.; Tolonen, A.C.; Ruvkun, G. DAF-16 target genes that control C. elegans Life-span and metabolism. Science 2003, 300, 644–647. [Google Scholar] [CrossRef] [Green Version]
- Kampkötter, A.; Pielarski, T.; Rohrig, R.; Timpel, C.; Chovolou, Y.; Wätjen, W.; Kahl, R. The Ginkgo biloba extract EGb761 reduces stress sensitivity, ROS accumulation and expression of catalase and glutathione S-transferase 4 in Caenorhabditis elegans. Pharmacol. Res. 2007, 55, 139–147. [Google Scholar] [CrossRef]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Poupet, C.; Saraoui, T.; Veisseire, P.; Bonnet, M.; Dausset, C.; Gachinat, M.; Camarès, O.; Chassard, C.; Nivoliez, A.; Bornes, S. Lactobacillus rhamnosus Lcr35 as an effective treatment for preventing Candida albicans infection in the invertebrate model Caenorhabditis elegans: First mechanistic insights. PLoS ONE 2019, 14, e0216184. [Google Scholar] [CrossRef] [PubMed]
- Porta-de-la-Riva, M.; Fontrodona, L.; Villanueva, A.; Cerón, J. Basic Caenorhabditis elegans methods: Synchronization and observation. J. Vis. Exp. 2012, 64, e4019. [Google Scholar] [CrossRef] [Green Version]
- Breger, J.; Fuchs, B.B.; Aperis, G.; Moy, T.I.; Ausubel, F.M.; Mylonakis, E. Antifungal Chemical Compounds Identified Using a C. elegans Pathogenicity Assay. PLoS Pathog. 2007, 3, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Fatima, S.; Haque, R.; Jadiya, P.; Kumar, L.; Nazir, A. Ida-1, the Caenorhabditis elegans Orthologue of Mammalian Diabetes Autoantigen IA-2, Potentially Acts as a Common Modulator between Parkinson’ s Disease and Diabetes: Role of Daf-2/Daf-16 Insulin Like Signalling Pathway. PLoS ONE 2014, 9, e113986. [Google Scholar] [CrossRef] [Green Version]
- Grompone, G.; Martorell, P.; Llopis, S.; González, N.; Genovés, S.; Mulet, A.P.; Fernández-Calero, T.; Tiscornia, I.; Bollati-Fogolín, M.; Chambaud, I.; et al. Anti-Inflammatory Lactobacillus rhamnosus CNCM I-3690 Strain Protects against Oxidative Stress and Increases Lifespan in Caenorhabditis elegans. PLoS ONE 2012, 7, e52493. [Google Scholar] [CrossRef] [PubMed]
- Poupet, C.; Veisseire, P.; Bonnet, M.; Camarès, O.; Gachinat, M.; Dausset, C.; Chassard, C.; Nivoliez, A.; Bornes, S. Curative treatment of candidiasis by the live biotherapeutic microorganism Lactobacillus rhamnosus lcr35® in the invertebrate model Caenorhabditis elegans: First mechanistic insights. Microorganisms 2020, 8, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol. 2007, 8, R19. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, H.; Shiozaki, T.; Kobatake, E.; Hosoya, T.; Moriya, T.; Sakai, F.; Taru, H.; Miyazaki, T. Effects and mechanisms of prolongevity induced by Lactobacillus gasseri SBT2055 in Caenorhabditis elegans. Aging Cell 2016, 15, 227–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millet, A.C.M.; Ewbank, J.J. Immunity in Caenorhabditis elegans. Curr. Opin. Immunol. 2004, 16, 4–9. [Google Scholar] [CrossRef]
- The C. elegans Sequencing Consortium Genome Sequence of the Nematode C. elegans: A Platform for Investigating Biology. Science 1998, 282, 2012–2018.
- Finch, C.E.; Ruvkun, G. The genetics of aging. Annu. Rev. Genomics Hum. Genet. 2001, 2, 435–462. [Google Scholar] [CrossRef] [Green Version]
- Kondo, M.; Yanase, S.; Ishii, T.; Hartman, P.S.; Matsumoto, K.; Ishii, N. The p38 signal transduction pathway participates in the oxidative stress-mediated translocation of DAF-16 to Caenorhabditis elegans nuclei. Mech. Ageing Dev. 2005, 126, 642–647. [Google Scholar] [CrossRef]
- Nakahara, T.; Moroi, Y.; Uchi, H.; Furue, M. Differential role of MAPK signaling in human dendritic cell maturation and Th1/Th2 engagement. J. Dermatol. Sci. 2006, 42, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pastuhov, S.I.; Hisamoto, N.; Matsumoto, K. MAP kinase cascades regulating axon regeneration in C. elegans. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2015, 91, 63–75. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extracts | Concentration (w/v) | ||
|---|---|---|---|
| 0.25% | 0.5% | 1% | |
| Freeze-dried milk (FDM) | X | X | |
| Milk lipid extract (ML) | X | X | X |
| Lipid-free milk (LFM) | X | X | X |
| Extract MA | X | X | |
| Extract MB | X | ||
| Extract MC | X | X | |
| Residual solid milk (RSM) | X | X | X |
| Gene Name | Gene Type | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Reference |
|---|---|---|---|---|
| Y45F10D.4 | housekeeping | CGAGAACCCGCGAAATGTCGGA | CGGTTGCCAGGGAAGATGAGGC | [19] |
| daf-16 | GOI | TTCAATGCAAGGAGCATTTG | AGCTGGAGAAACACGAGACG | [19,21] |
| sek-1 | GOI | GCCGATGGAAAGTGGTTTTA | TAAACGGCATCGCCAATAAT | [19,21] |
| pmk-1 | GOI | CCGACTCCACGAGAAGGATA | AGCGAGTACATTCAGCAGCA | [19,21] |
| Tested Conditions | Concentration (w/v) (%) | Mean Lifespan (Days) | Maximum Lifespan (Days) | Relative Increase in the Maximum Lifespan (%) | Percentage of Population Still Alive at 17 Days (%) | p-Value |
|---|---|---|---|---|---|---|
| CC2 | - | 10 | 17 | - | 0 | - |
| Freeze-Dried Cheese (FDC) | 0.5 | 10 | 17 | 0 | 0 | 0.001 |
| 1 | 10 | 17 | 0 | 0 | 0.03 | |
| Cheese-lipid extract | 0.5 | 10 | 18 | +6 | 1 | 0.02 |
| 1 | 10 | 16 | −6 | 0 | 0.5 |
| Tested Conditions | Concentration (w/v) (%) | Mean Lifespan (Days) | Maximum Lifespan (Days) | Relative Increase in the Maximum Lifespan (%) | Percentage of Population Still Alive at 17 Days (%) | p-Value |
|---|---|---|---|---|---|---|
| CC1 | - | 9 | 17 | - | 0 | - |
| Extract WF (40 °C) | 0.5 | 7 | 19 | +12 | 1 | <0.0001 |
| 1 | 8 | 19 | +12 | 2 | 0.0008 | |
| Extract W40 (40 °C) | 0.5 | 8 | 19 | +12 | 2 | <0.0001 |
| 1 | 8 | 20 | +18 | 1 | 0.0002 | |
| Extract W70 (70 °C) | 0.5 | 10 | 22 | +29 | 2 | 0.9 |
| 1 | 9 | 22 | +29 | 5 | 0.09 |
| Genes of Interest | |||
|---|---|---|---|
| Conditions | daf-16 | sek-1 | pmk-1 |
| FDC 3 days | 1.95 | 1.13 | 1.00 |
| FDC 10 days | 2.78 ** | 2.89 * | 2.80 * |
| Cheese-lipid extract 3 days | 1.07 | 1.02 | 1.09 |
| Cheese-lipid extract 10 days | 3.41 ** | 1.19 | 2.39 ** |
| Genes of Interest | |||
|---|---|---|---|
| Conditions | daf-16 | sek-1 | pmk-1 |
| WF (40 °C) 3 days | 5.52 ** | 1.11 | 1.27 |
| WF (40 °C) 10 days | 1.00 | 1.87 | 1.01 |
| W40 (40 °C) 3 days | 1.08 | 0.83 | 1.24 |
| W40 (40 °C) 10 days | 2.93 ** | 3.39 | 0.67 |
| W70 (70 °C) 3 days | 1.20 | 0.80 | 1.08 |
| W70 (70 °C) 10 days | 1.96 | 1.26 | 0.59 |
| Tested Conditions | Concentration (w/v) (%) | Mean Lifespan (Days) | Maximum Lifespan (Days) | Relative Increase of the Maximum Lifespan (%) | Percentage of Population Still Alive at 23 Days (%) | p-Value |
|---|---|---|---|---|---|---|
| CC1 | - | 12 | 23 | - | 0 | - |
| Freeze-Dried Milk (FDM) | 0.25 | 16 | 31 | +35 | 6 | <0.0001 |
| 0.5 | 15 | 26 | +13 | 5 | <0.0001 | |
| Milk Lipid Extract | 0.25 | 14.5 | 26 | +13 | 3 | <0.0001 |
| 0.5 | 15 | 30 | +30 | 5 | <0.0001 | |
| 1 | 15 | 23 | 0 | 0 | <0.0001 | |
| Extract MA | 0.25 | 15 | 31 | +35 | 5 | <0.0001 |
| 0.5 | 14 | 28 | +22 | 3 | <0.0001 | |
| Extract MB | 0.5 | 7 | 22 | −4 | 0 | <0.0001 |
| Extract MC | 0.25 | 14 | 26 | +13 | 2 | <0.0001 |
| 0.5 | 15 | 28 | +22 | 4 | <0.0001 | |
| Residual Solid Milk (RSM) | 0.25 | 15 | 28 | +22 | 5 | <0.0001 |
| 0.5 | 15 | 25 | +9 | 5 | <0.0001 | |
| 1 | 15 | 35 | +52 | 4 | <0.0001 |
| Tested Conditions | Concentration (w/v) (%) | Mean Lifespan (Days) | Maximum Lifespan (Days) | Relative Increase of the Maximum Lifespan (%) | Percentage of Population Still Alive at 23 Days (%) | p-Value |
|---|---|---|---|---|---|---|
| CC2 | - | 12 | 23 | - | 0 | - |
| Lipid-Free Milk (LFM) | 0.25 | 13 | 28 | +22 | 5 | <0.0001 |
| 0.5 | 14 | 29 | +26 | 7 | <0.0001 | |
| 1 | 15 | 24 | +4 | 5 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardin, G.; Poupet, C.; Bonnet, M.; Veisseire, P.; Ripoche, I.; Chalard, P.; Chauder, A.; Saunier, E.; Priam, J.; Bornes, S.; et al. A Mechanistic Study of the Antiaging Effect of Raw-Milk Cheese Extracts. Nutrients 2021, 13, 897. https://doi.org/10.3390/nu13030897
Cardin G, Poupet C, Bonnet M, Veisseire P, Ripoche I, Chalard P, Chauder A, Saunier E, Priam J, Bornes S, et al. A Mechanistic Study of the Antiaging Effect of Raw-Milk Cheese Extracts. Nutrients. 2021; 13(3):897. https://doi.org/10.3390/nu13030897
Chicago/Turabian StyleCardin, Guillaume, Cyril Poupet, Muriel Bonnet, Philippe Veisseire, Isabelle Ripoche, Pierre Chalard, Anne Chauder, Etienne Saunier, Julien Priam, Stéphanie Bornes, and et al. 2021. "A Mechanistic Study of the Antiaging Effect of Raw-Milk Cheese Extracts" Nutrients 13, no. 3: 897. https://doi.org/10.3390/nu13030897
APA StyleCardin, G., Poupet, C., Bonnet, M., Veisseire, P., Ripoche, I., Chalard, P., Chauder, A., Saunier, E., Priam, J., Bornes, S., & Rios, L. (2021). A Mechanistic Study of the Antiaging Effect of Raw-Milk Cheese Extracts. Nutrients, 13(3), 897. https://doi.org/10.3390/nu13030897